
杜鹃红山茶的光响应特性及其最适模型筛选
2015-12-08 02:43张方秋杨会肖徐斌潘文
生态环境学报 2015年10期
张方秋,杨会肖,徐斌,潘文
广东省森林病虫害生物防治重点实验室,广东 广州 510520;广东省林业科学研究院,广东 广州 510520
杜鹃红山茶的光响应特性及其最适模型筛选
张方秋,杨会肖,徐斌*,潘文
广东省森林病虫害生物防治重点实验室,广东 广州 510520;广东省林业科学研究院,广东 广州 510520
杜鹃红山茶(Camellia azalea)是我国特有的山茶原生种,仅见于广东省阳春市鹅凰嶂自然保护区内,其树形优美、四季开花,在园林绿化中具有广泛的应用。鉴于当前对杜鹃红山茶光响应曲线模型的适合性尚未见报道,本研究选用二次多项式回归模型、直角双曲线模型、非直角双曲线模型、直角双曲线修正模型、指数模型和指数改进模型对杜鹃红山茶叶片光响应曲线进行拟合,探讨不同光响应模型对杜鹃红山茶光合特性的适用性,筛选最佳光响应模型。结果表明,杜鹃红山茶叶片光合速率与光强之间存在非线性关系,直角双曲线修正模型的拟合效果最佳;对光响应曲线实测值与拟合值比较,发现二次多项式模型的拟合值与实测值相差较大,而其余5种模型的拟合值与实测值较为接近,但只有直角双曲线修正模型的拟合曲线出现光抑制现象;在各拟合曲线的光合参数中,只有直角双曲线修正模型拟合的Pmax、Isat、Ic、Rd值与实测值较吻合,相关系数R2值最大,为0.998;利用直角双曲线修正模型拟合出杜鹃红山茶叶片最大净光合速率(Pmax)为5.56 μmol·m-2·s-1,饱和光强(Isat)为1 327.30 μmol·m-2·s-1,光补偿点(Ic)为29.44 μmol·m-2·s-1,暗呼吸速率(Rd)为1.909 μmol·m-2·s-1,初始量子效率(α)为0.065,光合参数值表明,杜鹃红山茶具有较强的光能利用能力,符合阳生喜光植物的特性,同时又具有一定的耐荫能力,对光照的适应范围较宽。
杜鹃红山茶;光响应曲线模型;光合速率;光强
杜鹃红山茶(Camellia azalea),是我国特有的山茶科频危植物,属林下灌木至小乔木类型,主要分布在广东省阳春市鹅凰嶂省级自然保护区内。杜鹃红山茶树形优美、叶片光亮碧绿、花色艳丽密集、四季开花不断,且抗性强,能在强阳光和高温条件下开花和生长,冬季能够抵抗-5 ℃的低温,叶片厚革质,病虫害较少,在园林与观赏园艺方面具有广阔的应用前景(张宏达,1999;张方秋等,2012)。杜鹃红山茶作为一种喜光而耐半阴植物,对其光合生理特性方面的研究较少,主要有刘玉玲等人(2011)对其光合作用日变化特性,刘红晓等人(2013)对其光合作用特征的研究,而对杜鹃红山茶光响应曲线模型的适用性研究尚未见报道。因此本文以嫁接的4年生杜鹃红山茶为研究对象,应用当前最常用的二次多项式回归模型(郭水良等,2004)、直角双曲线模型(Cannell et al.,1998)、非直角双曲线模型(Dias-Filho,2002)、直角双曲线修正模型(Ye et al.,2008)、指数模型(Potvin et al., 1990)和指数改进模型(陈卫英等,2012)对其光响应曲线进行拟合研究,通过比较分析拟合的光响应曲线和光合参数,筛选适合模拟杜鹃红山茶光响应曲线的最优模型,确定其光响应特性,为杜鹃红山茶资源的引种栽培及园林应用提供科学依据。
1 材料与方法
1.1 试验材料
试验材料为广东省林业科学研究院苗圃内嫁接的4 a生杜鹃红山茶无性系植株,选择3株长势优良的无性系,每株选取3片植株中上部当年生向阳成熟叶片进行光响应曲线测定。
1.2 光响应曲线的测定
