
耐瘠杂交粳稻及高产粳稻品种的氮代谢相关指标研究
2015-12-05 02:59:48朱高倩张忠林金寿林字秋艳郑玉珍姜珍珍闫成强谭学林
作物研究 2015年4期
柳 展,朱高倩,张忠林,金寿林,字秋艳,郑玉珍,姜珍珍,闫成强,谭学林
(云南农业大学稻作研究所/云南省杂交粳稻工程技术研究中心,昆明650201)
氮素是水稻生长发育所需的主要元素,水稻靠其根系从土壤中吸收NO-3-N和NH+4-N[1]。根系吸收的-N通过硝酸还原酶(NR)和亚硝酸还原酶还原成NH+4以后被同化[2]。NH+4主要通过谷氨酰胺合成酶(GS)/谷氨酸合酶代谢途径,合成谷氨酸等有机氮[3],这一途径合成的有机氮占氮素总同化量的95%以上[4]。经氮代谢同化形成的有机氮供给植株地上部分生长发育。在低氮条件下,水稻的光合作用、氮代谢等在内的许多生理生化过程以及诸多农艺学性状都会受到负面影响,最终导致水稻的生物产量和经济产量减少[5]。在水稻大田生产中,稻农通常通过大量施用氮肥以获取水稻高产。在我国,用于水稻生产的氮肥施用量在过去几十年中迅速增加[6]。但是,由于所施用氮肥的30% ~70%都通过各种途径流失[7],导致氮肥的利用率平均只有35%左右[6]。氮肥利用率低增加了水稻的种植成本,导致稻农的经济收入下降。此外,大量的氮素流失也直接或间接地导致了环境的污染,影响了生态环境及农业的可持续发展[8]。
为促进不同稻区水稻产量的不断提高,针对云南省粳稻生产条件复杂多样的现状,云南的水稻育种工作者选育出了适宜云南不同气候和生产条件的品种,如高产优质滇型杂交粳稻滇杂31、滇杂35以及常规粳稻楚粳28等。虽然云南省粳稻种植面积占水稻总种植面积的60%,但与我国杂交粳稻总体发展缓慢相一致,云南杂交粳稻的种植面积也还不到粳稻面积的 10%[9,10],因此,研究低氮条件下杂交粳稻品种氮代谢的特点,对于选育氮肥高效利用的杂交粳稻品种,推广杂交粳稻种植,保证水稻高产稳产、降低水稻生产成本、进行环境友好水稻生产具有重要的意义。
本研究以云南生产上耐瘠性不同的5个粳稻品种为材料,研究在正常和低氮条件下供试材料的氮素代谢特点,分析7个氮素代谢指标和3个农艺指标,旨在为耐低氮水稻品种选育及水稻生产氮肥高效利用提供理论参考。
1 材料与方法
1.1 试验材料
供试材料共5个粳稻品种(系),分别是高产滇型杂交粳稻品种滇杂31、耐瘠滇型杂交粳稻品种滇杂35及其亲本保持系合系42-5B和恢复系南34、常规粳稻超级稻品种楚粳28。
1.2 试验设计
在云南农业大学稻作研究所网室内装置2个直径为15 cm,首尾相接呈“O”型的PVC管。在PVC管上依次开直径为10 cm的圆口,圆口间距为15 cm。PVC管中装7.5 cm高的培养液。试验设氮素正常含量组和低氮含量组(氮素含量为正常组氮素水平的1/4)2个处理。每个处理的每个品种栽5盆,重复3次,完全随机排列。