
饲料中维生素E水平对三疣梭子蟹幼蟹生长性能、血清非特异性免疫及抗氧化指标的影响
2015-12-21 05:33:38霍雅文侯迎梅周歧存
动物营养学报 2015年5期
李 弋 张 稳 金 敏 霍雅文 邱 红 侯迎梅 周歧存
(宁波大学海洋学院,鱼类营养研究室,宁波 315211)
维生素E又名生育酚,其与水生动物机体的代谢、抗氧化功能、免疫功能、抗病力和繁殖性能有着密切的联系。维生素E作为一种生物抗氧化剂,可预防饲料和机体组织中多种不饱和脂肪酸的氧化,并可促进虾蟹类成活和生长。水产动物对维生素E需求的研究目前主要集中在鱼类上,在甲壳类上的研究较少见报道。近年来,维生素E对水生动物免疫功能的影响已逐渐成为研究热点。动物免疫反应启动主要在细胞膜上发生,免疫系统为协助清除病原体而产生的不稳定自由基会破坏吞噬细胞,而维生素E可保护体细胞免遭自由基破坏,并使细胞膜维持正常功能[1]。饲料中补充维生素E能显著增强中国对虾血清中酚氧化酶(phenoloxidase,PO)的活性,并且随着维生素E水平的升高,PO活性增强。一定量的维生素E能促进酚氧化酶原(prophenoloxidase,proPO)的合成,增强中国对虾血清对副溶血弧菌和溶藻弧菌的吞噬活力[2]。Shiau 等[3]研究表明,饲料中维生素E水平为85~89 mg/kg时,斑节对虾生长最快、非特异性免疫力最强;维生素E水平为179 mg/kg时,组织中维生素E积累量最大。胡俊茹等[4]认为维生素E和硒对凡纳滨对虾抗氧化系统具有随时间变化的动态调节作用,维生素E和硒之间存在交互作用;当在基础饲料中维生素E和硒添加量分别在400、0.4 mg/kg时,凡纳滨对虾机体抗氧化能力整体达到平衡,能有效抵制氧自由基的损伤。维生素E除了促进生长、提高免疫力外,还可以促进繁殖和发育。吴桂玲等[5]研究表明,饲料中的维生素E与凡纳滨对虾的受精率、孵化率、幼体成活率以及仔虾的应激反应能力密切相关。在凡纳滨对虾亲体饲料中适量添加维生素E,能显著提高凡纳滨对虾的受精率、孵化率、幼体成活率及仔虾的应激反应能力。罗文等[6]认为饲料中维生素E水平为192 mg/kg时可明显提高红螯螯虾的繁殖性能,维生素E主要是通过提高红螯螯虾受精卵的质量来进一步提高胚胎发育的成活率和孵化率。林仕梅等[7]采用L9正交设计法,配制9种不同维生素C、维生素E、肌醇、胆碱水平的配合饲料喂养中华绒螯蟹,结果表明中华绒螯蟹配合饲料中维生素E适宜量为45 mg/kg。艾春香等[8]认为维生素E作为免疫刺激剂能有效地增强中华绒螯蟹的非特异性免疫能力,增强中华绒螯蟹免疫功能的维生素E适宜添加量为200~400 mg/kg。