光响应数据用美国生产的Li-6400便携式光合测定仪(Li-Cor Inc.,USA)测定,在2014年9月,选择晴朗无风(微风)的天气,早上 9:00─11:30之间进行。为保持其它环境因子的稳定,测定时,叶温设为(28±0.5) ℃,空气相对湿度为60%左右,叶室中CO2浓度为400 μmol·mol-1,用Li-6400的
02B-LED红蓝光源进行手动设置光合有效辐射强度(分别为2000、1800、1600、1400、1200、1000、800、600、400、200、100、50、20、0 μmol·m-2·s-1),重复测定10次,取其平均值。
1.3 数据处理
利用SPSS 19.0软件和Excel进行数据处理和光响应曲线的拟合和作图。所采用的模型分述如下:
二次多项式回归模型(郭水良等,2004):

式中,a、b、c为待定参数,Pn为净光合速率(μmol·m-2·s-1),I为光合有效辐射强度(光强)(μmol·m-2·s-1)。二次多项式模型在光响应曲线研究中仅作为一个纯数学模型来应用,根据二次多项式的特性可以求出光饱和点(Isat)、光补偿点(Ic)、最大净光合速率(Pmax)、暗呼吸速率(Rd)。
直角双曲线模型(Cannell et al.,1998):
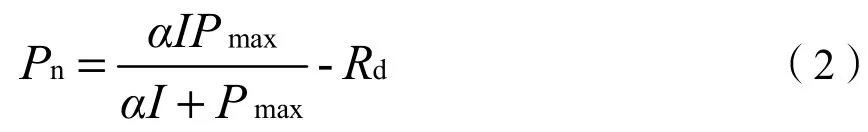
式中,Pn为净光合速率(μmol·m-2·s-1),I为光合有效辐射强度(μmol·m-2·s-1),α为植物光响应曲线在I=0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率,Pmax为光饱和时的最大净光合速率(μmol·m-2·s-1),Rd为暗呼吸速率(μmol·m-2·s-1)。直角双曲线模型是一个没有极值的函数,无法由上式直接求出植物的Pmax和Isat的解析解,因此,只能利用直线方程拟合弱光下(I≤200 μmol·m-2·s-1)的光响应数据得到(刘宇锋等,2005)。
非直角双曲线模型(Dias-Filho,2002):

式中,Pn、Pmax、I、α、Rd的含义与直角双曲线模型相同,θ为非直角双曲线的凸度,取值0≤θ≤1。非直角双曲线模型同样是一个没有极值的函数,无法求出Isat的解析解,估算Isat的方法与直角双曲线模型的方法相同。
直角双曲线修正模型(Ye et al.,2008):

式中,Pn、I、α的定义同前,Ic为光补偿点,β为修正系数,γ为一个与光强无关的系数,γ=α/Pmax。直角双曲线修正模型是一个具有极值的函数,由此式直接求出Isat、Ic、Pmax的解析解。
指数模型(Potvin et al.,1990):

式中,Pn、Pmax、I、α、Rd的定义同前,e为自然对数的底。指数函数方程是一个没有极值的函数,无法求取Isat,要估算饱和光强,需假设光合速率为 0.9Pmax或 0.99Pmax所对应的光强为饱和光强(黄红英等,2009)。
指数改进模型(陈卫英等,2012):

式中,Pn、I的定义同前,α、β和ε为修正系数,e为自然对数的底。当I=0时,Rd=α-γ,当Pn=0时,光补偿点任意光强下方程的导数为当'nP=0,说明 Pn存在最大值,光饱和光强光饱和点的

表1 杜鹃红山茶光合作用光响应曲线测量值及拟合值Table 1 The measured and estimated data of photosynthesis light-response curve of C. azalea
2 结果与分析
2.1 光响应曲线的拟合及其比较
从表1的光响应测量数据及各模型的拟合值可
