
野生花生全基因组抗病相关LOX基因的生物信息学分析
2015-12-23 09:32宋辉赵术珍侯蕾乔善良赵传志李爱芹王兴军
山东农业科学 2015年10期
关键词:抗病
宋辉 赵术珍 侯蕾 乔善良 赵传志 李爱芹 王兴军


摘要:脂肪氧合酶(lipoxygenases,LOX)是生物体内一种重要的双加氧酶,其在植物发育以及响应生物和非生物胁迫时发挥重要作用。本研究从Arachis duranensis和Arachis ipaensis两个野生花生基因组中各鉴定出18个LOX基因,分别命名为AdLOX1~18和AiLOX1~18。在AdLOX5、AdLOX7、AiLOX8和AiLOX18基因的CDS序列中发现提前终止翻译的终止密码子,而AiLOX9基因的CDS序列过短,推测这些基因可能是假基因。染色体定位结果显示,AdLOX和AiLOX基因主要分布在2、3、6、8、9和10号染色体上,且其直系同源基因分布在两套染色体相同或相近的位置。通过分析AdLOX和AiLOX基因与已知功能LOX基因的亲缘关系发现,AdLOX6、、AdLOX9、AdLOX11、AdLOX12、AdLOX17、AiLOX1、AiLOX4、AiLOX6、AiLOX16和AiLOX17基因可能参与花生的抗病过程,启动子分析结果也支持这一结论。基因选择压力研究表明,AdLOX和AiLOX基因在进化历程中受到纯化选择。
关键词:野生花生;LOX基因家族;抗病;系统发育;选择压力
中图分类号:S656.201
文献标识号:A
文章编号:1001-4942(2015)10-0001-07
花生(Arachis hypogaea L.)作为重要的油料经济作物被广泛地种植在热带和亚热带地区。目前,花生属中已经鉴定了80个种。大多数的野生花生为二倍体,而栽培花生为异源四倍体(AABB)植物。大量研究表明,栽培花生的二倍体祖先分别是提供A基因组的Arachis duranensis和提供B基因组的Arachis ipaensis。栽培花生对多种病虫害的抗性低,近年来,病虫害危害严重,造成花生大面积减产。而野生花生由于长期适应不同的恶劣环境,对病虫害有很高的抗性,甚至对有些严重影响栽培花生的病害免疫。因此,研究野生花生对病原菌的抗性机理,鉴定、挖掘野生花生中的抗病基因资源,对培育抗病能力强的优异花生新品种具有重要意义。2014年,A.duranensis和A.ipaensis全基因组测序工作已经完成,为从全基因组水平上分析和鉴定野生花生的抗病基因、研究其抗病机理奠定了坚实基础。
LOX蛋白是一种非血红素铁酶(non-hemeiron containing enzymes),该酶广泛地分布在动物、植物和微生物中。其主要功能是通过LOX途径降低多不饱和脂肪酸(polyunsaturated fattyacids)的含量。在LOX途径中,过氧化氢物(hydroperoxides)被转化为氧化脂类(oxylipins),而氧化脂类能激活LOX途径下游的多种酶,包括过氧化氢裂解酶(hydroperoxide lyase)、过氧化氢环化酶(peroxygenase)、丙二烯氧合酶(allene oxide synthase)和二乙烯基醚合酶(divinyl ether synthase)。在植物中,氧化脂类和其衍生物,例如茉莉酸和乙醛等,已经被证实参与植物的抗病和抗虫等过程。