三疣梭子蟹(Portunus trituberculatus)广泛分布于我国沿海,具有生长快、产量高、肉质好、环境适应能力强等优点,颇受广大消费者和养殖户的青睐,是我国重要的海产经济蟹类,在海洋捕捞和海水养殖业中占有极其重要的地位。长期以来,三疣梭子蟹养殖过程中多采用低值小杂鱼和贝类直接投喂,投喂生物饵料不仅造成三疣梭子蟹营养不平衡,而且还会破坏海洋渔业资源、污染水域环境、诱发三疣梭子蟹养殖病害发生等。因此,研制营养全面、优质高效的人工全价配合饲料已成为三疣梭子蟹产业迅速发展的迫切需要。目前有关蟹类的研究主要集中在能量蛋白质比、蛋白质、脂肪、粗纤维营养等主要营养素的营养需求方面[9-12],而有关维生素E营养的研究尚未见报道。本文以三疣梭子蟹幼蟹为研究对象,采用单体框养的方式,探讨饲料中维生素E水平对幼蟹生长性能以及血清中免疫相关指标和抗氧化相关指标的影响,为三疣梭子蟹高效配合饲料的研制开发提供理论依据。
1 材料与方法
1.1 试验饲料
本试验以秘鲁红鱼粉、豆粕、小麦蛋白粉和磷虾粉为蛋白质源,鱼油、大豆油和大豆卵磷脂为脂肪源,并补充矿物质和维生素预混料等以满足幼蟹的营养需求,配制基础饲料,在基础饲料基础上分别添加 0、10、20、40、80 和 160 mg/kg 的维生素E(帝斯曼公司生产的维生素E乙酸酯,纯度为50%),制成6种等氮等能的试验饲料,通过高效液相色谱检测饲料中维生素E水平分别为8.9、19.6、27.0、39.7、77.6 和 156.9 mg/kg。基础饲料组成及营养水平见表1。所有原料粉碎后过80目筛,分别称重、混匀后添加油脂和水,以双螺杆挤条机挤压并经饲料颗粒机制成2种规格的颗粒饲料(分别为直径3 mm、长度5 mm,直径5 mm,长度7 mm),饲料90℃熟化30 min,阴凉处风干,用封口袋封装后于-20℃下保存待用。
1.2 养殖管理