氮素含量正常组的培养液参照国际水稻所的Yoshida培养液配方[11](含114.25 mg/L NH4NO3、50.38 mg/L NaH2PO4·2H2O、89.25 mg/L K2SO4、110.75 mg/L CaCl2、405 mg/L MgSO4·7H2O、1.86 mg/L MnCl2·4H2O、0.093 mg/L(NH4)6·MO7O24·4H2O、1.17 mg/L H3BO3、0.044 mg/L ZnSO4· 7H2O、0.039 mg/L CuSO4·5H2O、9.63 mg/L FeCl3·6H2O、14.86 mg/L柠檬酸,所用试剂均为分析纯)配制,另作适当修改,即9.63 mg/L FeCl3·6H2O中的 Fe3+与 1.50 mg/L EDTA-Na2形成螯合物Fe(Ⅲ)-EDTA配制,正常组NH4NO3为1.44 mmol/L的氮含量,相当于大田施用分析纯NH4NO385.7 kg/hm2(参照文献[12]的方法换算)。低氮含量组除NH4NO3含量为正常组的1/4(0.36 mmol/L)之外,其余营养液成分与氮素正常组相同。
供试材料在大田按普通生产方法育苗,5叶1心时挑取长势一致的健壮秧苗,用蒸馏水冲洗根部,然后移栽至底部开孔、装有沙石为固定基质的塑料栽培盆(上端直径11 cm、下端直径7 cm、高9 cm),每盆2穴,每穴1粒谷秧,栽相同品种。将栽培盆分别置于2个装有不同氮含量培养液的PVC管上。水稻栽培过程中,利用潜水泵使培养液处于循环流动状态。每天补充新鲜的去离子水以维持营养液的体积,每天早晚各1次用稀盐酸和氢氧化钠溶液调节培养液pH至5.0左右,每4 d更换1次培养液。每个处理的每个品种各取样5株。在主穗幼穗抽出叶鞘2 mm时,分别测定根系氧化力等7项生理生化指标值,成熟期取样测定有效穗数、株高以及生物产量。
1.3 测定项目与方法
1.3.1 根系氧化力和根系还原力的测定
随机取每棵样株的粗分枝根和细分枝根,用蒸馏水洗净并用滤纸吸干水分,然后称取0.50 g,采用α - 萘胺法[13,14]和 TTC(三苯基氯化四氮唑)还原法[13]分别测定根系氧化力和根系还原力。
1.3.2 叶片NR活性及GS活性的测定
随机取每棵样株2个分蘖茎的水稻倒二叶,用预冷的蒸馏水洗净并用滤纸吸干水分之后,去除中脉,迅速分别称取0.30 g及0.50 g,并采用陈薇等[2]和 Teng 等[15]的方法分别进行叶片 NR 和 GS活性的测定。酶的提取过程在4℃条件下进行。
1.3.3 叶片叶绿素含量的测定
随机取每棵样株2个分蘖茎的水稻倒二叶,用蒸馏水洗净并用滤纸吸干水分之后,去除中脉,称取0.50 g。参照Yoshida等[11]的方法测定叶片的叶绿素含量。
1.3.4 有效穗数、株高及单株生物产量的测定
在成熟期,测定每棵样株的有效穗数和株高,并收获水稻全株,用蒸馏水冲洗过后自然风干至恒重,用电子天平测定水稻的单株生物产量。
1.4 数据处理
采用Excel 2007以及SPSS统计软件对数据进行方差分析。
2 结果与分析
2.1 2种氮量条件下供试水稻品种的根系氧化力及还原力
与正常氮素组相比,5个供试水稻品种在低氮组的根系氧化力均出现不同程度的下降(表1),其中保持系合系42-5B、恢复系南34、高产品种楚粳28以及滇杂31的根系氧化力下降显著,而耐瘠品种滇杂35的根系氧化力虽有下降,但与在正常氮素条件下相比,差异并不显著。
与根系氧化力的变化趋势相反,在低氮条件下5个品种的根系还原力均比正常氮素条件下有所增大(表1),其中,滇杂35在2种生长条件下的差异达到显著水平,但其余4个水稻品种在2种生长条件下的差异不显著。
2.2 2种氮量条件下供试水稻品种的NR和GS活性