因此,研究LOX基因对阐明植物的抗病和抗虫机理具有重要的作用。在拟南芥(Arabidopsis thaliana)中,AtLOX2基因被确定参与茉莉酸途径;当拟南芥受到生物胁迫时,AtLOX1基因的表达也会上调。栽培花生LOX基因在抗病方面的研究已有报道。Burow等克隆了栽培花生AhLOX1基因,当花生受到茉莉酸、伤害和曲霉侵染时,该基因的表达会显著提高。在栽培花生成熟种子中过量表达AhLOX2和AhLOX3基因,会提高其抵御黄曲霉侵染的能力。但花生LOX基因家族的系统研究尚未见报道。
目前,关于LOX基因家族的分类主要有两种方法。第一种方法是根据亚油酸(linoleic acid)被氧化的顺序分为9-LOX(C-atom 9)和13-LOX(C-atom 13)。第二种方法是根据氨基酸序列的相似性将LOX基因分为Type I-LOX和TypeⅡ-LOX,Type I-LOX的序列相似性约为75%,其N段缺少一段叶绿体运输多肽;而TypeⅡ-LOX的序列相似性约为35%,其N段存在叶绿体运输多肽,且具有13-LOX活性。
本研究利用生物信息学方法从A.duranensis和A.ipaensis基因组中分别鉴定出18个LOX基因,从分子系统进化、染色体定位和选择压力方面对AdLOX和AiLOX基因进行了系统的分析,并参考已报道的参与植物免疫过程的LOX基因,预测了在A.duranensis和A.ipaensis中参与植物免疫反应的LOX基因。
1材料与方法
1.1二倍体野生花生LOX基因序列的获得
从花生基因组计划网站(http://peanutbase.org/)下载A.duranensis和A.ipaensis基因组序列。以A.duranensis和A.ipaensis的氨基酸序列作为数据库,使用Lipoxygenase结构域(PF00305.15)作为查询序列,采用HMMER程序分别搜寻以上两个物种的LOX氨基酸序列。将得到的所有序列重新提交到Pfam数据库,剔除只含有Lipoxygenase结构域的序列,保留同时含有Lipoxygenase和PLAT/LH两种结构域的序列。
1.2LOX基因的系统进化和染色体定位分析
为对AdLOX和AiLOX基因进行分类和明确其进化关系,本研究在NCBI数据库中下载了单子叶植物大麦(Hordeum vulgare)、水稻(Oryza sativa)、玉米(Zea mays)和小麦(Triticum aestivum)以及双子叶植物拟南芥、花生、大豆(Glycine max)、豌豆(Pisum sativum)和菜豆(Phaseolus vulgaris)的LOX基因信息,使用MEGA 6.0软件对下载的LOX氨基酸序列及AdLOX和AiLOX氨基酸序列构建系统发育树。系统发育树采用邻接法(NJ),自举值(bootstrap)设置为1000。另外,为挖掘两种二倍体野生花生LOX基因的直系同源基因,使用AdLOX和AiLOX氨基酸序列构建系统发育树,如果AdLOX和AiLOX聚集在系统发育树末端的同一个分支上,则认为该基因对为直系同源基因。endprint
AdLOX和AiLOX基因在染色体上的定位信息从公布基因组序列的在线数据库http://pea-nutbase.org/获得。使用MapInspect软件(http://mapinspect.software.informer.com/)构建基因的染色体定位图。