养殖试验在宁波象山县东陈乡鑫亿梭子蟹水产公司进行,试验用三疣梭子蟹幼蟹购自宁波兢业育苗厂,幼蟹置于水泥池中暂养。本试验采用单体筐养的方式,塑料筐规格为80 cm×40 cm×40 cm,养殖筐中间设置一隔板,隔板两边各放一装有沙子的沙盒,作为幼蟹的栖息场所,沙盒规格为25 cm×15 cm×10 cm。试验挑选360只初始体重为(5.94±0.17)g的幼蟹,随机分配于180只养殖筐中(2只/筐),试验筐随机分组,每种试验饲料投喂60只幼蟹,每20只幼蟹作为1个重复。试验开始前统一用未添加维生素E的试验饲料驯化1周,试验开始后每种饲料每天投喂2次(07:00、17:00),每天投喂量控制在幼蟹体重的6%~8%,每天早晨捞取残饵和蜕壳,记录死亡数、投饵量和蜕壳次数。所有养殖筐置于1个水泥池中(70 m×80 m×0.6 m),池底铺设充氧管不间断充氧,试验期间水温26~30℃,溶解氧浓度6.5 ~7.0 mg/L、氨氮浓度≤0.5 mg/L、pH 7.8~8.2、盐度26‰ ~28‰,2 ~3 d换水1 次,每次换水量约为30%,试验期为8周。
1.3 样品采集、分析与计算
试验结束时,清点每组存活的三疣梭子蟹的数量,计算成活率。停食24 h后取样,每组蟹子分别称重,并在每组中随机取4只蟹从第3步足抽取血液,分别放入1.5 mL离心管中,4℃静置过夜,3 000 r/min离心8 min,取上清,用于血清指标的测定。每组另随机取4只蟹进行常规营养成分分析。
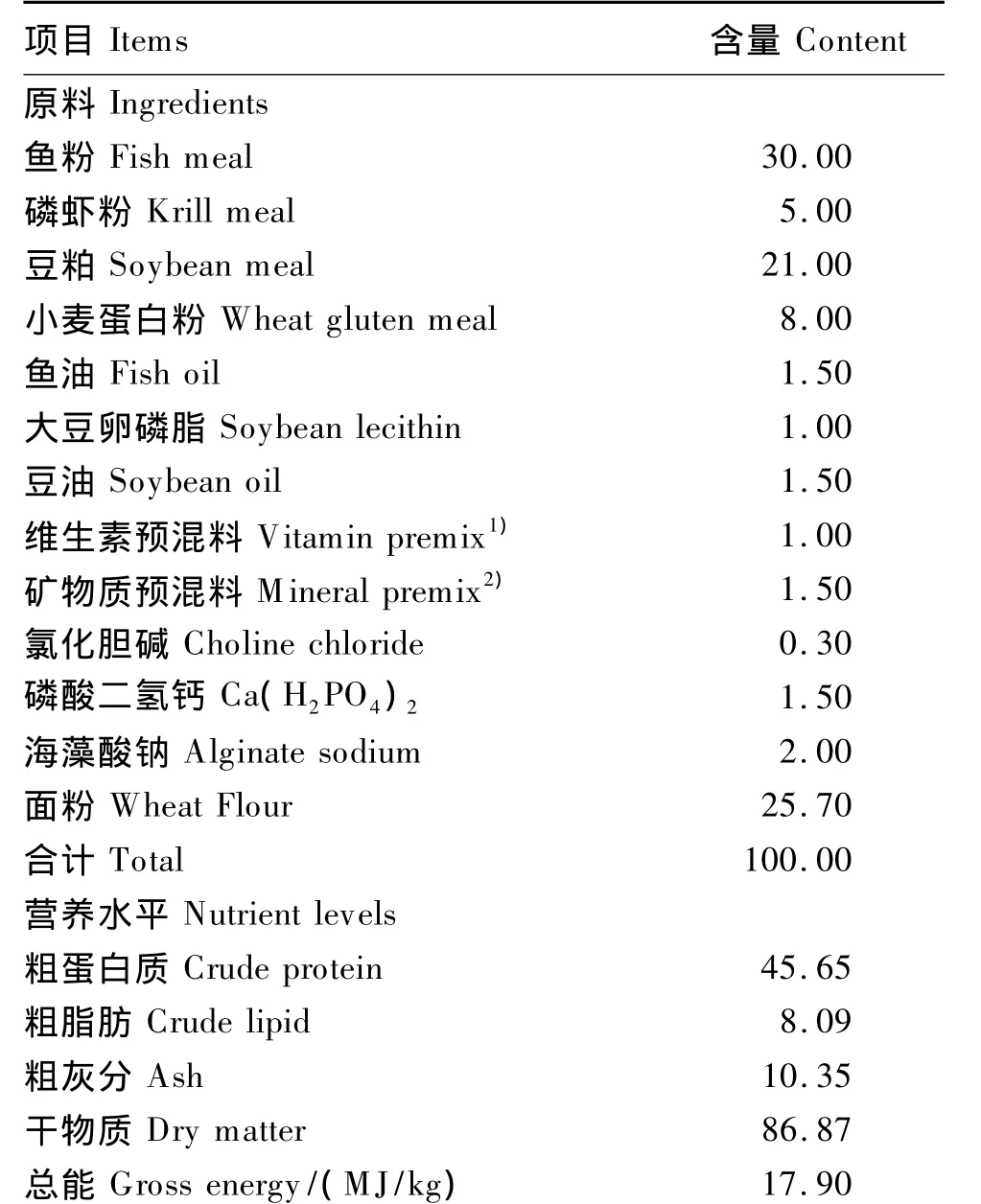
表1 基础饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of the basal diet(DM basis) %
1.3.1 生长指标
增重率(WGR,%)=100×(终末体重-初始体重)/初始体重;
特定生长率(SGR,%/d)=100×(ln终末体重-ln初始体重)/试验天数;
饲料系数(FCR)=饲料摄入量(干重)/幼蟹体增重(湿重);
蜕壳率(MR,%)=100×2×蜕壳次数/(开始时蟹个数+结束时蟹个数)[13];
成活率(SR,%)=100×开始时蟹个体数/结束时蟹个体数;
1.3.2 常规营养成分分析
用于常规营养成分分析的饲料和幼蟹样品置于105℃烘箱中至恒重,测定方法参照AOAC(1995)。水分含量由烘干前后样品的质量变化量计算而得;粗蛋白质含量采用凯氏定氮仪(FOSS,Tecator,Hoganas,Sweden)测定样品的总氮量后,通过总氮量乘以6.25而得;粗脂肪含量采用索氏抽提仪(Soxtec System HT6,Tecator,Sweden)测定;粗灰分含量采用高温灼烧法测定,将样品置于马福炉中550℃下灼烧8 h至恒重。所有指标均重复测定2次,若同一样品测定结果相对偏差大于2%,则增加重复数,采用相对偏差2%以内的2个测定值的平均值作为测定结果。