与正常氮素组相比,在低氮条件下,滇杂35的NR活性下降显著,而其余4个水稻品种均下降极显著。GS活性的变化有所不同:与正常氮素组相比,虽然不同水稻品种的GS活性均出现不同程度的下降,但南34及楚粳28的下降达极显著,合系42-5B及滇杂31的下降达到显著水平,而滇杂35的酶活性虽有下降,但差异不显著(表1)。
2.3 2种氮量条件下供试水稻品种的叶绿素含量
与正常供氮处理相比,在低氮条件下,滇杂35的叶绿素a、b含量及总含量有升有降,但差异不显著(表1)。与滇杂35不同的是,合系42-5B、南34以及滇杂31的叶绿素a含量显著升高,叶绿素b及叶绿素总含量却显著下降,且达到极显著差异水平。楚粳28的叶绿素a含量与滇杂35相似,在低氮条件下虽有上升,但差异不显著;叶绿素b及叶绿素总含量与合系42-5B的变化相似,下降达到极显著水平。
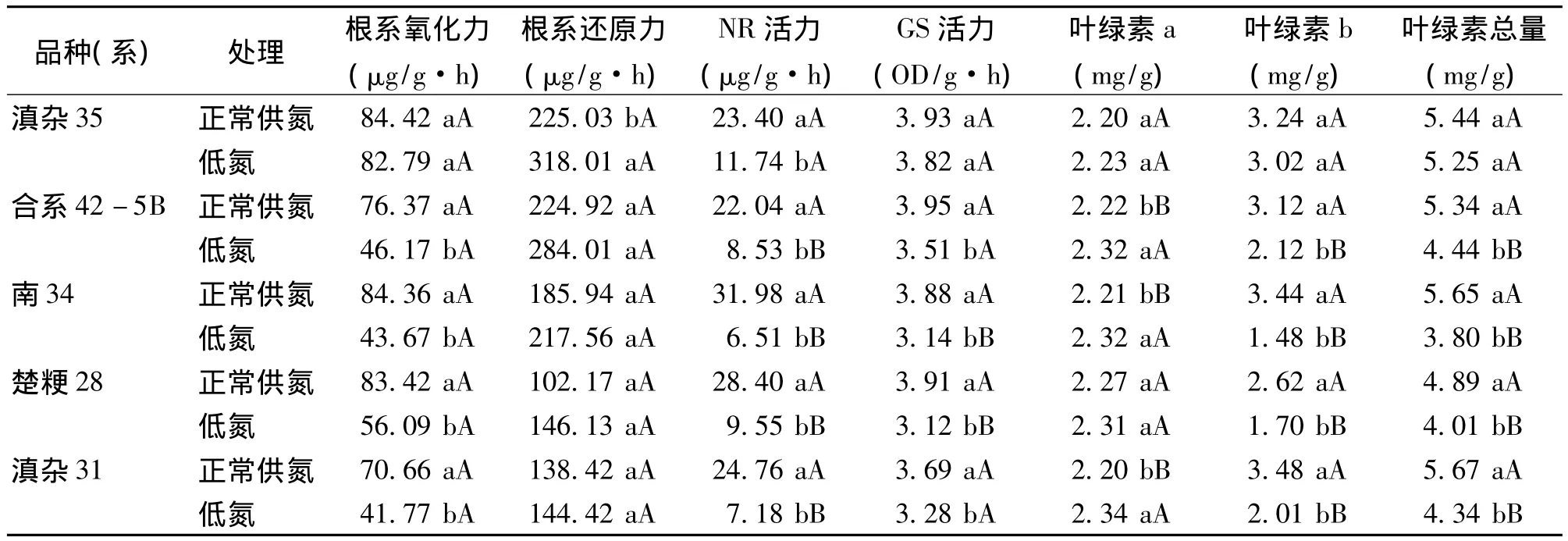
表1 2种氮量条件下供试水稻品种的7个氮代谢指标
2.4 2种氮量条件下供试水稻品种的有效穗数、株高及生物产量
与正常供氮处理相比,在低氮条件下,滇杂35的有效穗数略有减少,但差异不显著,而其他4个水稻品种的有效穗数减少明显,差异显著或极显著(表2)。
与正常供氮处理相比,在低氮条件下,5个供试品种的株高及生物产量均下降明显,差异极显著(表2)。

表2 2种氮量条件下供试水稻品种的3个农艺性状指标
3 结论与讨论
耐低氮水稻品种对低氮胁迫的响应通常是通过较强的氮素吸收或者氮素同化利用能力来获得足够的养分[16]。根系是水稻从土壤中吸收氮素的主要器官,而硝酸还原酶等氮代谢酶又是将根系吸收的无机氮迅速同化为有机氮的关键物质[17]。在低氮条件下,耐低氮水稻品种均具有较大的根长、根体积和根密度以及较强的根系活力,同时氮代谢相关酶活性均比非耐低氮品种的要高[3,4,17,18],在耐瘠品种选育中,这些均可作为耐低氮水稻品种筛选的指标[16,17,22]。氮素是叶绿素 a 和叶绿素 b 的重要组成成分。在水稻叶片中约75%的氮与叶绿体有关[19]。水稻的株高和有效穗数[20]及生物产量[21]与水稻的氮素利用以及最终的籽粒产量均有密切的关系。本研究中在低氮胁迫条件下,耐瘠水稻品种滇杂35具有根系还原力显著上升,而根系氧化力、谷氨酰胺合成酶(GS)活性减少不明显,叶绿素b、叶绿素总含量和有效穗数无显著变化的特性,这与魏海燕等[22]以及程建峰等[17]的研究结果相一致。综合来看,耐瘠水稻品种的耐低氮特性可能是根系形态与生理特性以及氮素同化、地上部光合作用等共同作用的结果。本试验结果与通过高、中、低3种施氮水平对滇杂35进行的耐瘠性田间试验结果一致[23],因此,根系氧化力、根系还原力、GS 活力、叶绿素b、叶绿素总含量和有效穗数可以作为筛选耐低氮水稻品系和种质的参考指标。值得注意的是,从7个氮代谢相关指标和3个农艺性状来看,滇杂35对低氮胁迫的响应都与其2个亲本保持系合系42-5B和恢复系南34明显不同,这说明氮代谢和农艺性状的相关指标可以作为解析水稻耐低氮胁迫的杂种优势的分子遗传及生理生化的基础。
[1]张辰明,徐烨红,赵海娟,等.不同氮形态对水稻苗期氮素吸收和根系生长的影响[J].南京农业大学学报,2011,34(3):72 -76.