1.3LOX基因启动子顺式作用元件分析
顺式作用元件(cis-acting element)位于转录起始位点的上游区域,具有调控基因表达的功能。为预测抗病相关LOX基因的顺式元件,本研究使用在线数据库PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)作为分析工具、以转录位点上游2000bp序列作为分析序列进行分析。
1.4选择压力分析
基因在长期的进化过程中经受不同的选择压力。评价基因在进化中所受选择压力的参数主要有两个:同一性替换(synonymous substitution,Ks)和非同一性替换(Non-synonymous substitution,Ka)。如果Ka/Ks>1,则表示受正选择压力;Ka/Ks=1,表示受中性选择;Ka/Ks<1,表示受纯化选择。本研究使用PAML程序的位点模型(site model)分别计算AdLOX和AiLOX蛋白在进化中所受的选择压力。设置M0(one ratio)、M1a(neutral)、M2a(selection)、M3(discrete)、M7(β)和M8[(β+ω)>1]6种计算模型,根据LRT的值判断在M1a vs M2a、M0 vs M3和M7 vs M8下的选择压力。
2结果与分析
2.1LOX基因的鉴定与分类
本研究使用HMMER程序共检测到23条AdLOX和25条AiLOX氨基酸序列含有Lipoxygenase结构域。先前的研究已经证实,只有同时含有Lipoxygenase和PLAT/LH结构域的序列才为LOX基因家族。使用Pfam数据库分析23条AdLOX和25条AiLOX氨基酸序列发现,18条AdLOX和18条AiLOX氨基酸序列同时含有以上两个结构域。根据测序的ID号,分别将AdLOX和AiLOX序列命名为AdLOX1~18和AiLOX1~18。进一步分析CDS和氨基酸序列证实,AdLOX5、AdLOX7、AiLOX8和AiLOX18的CDS序列中均含有提前终止翻译的终止密码子;AiLOX9的CDS序列长度只有525bp。推测以上序列可能是假基因或由于测序质量导致,在本研究中不对以上序列进行分析。余下16条AdLOX的CDS序列长度在1530~4374bp之间,氨基酸长度为509~1457aa;15条AiLOX的CDS序列长度为1737~2793bp,氨基酸序列长度为578~930aa(表1)。
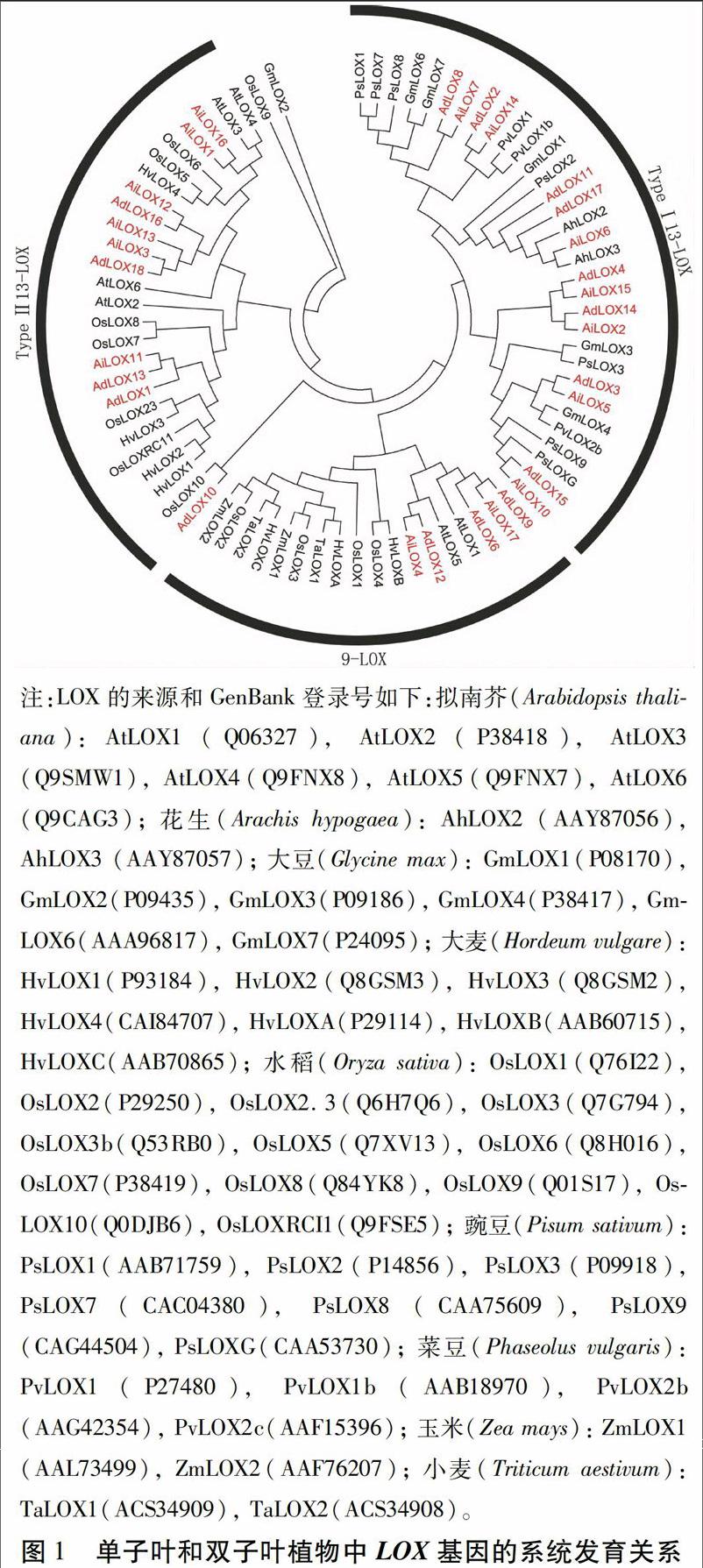
LOX基因家族可以分为2个亚族,即9-LOX和13-LOX,根据其氨基酸结构可以进一步将13-LOX分为两类,即Type I 13-LOX和TypeⅡ13-LOX。使用单子叶植物大麦、水稻、小麦和玉米以及双子叶植物拟南芥、大豆、菜豆和豌豆的LOX氨基酸序列与野生花生AdLOX和AiLOX氨基酸序列构建系统发育树,结果表明,AdLOX分别含有3个9-LOX、8个Type I 13-LOX和5个TypeⅡ13-LOX类型基因。AiLOX分别含有2个9-LOX、7个Type I 13-LOX和6个TypeⅡ13-LOX基因(图1)。此外,Type I 13-LOX的分支中仅含有双子叶豆科植物LOX基因,而9-LOX和TypeⅡ13-LOX分支中同时含有双子叶植物和单子叶植物LOX基因,表明Type I13-LOX类型基因是在豆科植物分化以后形成的,而9-LOX和TypeⅡ13-LOX类型基因是在单子叶植物和双子叶植物分化之前已经出现。