1.3.3 血清生化指标
血清中总蛋白(total protein,TP)、甘油三酯(triglyceride,TG)、葡萄糖(glucose,GLU)含量均在全自动生化分析仪(OLYMPUSAU 600,日本)上测定。
1.3.4 血清非特异性免疫指标
血清溶菌酶(lysozyme,LSZ)活性测定采用Hultmark等[14]的方法并加以改进:以溶壁微球菌冻干粉为底物,用0.1 mol/L的磷酸钾盐缓冲液(pH=6.4)配制成一定浓度的底物悬液(在570 nm处的吸光度值为0.3),在此法规定条件下,溶菌LSZ活性(U/mL)=(A0-A)/A(A0为反应之前菌悬液的吸光度,A为反应之后菌悬液的吸光度)。碱性磷酸酶活性(alkaline phosphatase,AKP)活性采用南京建成工程生物研究所试剂盒进行测定。
1.3.5 血清抗氧化指标
血清超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量采用南京建成工程生物研究所试剂盒进行测定。
1.4 数据统计与分析
采用SPSS 16.0对所有数据进行单因素方差(one-way ANOVA)分析,若组间差异显著(P<0.05),则进行Tukey多重比较。分析结果均用“平均值±标准误”表示。
2 结果
2.1 饲料中维生素 E水平对三疣梭子蟹幼蟹生长性能的影响
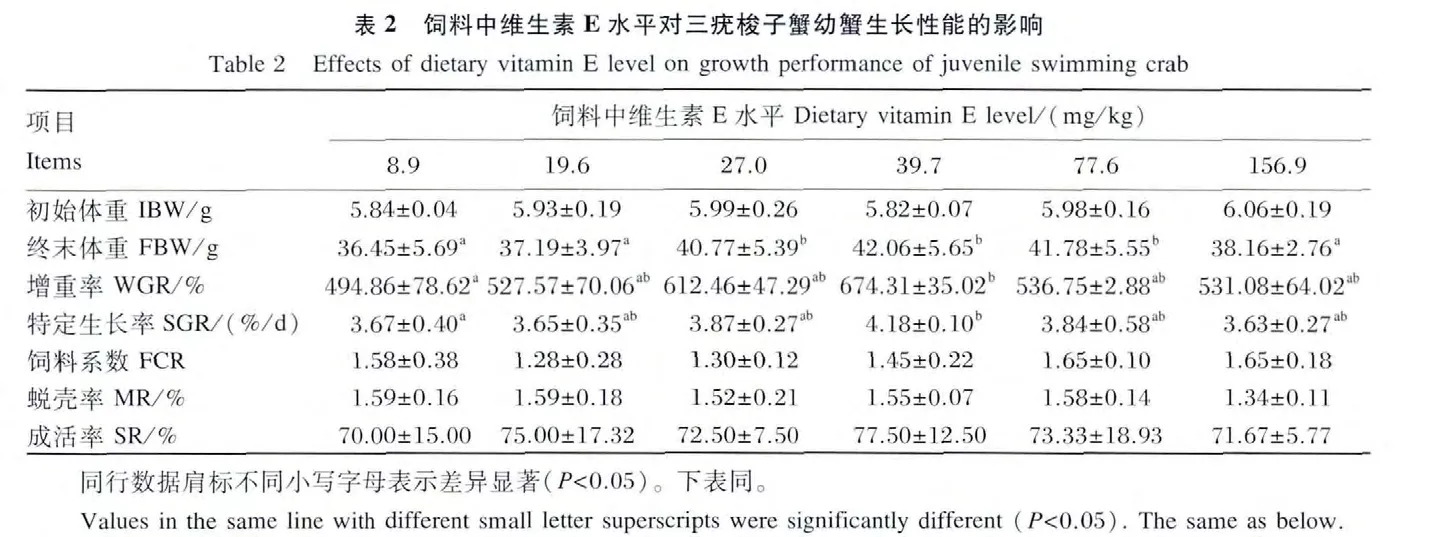
由表2可见,当饲料中维生素 E水平从8.9 mg/kg提高到39.7 mg/kg时,幼蟹的 WGR 和SGR显著提高,即39.7 mg/kg组的WGR和SGR显著高于8.9 mg/kg组(P<0.05);当饲料中维生素E水平超过39.7 mg/kg时,WGR和SGR不再增高,反而出现降低的趋势。以SGR为评价指标,得折线回归方程 y=0.017 6x+3.424 8(R2=0.857 6,x <39.0)和 y=0.004 4x+4.283 7(R2=0.890 8,x>39.0),由此得出三疣梭子蟹幼蟹的维生素E最适需求量为39.0 mg/kg(饲料干重,图1)。FCR由于组内差异较大,导致各组间差异不显著(P>0.05),但是19.6和27.0 mg/kg组要低于其他各组。饲料中维生素E水平对幼蟹的MR和SR均没有显著影响(P >0.05)。