以看出,二次多项式模型的拟合值与实测值相差较大,特别是在光强≤600 μmol·m-2·s-1和光强≥1600 μmol·m-2·s-1时,且当I为0 μmol·m-2·s-1时,出现Pn为正值的错误结果,其余5种模型的拟合值与实测值较为接近。根据各模型拟合的光响应曲线可知(图 1),直角双曲线模型、非直角双曲线模型、指数模型与指数改进模型拟合曲线都没出现光饱和区域的光抑制现象,在强光下曲线仍呈上升趋势。二次多项式模型拟合曲线虽然有光饱和点,但不能较好的解释光强超过此计算所得Isat后,Pn值快速下降的原因。相反,直角双曲线修正模型的拟合曲线体现出杜鹃红山茶光合作用光抑制现象,即达到光饱和点后,净光合速率随光强的增加而缓慢降低。

图1 6种模型拟合的杜鹃红山茶光合-光响应曲线Fig. 1 Light-response curve of C. azalea fitted by six models

表2 6种模型拟合得到的光响应曲线参数及其测量值Table 2 The estimated photosynthetic parameters fitted by six models and Corresponding measured data of C. azalea
2.2 6种光响应模型拟合的杜鹃红山茶光响应曲线参数比较
利用上述的6种模型分别拟合杜鹃红山茶光响应实测数据得到的光合参数见表 2。从结果可以看出,通过直角双曲线修正模型和二次多项式模型拟合的Pmax值与实测值较为接近,其余4个模型的拟合值则与实测值相差较大,特别是直角双曲线、非直角双曲线以及指数模型的 Pmax值明显大于实测值;就 Isat而言,直角双曲线修正模型和二次多项求解得到的 Isat值与实测值较为接近,这个值与刘红晓等人(2013)所测结果基本一样,而指数改进模型、直角与非直角双曲线模型通过直线方程求解得到的 Isat则远小于实测值,此外,根据以往的经验,指数方程按0.9Pmax或0.99Pmax假设仍无法求出Isat;通过二次多项式拟合得到的Ic值是负值,不符合生物学常识,而用其它模型拟合的 Ic值都在 30 μmol·m-2·s-1左右,稍大于实测值,其中直角双曲线、直角双曲线修正模型拟合值最小;在暗呼吸速率Rd拟合中,指数模型和指数改进模型拟合的Rd值都小于实测值,二次多项式拟合值为负值,这明显也是错误的,而直角双曲线、直角双曲线修正和非直角双曲线模型的拟合值与实测值较为接近;对于初始量子效率α来说,二次函数不能算出具有合适意义的 α值,其它模型拟合值差异较大,但都在0<α<0.125范围内,符合生物学规律值,其中,直
角双曲线模型拟合的α值为0.097,接近理论上限,数值有点偏大;从拟合度可以看出,除二次多项式拟合度较低外,其余模型的拟合度都较高,决定系数R2值均在0.990以上,因此,通过对比不同模型拟合的光合参数可以看出,R2值越大只能说明该模型拟合度较高,但并不能保证拟合结果就一定与实测值相符,在本研究中,只有直角双曲线修正模型估算的各项光合参数值与实测值均较为吻合。
综上所述,采用直角双曲线修正模型对于拟合杜鹃红山茶光合作用光响应曲线最为合适。由该拟合模型求得的杜鹃红山茶光饱和点(1327.299 μmol·m-2·s-1)较高,这说明该物种具有较强的光能利用能力,符合阳生喜光植物的特性。同时,其光补偿点(29.447 μmol·m-2·s-1)又较低,表明其在较弱光照环境下仍能进行有机物合成,具有一定的耐荫能力,且对光照的适应范围较宽。此外,杜鹃红山茶的初始量子效率α值(0.065)在0.04~0.07之间(户桂敏等,2009),也说明其对光照的适应范围较宽。
3 讨论与结论
3.1 讨论