由系统发育树可知,AdLOX和AiLOX基因与AtLOX1、AtLOX3、AtLOX4、AtLOX5、GmLOX4、OsLOX10、PsLOXG、PvLOX1、PvLOX1b和PvLOX2b的亲缘关系较近(图1)。其中,AtLOX1与AdLOX6、AdLOX9和AiLOX17聚集在一个分支;AtLOX3和AtLOX4与AiLOX1和AiLOX16聚集在一个分支;AtLOX5与AdLOX12和AiLOX4聚集在一个分支。研究发现,AtLOX1和AtLOX5的表达能提高植物体对病原菌和蚜虫的抗性,而AtLOX3和AtLOX4的过量表达能提高植物对线虫的抗性。由此推断,AdLOX6、AdLOX9、AdLOX12、AiLOX1、AiLOX4、AiLOX16和AiLOX17基因可能参与野生花生的免疫反应。在栽培花生中已确定AhLOX2和AhLOX3基因参与花生的抗病过程。在系统发育树中,AhLOX2和AhLOX3基因和AdLOX11、AdLOX17以及AiLOX6聚集在同一个分支上,由此推测,AdLOX11、AdLOX17和AiLOX6也可能在野生花生抗病过程中起着重要作用。先前的研究证实,PvLOX1基因在植物的发育过程中起一定的作用,AdLOX2和AiLOX14基因与其聚集在同一个分支,由此判断这两个基因也可能具有相似的功能;而GmLOX4、OsLOX10、PsLOXG、PvLOX1b和PvLOX2b基因的功能尚不完全明确,与这些基因聚集在一起的AdLOX和AiLOX基因的功能需要进一步的验证。
2.2LOX基因染色体定位
二倍体野生花生共有10条染色体。根据AdLOX和AiLOX基因在染色体上的定位信息可知AdLOX基因分布在6条染色体上,分别是2、3、6、8、9和10号染色体;AiLOX基因分布在7条染色体上,分别是2、3、4、6、8、9和10号染色体(图2)。AdLOX基因在9号染色体上分布的最多,有8个成员;其次,3和8号染色体上分别有3个AdLOX基因;6号染色体上含有2个AdLOX基因;而2和10号染色体上分别有1个AdLOX基因。于此相似,AiLOX基因在9号染色体上分布最多,共有7个;3和6号染色体上各3个;8号染色体上2个;2、4和10号染色体上各分布1个AiLOX基因。进一步分析表明,AdLOX和AiLOX基因在9号染色体上成簇集中分布,其中在A基因组的9号染色体和B基因组的9号染色体6.3Mb和10.2Mb的范围内各有6个LOX基因(表1和图2),表明9号染色体是LOX基因的起源位点。该结果对野生花生与栽培花生近缘杂交、野生种质抗病基因资源的利用以及分子标记辅助选择具有重要指导意义。endprint