?
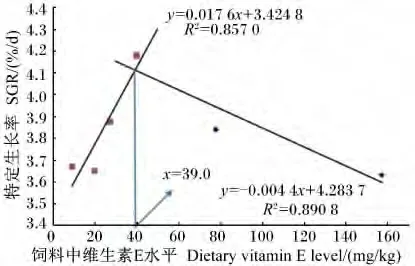
图1 饲料中维生素E水平与三疣梭子蟹幼蟹SGR的关系Fig.1 Relationship between the dietary vitamin E level and SGR of juvenile swimming crab
2.2 饲料中维生素E水平对三疣梭子蟹幼蟹全蟹常规营养成分的影响
由表3可见,饲料中维生素E水平对幼蟹全蟹水分、粗灰分和粗蛋白质含量的影响不显著(P>0.05),而对粗脂肪含量有显著影响(P<0.05)。当饲料中维生素 E水平从 8.9 mg/kg增加到39.7 mg/kg时,幼蟹全蟹粗脂肪含量逐渐降低,维生素E水平再增加到77.6 mg/kg时,幼蟹粗脂肪含量逐渐升高。
2.3 饲料中维生素E水平对三疣梭子蟹幼蟹血清生化指标的影响
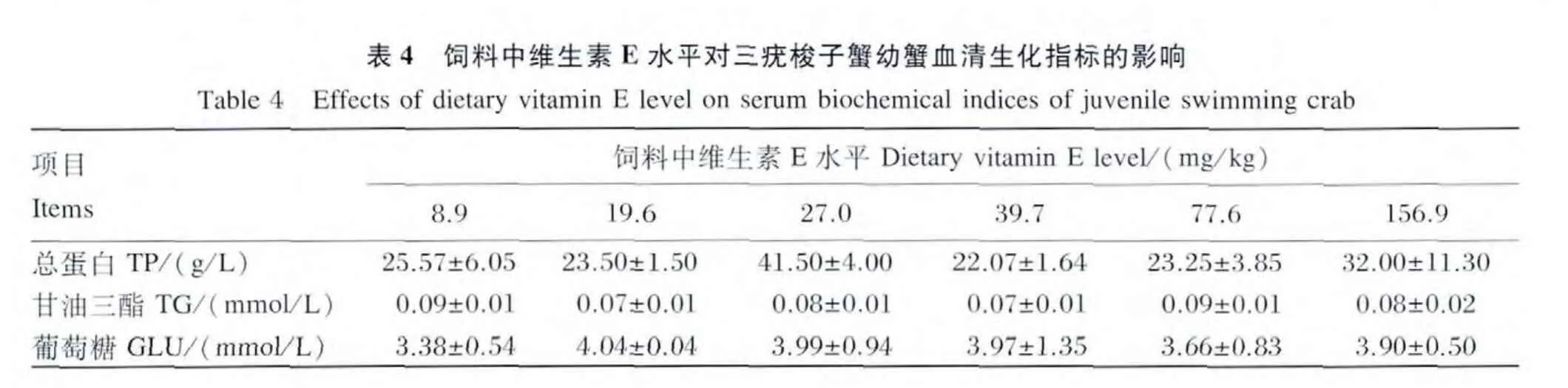
由表4可见,饲料中维生素E水平对幼蟹血清中TP、TG和 GLU含量均没有显著影响(P>0.05)。
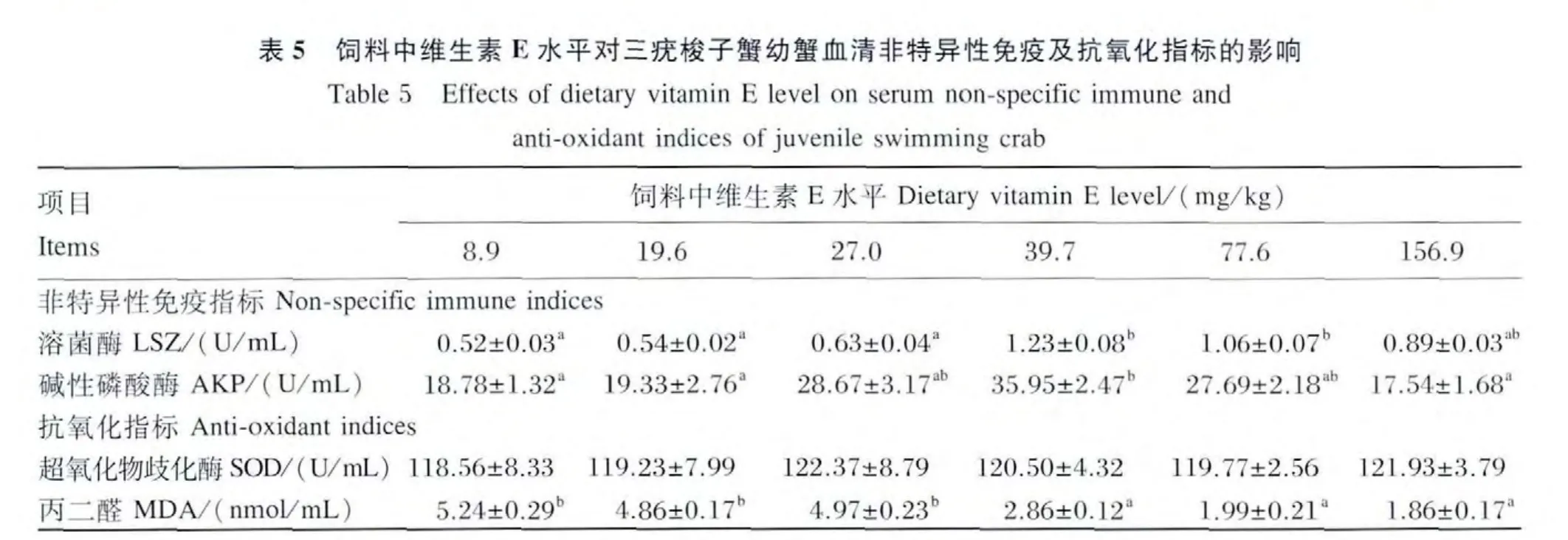
2.4 饲料中维生素E水平对三疣梭子蟹幼蟹血清非特异性免疫指标及抗氧化指标的影响
由表5可见,当饲料中维生素 E水平从8.9 mg/kg增加到39.7 mg/kg时,血清中 LSZ 活性逐渐升高,39.7 mg/kg 组显著高于8.9、19.6 和27.0 mg/kg组(P<0.05);当饲料中维生素E水平继续增加到77.6 mg/kg时,血清中LSZ活性不再升高,反而有降低的趋势。饲料中维生素E水平从8.9 mg/kg增加到39.7 mg/kg时,血清中 AKP活性逐渐升高,39.7 mg/kg组显著高于 8.9和19.6 mg/kg组(P <0.05);当饲料中维生素 E 水平继续增加到77.6 mg/kg时,血清中AKP活性不再升高,反而有降低的趋势。
由表5可见,饲料中维生素E水平对幼蟹血清中SOD活性没有显著影响(P>0.05)。血清中MDA含量随着饲料中维生素E水平的增加逐渐降低,当饲料中维生素 E水平增加到156.9 mg/kg时,血清中MDA含量最低,显著低于8.9 mg/kg组(P <0.05)。

?
?
?