[2]陈 薇,张德颐.植物组织中硝酸还原酶的提取、测定和纯化[J].植物生理学通讯,1980(4):45-49.
[3]Campbell WH.Molecular control of nitrate reductase and other enzymes involved in nitrate assimilation[A].In:Foyer CH,Noctor G.Photosynthetic Nitrogen Assimilation and Associated Carbon and Respiratory Metabolism[C].The Netherlands:Kluwer Academic,2002.35 -48.
[4]Barneix AJ.Physiology and biochemistry of source-regulated protein accumulation in the wheat grain[J].Journal of Plant Physiology,2007,164:581 -590.
[5]Novoa R,Loomis RS.Nitrogen and plant production[J].Plant and Soil,1981,58:177 - 204.
[6]杨林章,孙 波,刘 健,等.农田生态系统养分迁移转化与优化管理研究[J].地球科学进展,2002,17(3):441-445.
[7]Zhu ZL,Chen DL.Nitrogen fertilizer use in China- con-tributions to food production,impacts on the environment and best management strategies[J].Nutrient Cycling in Agroecosystems,2002,63:117 -127.
[8]Erisman JW,Vries W.Nitrogen deposition and effects in European forests[J].Environ Rev,2000,8(2):65 -93.
[9]赵江海,邓华凤,陈光辉.杂交粳稻生产中存在的问题与对策[J].作物研究,2007,21(5):612-615.
[10]李全衡.加速发展云南杂交粳稻之我见[J].种子科技,2011,29(4):15 -16.
[11]Yoshida S,Forno DA,Cook JH,et al.Laboratory Manual for Physiological Studies of Rice:3rd Edn[M].Manila:International Rice Research Institute,1976:60 -65.
[12]戢 林,李廷轩,张锡洲,等.氮高效利用基因型水稻根系形态和活力特征[J].中国农业科学,2012,45(23):4770-4781.
[13]Gunter Trolldenier.Visualization of oxidizing power of rice roots and of possible participation of bacteria in iron deposition [J].Z Pflanzenerndhr Bodenk,1988,151:117-121.
[14]张志良,翟伟菁,李小方.植物生理学实验指导[M].北京:高等教育出版社,2009.30-33.
[15]Teng YB,Li YJ.Characterization of nitrogen metabolism in the low-nitrogen tolerant int1 mutant of Arabidopsis thaliana under nitrogen stress[J].Pedosphere,2010,20(5):623-632.
[16]杨肖娥,孙 羲.不同水稻品种对低氮反应的差异及其机制的研究[J].土壤通报,1992,29(1):73-79.
[17]程建峰,戴廷波,荆 奇,等.不同水稻基因型的根系形态生理特性与高效氮素吸收[J].土壤学报,2007,44(2):266-272.
[18]Miflin BJ,Habash DZ.The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops[J].Journal of Experimental Botany,2002,53:979-987.
[19]Dalling MJ.The physiological basis of nitrogen redistribution during filling in cereals[A].In:Harper JE.Exploitation of Physiological and Genetic Variability to Enhance Crop Productivity[C].Rockville:Am Soc of Plant Physiologists,1985.55 -71.
[20]郑家奎,文春阳,张 涛,等.耐低氮水稻材料筛选及筛选指标研究[J].安徽农业科学,2009,37(16):7361-7363,7377.
[21]杨惠杰,李义珍,杨仁崔,等.超高产水稻的干物质生产特性研究[J].中国水稻科学,2001,15(4):265-270.
[22]魏海燕,张洪程,张胜飞,等.不同氮利用效率水稻基因型的根系形态与生理指标的研究[J].作物学报,2008,34(3):429 -436.
[23]金寿林,谭学林,张忠林,等.滇型杂交粳稻新组合滇杂35[J]. 杂交水稻,2008,23(1):87-88.
猜你喜欢
安徽农学通报(2024年12期)2024-07-13 20:26:34
今日农业(2021年15期)2021-10-14 08:20:44
农民致富之友(2020年32期)2020-12-03 13:49:21
江苏农业科学(2020年22期)2020-03-03 14:37:13
现代农业科技(2018年14期)2018-10-31 10:50:20
河南农业(2017年7期)2017-07-25 09:22:05
安徽农业科学(2017年28期)2017-05-30 14:55:41
浙江农业科学(2016年11期)2016-05-04 04:16:39
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