通过植物光响应曲线估算的光合速率、初始量子效率、光饱和点、光补偿点等光合参数是植物重要的生态生理参数,因此,确定植物光合作用的光响应曲线对于研究植物的光合特性具有重要意义。前期相关研究表明,由于不同模型的意义不完全相同,不同的植物使用不同的模型其拟合度和与实测值的匹配存在一定的差异,因此对植物光响应曲线模型的适用性分析也是十分必要(叶子飘等,2008;Chen et al.,2011;Koyama et al.,2010)。在本研究中,除了二次多项式拟合的相关度较差外(R2=0.839),其余各模型的拟合相关度较高,均大于0.990,特别是直角双曲线修正模型拟合的杜鹃红山茶的光响应数据决定系数高达0.998。有较多的报道表明,直角双曲线与非直角双曲线模型拟合Pmax值较实测值偏大,且曲线没有光抑制情况(王帅等,2014;Peek et al.,2002;廖小锋等,2012;段爱国等,2009)。本研究也表明,采用直角双曲线、非直角双曲线进行光响应拟合时,最大净光合速率(Pmax)比实测值高出许多,而饱和光强却远小于实测值,同时,由于没有极值的特性使得这2个模型都无法处理植物在达到饱和光强后,净光合速率随光强的增强而降低的光响应问题;采用二次多项式拟合结果与其在其它植物光合生理相关研究一样(廖小锋等,2012;闫小红等,2013;王秀伟等,2009),虽然最大净光合速率、光饱和点与实测值最为接近,可以处理植物光抑制条件下的光响应问题,但存在光补偿点和暗呼吸速率为负值及光强达到光饱和点后,净光合速率快速下降的问题;指数函数拟合结果与其它类似研究一样,最大净光合速率和呼吸速率与实测值相差较大,且无法处理光抑制的问题,此外,通过以往经验仍无法求出假定0.99Pmax所对应的饱和光强。在指数改进模型的拟合中,虽然拟合度较高,能直接求出最大净光合速率与光饱和点,但拟合值与实测值相差较大。
直角双曲线修正模型在拟合杜鹃红山茶光响应曲线时,不仅能直接求取最大净光合速率、光饱和点等光合参数,而且求取的参数拟合值与实测值更为接近,同时也解决了光响应数据的光抑制现象,在达到饱和光强时,净光合速率随光强的增加而降低。因此,无论从模型的拟合度及其拟合值来看,在杜鹃红山茶光响应数据拟合中,直角双曲线修正模型的拟合效果都优于本研究中其它的应用模型,克服了其它模型拟合植物的光合参数时出现的上述问题,完全适用于杜鹃红山茶光响应曲线的拟合。有研究表明,此模型也适合于桉树、油茶、水稻和雷公藤等植物光响应的研究(侯智勇等,2009;洪伟等,2012;佘诚棋等,2012;叶子飘,2007)。
综上表明,在研究植物光响应曲线时应选用合适的模型进行分析,而衡量一种模型对植物光响应数据拟合是否适合,除了根据拟合的决定系数外,还应考虑光合参数模拟值与实测值的差异,同时依据植物自身的生长规律等进行判断。
3.2 结论
使用不同数学模型对杜鹃红山茶叶片光响应曲线的拟合值与实测值的之间差异较大,直角双曲线修正模型拟合效果最佳,该模型对杜鹃红山茶叶片光响应曲线的拟合值与实测值较为接近,相关系数R2值达0.998,其拟合的光合参数Pmax、Isat、Ic、Rd值与实测值较吻合,能较好解决强光下的杜鹃红山茶光抑制现象,是杜鹃红山茶光响应特性拟合的最适模型;杜鹃红山茶光饱和点(1327.299 μmol·m-2·s-1)较高,具有较强的光能利用能力,符合阳生喜光植物的特性。同时,其光补偿点(29.447μmol·m-2·s-1)又较低,具有一定的耐荫能力,对光照的适应范围较宽。
CANNELL M G R, THORNLEY J H M. 1998. Temperature and CO2responses of leaf and canopy photosynthesis: a clarification using the non-rectangular hyperbola model of photosynthesis [J]. Ann Bot, 82(6): 883-892.