3 讨论
3.1 饲料中维生素E水平对三疣梭子蟹幼蟹生长性能的影响
维生素E作为一种抗氧化剂,可预防饲料和机体组织中多不饱和脂肪酸的氧化,从而促进水生动物的成活和生长。Kanazawa[15]研究发现,饲料中添加维生素E能提高日本对虾幼体的SR。He等[16-17]研究表明,饲料中添加适量的维生素E可提高凡纳滨对虾的WGR和SR。陈四清等[18]认为在饲料中添加适量的维生素E有助于提高中国对虾对蛋白质的消化吸收,从而促进其生长。本试验结果表明,当饲料维生素E水平在8.9~39.7 mg/kg时随着饲料中维生素E水平的增加,三疣梭子蟹幼蟹的WGR、SGR显著上升,这一结果表明饲料中适宜水平的维生素E可以促进三疣梭子蟹幼蟹的生长;但并非饲料中维生素E水平越高越好,当维生素E水平超过39.7 mg/kg时,幼蟹的WGR、SGR不再显著上升。因此,本试验中,饲料中维生素 E水平为39.7 mg/kg时,三疣梭子蟹幼蟹的生长最好。这与林仕梅等[7]对中华绒螯蟹的研究结果一致,林仕梅等[7]以 SGR和FCR为指标,中华绒螯蟹对维生素E的适宜需求量为45 mg/kg。甲壳类动物对维生素的需求量受饲料中营养成分(如饲料脂肪与维生素E水平等)、养殖动物的生长阶段、生理状态以及养殖环境密切相关[19]。但是,周立斌等[20]研究表明,美国红鱼最佳生长性能的饲料维生素E添加量为102.8 mg/kg。陈骋等[21]以黄颡鱼幼鱼为研究对象,饲料中维生素E的添加量对幼鱼的特定生长率和饲料效率没有显著性影响,而添加量为175 mg/kg时,幼鱼摄食率显著降低。这可能与试验物种、饲养模式、维生素E添加形式和水质的不同有关。艾春香等[22]发现,饲料中添加维生素E 500 mg/kg相较于添加维生素E 300 mg/kg时,仔蟹的SR、WGR和蜕皮频率显著降低,这与本试验结果相符。
3.2 饲料中维生素E水平对三疣梭子蟹幼蟹全蟹常规营养成分的影响
很多矿物质和维生素是与脂类代谢相关酶的辅基和辅酶,参与体内的转甲基反应,因而对于脂肪代谢非常重要。维生素E、硒协同作用与脂类代谢关系极为密切,能防止并清除脂类代谢过程中产生的过氧化物[23]。因此,当饲料中维生素E不足时,动物体内的脂肪就不能作为能源有效地被利用,而是累积在体内,此时蛋白质就被消耗提供能量。周震等[24]研究了鲤鱼维生素E的缺乏症,结果表明,不添加维生素E组约有68%的鲤鱼发病,且病鱼肌肉水分的含量非常高,蛋白质含量有降低的趋势,肌肉纤维萎缩,鲤表现营养性肌病和白肌病等,而添加维生素E组则没有此现象发生。本试验结果表明,饲料中维生素E水平对三疣梭子蟹幼蟹全蟹粗脂肪含量有显著影响,在一定范围内,蟹体粗脂肪含量随着饲料中维生素E水平的增加而降低,说明维生素E可以促进幼蟹体内脂肪代谢,使脂肪积累量减少;但是随着饲料中维生素E水平进一步增加,蟹体粗脂肪含量升高,说明饲料中维生素E水平过量,抑制了体内脂肪代谢过程,从而使脂肪积累量增加。但是,本试验没有发现全蟹粗蛋白质含量降低的现象,这也许是由于本试验中未添加维生素E组(饲料中维生素E水平为8.9 mg/kg)饲料足以使体内的脂肪分解产生生长所需的能量,不需要再消耗体内的蛋白质提供能源。
3.3 饲料中维生素E水平对三疣梭子蟹幼蟹血清生化指标的影响
动物血液指标被广泛地用来评价其健康状况、营养状况及对环境的适应状况,是重要的生理、病理和毒理学指标[25-26]。血清TP含量反映鱼体的营养与代谢状况,也间接反映了机体的免疫水平的高低。不同程度的脂肪肝与肝内TG积聚的量有关。徐文[27]研究显示,高TG增加脂肪肝的患病率,而单纯的高胆固醇(CHOL)对脂肪肝的产生没有多大影响,脂肪肝与TG、CHOL等含量之间的关系还有待进一步的研究。血糖是反映动物糖代谢和全身组织细胞功能状态以及内分泌机能的一个重要指标。本试验的结果表明,饲料中维生素E水平对三疣梭子蟹幼蟹血清中TP、TG和GLU含量均没有显著影响。
3.4 饲料中维生素E水平对三疣梭子蟹幼蟹血清非特性免疫指标的影响
LSZ是吞噬细胞杀菌的物质基础,在甲壳类动物的免疫防御中起重要作用。它能够水解革兰氏阳性细菌的细胞壁,破坏入侵体内的异物,从而担负起机体防御的功能[28]。艾春香等[29]研究表明,维生素E添加量对中华绒螯蟹各组织、器官中PO活性、抗菌力和LSZ活性有显著或极显著的影响,在适宜范围内,随着维生素E添加量的增加,PO活性、抗菌力和LSZ活性显著增强,从而增强中华绒螯蟹的非特异性免疫力。周立斌[20]等研究表明,美国红鱼血清LSZ和血清总补体活性随着维生素E的添加量的增加而显著升高。本试验研究显示,饲料中适宜水平的维生素E,可以显著提高血清中LSZ活性,但当维生素E水平增加到一定程度后,血清中LSZ活性降低。这与以上的研究结果相一致。
AKP可催化所有的磷酸单酯及磷酸基团的转移反应,它直接参与磷的代谢,亦与 DNA、RNA、蛋白质、脂质等代谢有关,它对钙质的吸取、磷酸钙沉积、骨骼形成、甲壳素分泌及形成都有重要的作用;虾蟹类等甲壳动物在生长过程中都要经历蜕壳过程,该酶对虾蟹类的生存、生长有特别重要的意义[30]。艾春香等[31]研究表明,中华绒螯蟹血清AKP活性随着维生素E含量的增加而显著升高。本试验研究显示,饲料中维生素E水平在一定范围内增加,血清中AKP活性会逐渐升高,但当维生素E水平超过一定的量后血清中AKP活性又开始降低。可见,饲料中适宜水平的维生素E可以显著提高三疣梭子蟹的非特异性免疫力,但当维生素E水平进一步增加后三疣梭子蟹的非特异性免疫力反而下降。
3.5 饲料中维生素E水平对三疣梭子蟹幼蟹血清抗氧化指标的影响