通过使用系统发育树法共鉴定出12对AdLOX和AiLOX基因的直系同源基因对(图3)。A.duranensis和A.ipaensis直系同源基因对分别分布在2、3、6、8和9号染色体上,其中9号染色体上分布的最多,有6对;3和8号染色体上各有2对,2和6号染色体上各有1对。对直系同源基因对的分析发现,这些基因对对应于A.duranensis和A.ipaensis染色体上相同或相似的位置,且其结构域保守,表明这些基因在功能上也相对保守;相反,有些在两套染色体相同或相近位置上的基因却不存在直系同源关系,如AdLOX5和AiLOX13(图2),可能这些基因对在进化过程中发生了变异,或已分化出不同的生物学功能。
2.3启动子抗病相关顺式作用元件分析
通过分析AdLOX和AiLOX顺式作用元件发现,所有预测到参与抗病过程的AdLOX和AiLOX基因的启动子区域均含有参与抗病响应的顺式作用元件,包括Box-W1、CGTCA-motif、TC-richrepeats、TCA-element和TGACG-motif,主要参与真菌诱导响应、茉莉酸途径、免疫防御响应和水杨酸途径(表2)。在10个抗病相关的LOX基因中,除了AdLOX9,其他9个基因均含有响应水杨酸或茉莉酸的顺式作用元件。茉莉酸途径主要响应死体型病原菌(necrotrophic pathogen)的侵入,而水杨酸途径主要响应活体营养性病原菌(biotrophic pathogen)的侵入。虽然茉莉酸和水杨酸途径响应不同类型病原菌的侵入,但也有研究表明,这两种植物激素在防御病原菌侵入的过程中具有协同效应(synergistic effect)。参与抗病反应的AdLOX和AiLOX基因具有响应水杨酸和(或)茉莉酸途径的顺式作用元件,表明AdLOX和AiLOX基因能抵御多种病原菌的侵入。
2.4选择压力分析
为了检测哪种选择压力影响AdLOX和AiLOX基因的结构(或功能),本研究使用PAML程序中的位点模型分别计算了AdLOX和AiLOX蛋白所受的选择压力。由表3可知,大多数Ka/Ks<1,且未检测到正选择位点,表明AdLOX和AiLOX基因在进化过程中受到纯化选择。纯化选择会使基因的结构和生物学功能更加保守。Chen等对毛果杨(Populus trichocarpa)LOX基因在进化过程中所受到的选择压力研究发现,大多数旁系同源基因对Ka/Ks<0.4,表明这些基因在进化历程中受到纯化选择。这些结果表明,LOX基因可能在蛋白质水平上的进化相对缓慢。
3结论
本研究利用生物信息学方法对两个野生花生种的LOX基因家族进行了鉴定和分类,共确定了18个AdLOX和18个AiLOX基因。通过分子系统发育关系分析发现,AdLOX6、AdLOX9、AdLOX11、AdLOX12、AdLOX17、AiLOX1、AiLOX4、AiLOX6、AiLOX16和AiLOX17基因可能参与植物体的抗病过程,其中,直系同源基因的功能相对保守。AdLOX和AiLOX基因在进化过程中受到纯化选择作用。endprint
猜你喜欢
山地农业生物学报(2019年1期)2019-09-10
发明与创新·大科技(2019年5期)2019-07-31
湖北农业科学(2017年17期)2017-10-13
湖北农业科学(2017年16期)2017-09-14
农民致富之友(2017年10期)2017-06-15
农家科技下旬刊(2017年3期)2017-04-26
江苏农业科学(2016年12期)2017-04-05
山东农业科学(2014年5期)2014-07-18
湖北农业科学(2014年5期)2014-07-05
中学生物学(2008年3期)2008-06-03