CHEN Z Y, PENG Z S, YANG J, et al. 2011. A mathematical model for describing light-response curves in Nicotiana tabacum L. [J].
Photosynthetica, 49(3): 467-471.
DIAS-FILHO M B. 2002.Photosynthetic light response of the C4grasses Brachiaria brizantha and B. humidicola under shade [J]. Scientia Agricola, 59(1): 65-68.
KOYAMA K, KIKUZAWA K. 2010. Geometrical similarity analysis of photosynthetic light response curves, light saturation and light use efficiency [J]. Oecologia, 164(1): 53-63
PEEK M S, RUSSEK-COHEN E, WAIT D A, et al. 2002. Physiological response curve analysis using nonlinear mixed models [J]. Oecologia, 132(2): 175-180.
YE Z P, YU Q. 2008. A coupled model of stomatal conductance and photosynthesis for winter wheat [J]. Photosynthetica, 46(4): 637-793.
POTVIN C, LECHOWICZ MJ, TARDIE S. 1990. The statistical analysis of ecophysiological response curves obtained from experiments involving repeated measures [J]. Ecology, 71(4): 1389-1400.
陈卫英, 陈真勇, 罗辅燕, 等. 2012. 光响应曲线的指数改进模型与常用模型比较[J]. 植物生态学报, 36(12): 1277-1285.
段爱国, 张建国. 2009. 光合作用光响应曲线模型选择及低光强属性界定[J]. 林业科学研究, 22(6): 765-771.
郭水良, 方芳, 黄华, 等. 2004. 外来入侵植物北美车前繁殖及光合生理生态学研究[J]. 植物生态学报, 28(6): 787-793.
洪伟, 杨细明, 林勇明, 等. 2012. 不同光合光响应模型在雷公藤研究中的应用[J]. 福建农林大学学报(自然科学版), 41(1): 29-33.
侯智勇, 洪伟, 李健, 等. 2009. 不同桉树无性系光响应曲线研究[J]. 福建林学院学报, 29(2): 97-102.
户桂敏, 王文天, 彭少麟. 2009. 不同氮磷比下入侵种五爪金龙和本地种鸭脚木的竞争表面[J]. 生态环境学报, 18(4): 1449-1454.
黄红英, 窦新永, 孙蓓育, 等. 2009. 两种不同生态型麻疯树夏季光合特性的比较[J]. 生态学报, 29(6): 2861-2867.
廖小锋, 刘济明, 张东凯, 等. 2012. 野生小蓬竹的光合光响应曲线及其模型拟合[J]. 中南林业科技大学学报, 32(3): 124-128.
刘红晓, 袁莲莲, 刘楠, 等.2013. 杜鹃红山茶的扦插苗、嫁接苗和砧木的光合作用特征[J]. 生态环境学报, 22(6): 996-1000.
刘宇锋, 萧浪涛, 童建华, 等. 2005. 非直角双曲线模型在光合光响应曲线数据分析中的应用[J]. 中国农学通报, 21(8): 76-79.
刘玉玲, 刘信凯, 谌光晖, 等. 2011. 杜鹃红山茶与3个山茶栽培光合日变化特性研究[J]. 广东林业科技术, 27: 17-22.
佘诚棋, 程鹏, 季琳琳, 等. 2012. 油茶光合作用光响应曲线的拟合[J].经济林研究, 30(1): 118-121.
王帅, 韩晓日, 战秀梅, 等. 2014. 不同氨肥水平下玉米光响应曲线模型的比较[J]. 植物营养与肥料学报, 20(6): 1403-1412.
王秀伟, 毛子军. 2009. 7个光响应模型对不同植物种的实用性[J]. 植物研究, 29(1):43-48.
闫小红, 尹建华, 段世华, 等. 2013. 四种水稻品种的光合光响应曲线及其模型拟合[J]. 生态学杂志, 32(3): 604-610.