维生素E之所以具有抗氧化作用是由于它容易被氧化,可以代替其他物质首先被氧化,因而可以起到保护其他物质不被氧化的作用。维生素E本身具有产生酚氧化基的结构,产生的酚氧基能够猝灭并能同单线态氧反应,保护不饱和脂肪酸免受单线态氧损伤,还可以被超氧阴离子自由基和羟自由基氧化,使不饱和脂肪酸免受自由基攻击,从而抑制脂肪酸的自动氧化[32]。SOD是生物体内一种重要抗氧化防御酶,其基本功能是清除由代谢产生的活性氧,控制自由基引起质膜过氧化[33]。当虾蟹类机体受到轻度逆境胁迫时,SOD活性被诱导,而受重度逆境胁迫时,其活性则被抑制。SOD活性可以作为虾蟹类免疫反应的一个重要指标[34-35]。MDA是膜脂过氧化的最终分解产物,其含量可以反映机体遭受过氧化损害的程度。本试验结果显示,随着饲料中维生素E水平的增加,三疣梭子蟹幼蟹血清中SOD活性未发生显著变化,血清中MDA含量随着饲料中维生素E水平的增加而逐渐降低。艾春香等[31]研究表明,饲料中维生素E添加量对中华绒螯蟹组织SOD活性有显著影响,这与本试验的试验结果并不相符。这可能与试验动物的生长阶段不同有关,由于幼蟹SOD活性显著低于成蟹[36],造成了幼蟹SOD活性变化并不明显。王立改[37]研究表明,维生素E添加量为100 mg/kg饲料组幼蟹肝胰腺MDA含量显著低于维生素E添加量为0、300 mg/kg饲料组,这与本试验结果相符。这表明在饲料中添加维生素E有助于提高三疣梭子蟹的抗氧化能力。
4 结论
综上,本研究结果表明饲料维生素E水平影响三疣梭子蟹的生长性能、血清非特异性免疫及抗氧化指标,基于SGR的折线模型拟合后获得三疣梭子蟹幼蟹对维生素E的适宜需求量为39.0 mg/kg(饲料干重)。
[1] LIKOFF R O,GUPTILL D R,LAWRENCE L M,et al.Vitamin E and aspirin depress prostaglandins in protection of chickens against Escherichia coli infection[J].The American Journal of Clinical Nutrition,1981,34(2):245-251.
[2] 王伟庆,李爱杰.LAPP对中国对虾(Penaeus chinensis)生长、缺氧耐受力及免疫抵抗力的影响[J].海洋湖沼通报,1996(1):42-49.
[3] LEE M H,SHIAU S Y.Vitamin E requirements of juvenile grass shrimp,Penaeus monodon,and effects on non-specific immune responses[J].Fish & Shellfish Immunology,2004,16(4):475-485.
[4] 胡俊茹,王安利,曹俊明.维生素E和硒互作对凡纳滨对虾(Litopenaeus vannamei)抗氧化系统的调节作用[J].海洋与湖沼,2010,41(1):68-74.
[5] 吴桂玲,梁萌青,常青,等.维生素A、E对凡纳滨对虾受精率、孵化率及幼体存活率的影响[J].海洋水产研究,2008,29(4):58-62.
[6] 罗文,王群,赵云龙,等.维生素E对红螯螯虾(Cherax quadricarinatus)繁殖性能的影响[J].海洋与湖沼,2005,36(4):335-342.
[7] 林仕梅,叶元土,罗莉,等.中华绒螯蟹对 VC、VE、肌醇和胆碱需要量的研究[J].饲料工业,2000,21(8):21-23.
[8] 艾春香,林琼武,李少菁,等.蟹类的营养需求研究及其配合饲料研制[J].厦门大学学报:自然科学版,2006,45(增刊 2):205-212.
[9] 高红建,何中央,丁雪燕,等.三疣梭子蟹幼蟹配合饲料中蛋白质、脂肪、粗纤维适宜含量研究[J].渔业现代化,2009,36(1):26-29.
[10] CATACUTAN M R.Growth and body composition of juvenile mud crab,Scylla serrata,fed different dietary protein and lipid levels and protein to energy ratios[J].Aquaculture,2002,208(1/2):113-123.
[11] HUO Y W,JIN M,ZHOU P P,et al.Effects of dietary protein and lipid levels on growth,feed utilization and body composition of juvenile swimming crab,Portunus trituberculatus[J].Aquaculture,2014,434:151-158.
[12] SHEEN SS,WU SW.The effects of dietary lipid levels on the growth response of juvenile mud crab Scylla serrata[J].Aquaculture,1999,175(1/2):143-153.
[13] 陈立侨,堵南山,赖伟.中华绒螯蟹蟹种配饵中豆饼替代部分鱼粉的适宜含量[J].水产学报,1994,18(1):24-31.
[14] HULTMARK D,STEINER H,RASMUSON T.Insect immunity.Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia[J].European Journal of Biochemistry,1980,106(1):7-16.
[15] KANAZAWA A.Nutrition of penaeid prawns and shrimps[C]//TAKI Y,PRIMAVERA J H,LLOBRERA J A.Proceedings of the First International Conference on the Culture of Penaeid Prawns/Shrimps,4-7 December 1984,Iloilo City,Philippines.Iloilo City,Philippines:Aquaculture Department,Southeast Asian Fisheries Development Center,1985:123-130.
[16] HE H,LAWRENCE A L,LIU R Y.Evaluation of dietary essentiality of fat-soluble vitamins,A,D,E and K for penaeid shrimp(Penaeus vannamei)[J].Aquaculture,1992,103(2):177-185.
[17] HE H Q,LAWRENCE A L.Vitamin E requirement of Penaeus vannamei[J].Aquaculture,1993,118(3/4):245-255.
[18] 陈四清,李爱杰.中国对虾对维生素E,K营养需要的研究[J].海洋科学,1993(5):1-4.
[19] HE H Q,LAWRENCE A L.Vitamin C requirements of the shrimp Penaeus vannamei[J].Aquaculture,1993,114(3/4):305-316.
[20] 周立斌,王安利,张伟,等.饲料维生素E含量对美国红鱼生长和非特异性免疫的影响[J].渔业科学进展,2009,30(1):47-53.
[21] 陈骋,熊晶,左永松,等.饲料中不同维生素E添加量对黄颡鱼幼鱼生长性能及免疫功能的影响[J].中国水产科学,2010,17(3):521-526.
[22] 艾春香,陈立侨,温小波.中华绒螯蟹仔蟹维生素E营养需求研究[J].海洋科学,2002,26(11):1-5.
[23] SUÁREZ A,RAMÍREZ-TORTOSA M,GIL A,et al.Addition of vitamin E to long-chain polyunsaturated fatty acid-enriched diets protects neonatal tissue lipids against peroxidation in rats[J].European Journal of Nutrition,1999,38(4):169-176.
[24] 周震.鲤鱼维生素E缺乏的病理学研究[D].硕士学位论文.雅安:四川农业大学,2004.
[25] 周玉,郭文场,杨振国,等.鱼类血液学指标研究的进展[J].上海水产大学学报,2001,10(2):163-165.
[26] BAHMANI M,ORYAN S,POURKAZEMI M,et al.Effects of ecophysiological stress on cellular immunity system of Persian sturgeon Acipenser persucus[R].Tehran:14th Iranian Congress of Physiology and Pharmacology,1999.
[27] 徐文.高血脂症与脂肪肝的关系探讨[J].医学信息,2010,23(7):2098-2099.
[28] 陈竞春,石安静.贝类免疫生物学研究概况[J].水生生物学报,1996,20(1):74-78.
[29] 艾春香,陈立桥,刘晓玲,等.维生素E对中华绒螯蟹(Eriocheir sinensis)酚氧化酶、抗菌力和溶菌酶活性的影响[J].海洋与湖沼,2008,39(2):119-123.
[30] 陈四清,陈素丽,石艳,等.长毛对虾碱性磷酸酶性质[J].厦门大学学报:自然科学版,1996,35(2):257-261.
[31] 艾春香,陈立桥,温小波,等.VE对河蟹血清和组织中超氧化物歧化酶及磷酸酶活性的影响[J].台湾海峡,2003,22(1):24-31.
[32] 郭红菊.维生素E功能的研究进展[J].天水师范学院学报,2005,25(5):44-46.
[33] FRIDOVICH I.Superoxide radical and superoxide dismutases[J].Annual Review of Biochemistry,1995,64(1):97-112.
[34] CAMPA-CÓRDORA A I,HEMÁNDEZ-SAAVEDA N Y,DE PHISLIPPIS R,et al.Generation of superoxide anion and SOD activity in haemocytes and muscle of American white shrimp(Litopenaeus vannamei)as a response to β-glucan and sulphated polysaccharide[J].Fish & Shellfish Immunology,2002,12(4):353-366.
[35] CAMPA-CÓRDORA A I,HEMÁNDEZ-SAAVEDA N Y,ASCENCIO F.Superoxide dismutase as modulator of immune function in American white shrimp(Litopenaeus vannamei)[J].Comparative Biochemistry and Physiology Part C:Toxicology & Pharmacology,2002,133(4):557-565.
[36] 江水恒,徐善良,邵波,等.野生和养殖三疣梭子蟹消化酶及 SOD酶活性的比较分析[J].海洋渔业,2012,34(2):164-167
[37] 王立改.维生素E对中华绒螯蟹饲料脂肪利用的保护作用研究[D].博士学位论文.上海:华东师范大学,2014.
猜你喜欢
今日农业(2022年15期)2022-09-20 06:55:36
当代水产(2022年3期)2022-04-26 14:26:58
现代畜牧科技(2021年8期)2021-10-13 07:21:58
中老年保健(2021年5期)2021-08-24 07:07:02
保健医苑(2020年1期)2020-07-27 01:58:14
童话世界(2020年8期)2020-06-15 11:32:40
新世纪智能(英语备考)(2019年11期)2020-01-18 11:49:48
海峡姐妹(2018年11期)2018-12-19 05:18:30
北京广播电视报(2018年39期)2018-11-23 06:01:14
现代营销·经营版(2017年3期)2017-05-17 10:19:46