叶子飘, 于强. 2008. 光合作用光响应模型的比较[J]. 植物生态学报, 32(6): 1356-1361.
叶子飘. 2007. 光响应模型在超级杂交稻组合-II优明86中的应用[J]. 生态学杂志, 26(8): 1323-1326.
张方秋, 李小川, 潘文, 等. 2012. 广东生态景观树种栽培技术[M]. 北京: 中国林业出版社: 142-143.
张宏达. 1999. 中国植物志[M]. 北京: 科学出版社: 7.
Photosynthesis Light Response Characteristics of Camellia azalea and Fitting of Application Models
ZHANG Fangqiu, YANG Huixiao, XU Bin*, PAN Wen
Guangdong Provincial Key Laboratory of Bio-control for the Forest Disease and Pest, Guangzhou 510520, China; Guangdong Academy of Forestry, Guangzhou 510520,China
Camellia azalea is a Camellia species endemic to China, and only confined to the E,huangzhang Nature Reserve in Yangchun City of Guangdong Province. It is widely used for landscapes because of its perfect form and the characteristics to bloom year-round . The reports on fitting of photosynthesis light-response curve model of C.azalea were seldom. Six typical models of light-response curve of leaf net photosynthesis, such as Binomial regressions model, Non-rectangular hyperbola model, Rectangular hyperbola model, Modified rectangular hyperbola model, Exponential model and Modified exponential model, were used to fit the photosynthesis light response curves of C. azalea. The results showed that there was a nonlinear relationship between photosynthesis rate and light intensity. Modified rectangular hyperbola model provided a best fit for the photosynthetic light-response curve. Except the Binomial regressions model, the estimated data from the photosynthesis light-response curve of C. azalea by the other models all were close to the measured values, and only the Modified rectangular hyperbola model could fit the data under photoinhibition. However, only the photosynthetic parameters estimated by Modified rectangular hyperbola model were fitting well with the measured values, and the correlation coefficient (R2) is 0.998. The estimated Pmax, Isat, Ic, Rdand initial quantum efficiency (α) of C. azalea were 5.56 μmol·m-2·s-1, 1 327.30 μmol·m-2·s-1, 29.44 μmol·m-2·s-1, 1.909 μmol·m-2·s-1and 0.065 respectively. These data indicated that C. azalea is a sun plant, with strong light energy utilization ability, and has a certain shade tolerance and a wide range of adaptation to illumination.
Camellia azalea; light-response curve model of photosynthesis; photosynthesis rate; light intensity
10.16258/j.cnki.1674-5906.2015.10.002
Q948;X173
A
1674-5906(2015)10-1599-05
张方秋,杨会肖,徐斌,潘文. 杜鹃红山茶的光响应特性及其最适模型筛选[J]. 生态环境学报, 2015, 24(10): 1599-1603.
ZHANG Fangqiu, YANG Huixiao, XU Bin, PAN Wen. Photosynthesis Light Response Characteristics of Camellia azalea and Fitting of Application Models [J]. Ecology and Environmental Sciences, 2015, 24(10): 1599-1603.
国家“十二五”农村领域国家科技计划子研究项目资助项目(2012BAD01B0703-2)
张方秋(1964年生),男,研究员,硕士,主要研究方向为林木遗传育种。E-mail: fqzhang001@sinogaf.cn *通信作者:徐斌(1974年生),男,教授级高工,博士,主要研究方向为林木遗传育种。E-mail: xubin@sinogaf.cn
2015-08-04
猜你喜欢
文苑(2020年6期)2020-06-22
小学生学习指导(低年级)(2019年9期)2019-09-25
中学生数理化·中考版(2019年8期)2019-07-13
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
大观(书画家)(2018年6期)2018-07-08
中学生数理化·七年级数学人教版(2017年9期)2017-08-15
新课程·下旬(2017年7期)2017-08-14
民族音乐(2016年1期)2016-08-28
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
