
冬虫夏草菌在寄主钩蝠蛾幼虫中的潜伏侵染过程研究
2015-12-03 05:55彭青云张古忍
环境昆虫学报 2015年2期
雷 桅,彭青云,张古忍,刘 昕
(中山大学生命科学学院,广州 510275)
虫生真菌侵染昆虫的途径通常有两种,其中少量的是通过口腔进入寄主,而余下大多数则是通过昆虫表皮侵入体内。口腔侵染研究较少,一般认为病原微生物主要从中肠侵入血腔。表皮侵染机制存在于大量真菌中,特别是生物防治领域,其中白僵菌、绿僵菌等研究较为深入,其过程可分为10个阶段:分生孢子附着、分生孢子在昆虫表皮上萌发、穿透表皮、菌丝在血腔内生长、毒素的产生、寄主死亡、菌丝侵入寄主的所有器官、菌丝穿出表皮、产生侵染单位、侵染单位扩散(蒲蜇龙和李增智,1996)。然而,由于种种原因,冬虫夏草菌的侵染机理至今仍几乎一无所知,因此有人推测该菌应该与其他虫生真菌类似,是通过表皮侵染进入钩蝠蛾幼虫(陆龙喜等,2002),而也有文献推论冬虫夏草菌可能经口腔进食途径侵入寄主(Zhang et al.,2008),但迄今仍无确切证据支持哪种途径才是冬虫夏草菌的侵染方式。
冬虫夏草是青藏高原特色名贵中药材,,具有极高的药用和经济价值,自然资源十分稀少且至今仍无法人工培植(Yue et al.,2013)。它是冬虫夏草菌(冬虫夏草无性型:中国被毛孢Hirsutella sinensis)寄生于鳞翅目 Lepidoptera 蝠蛾科Hepialidae 钩蝠蛾属Thitarodes 幼虫后形成的虫生子囊真菌,其发生机理一直是学术界非常关注却难以破解的课题,并极大地阻碍了冬虫夏草可持续利用关键技术的发展,也挑战着虫生真菌领域的广大科研工作者。其中首要问题就是冬虫夏草菌的侵染途径,即它是如何侵入寄主幼虫体内的(Zhong et al.,2010;Hu et al.,2013),从而进一步在某种条件下,子座开始形成并继续长出地面,由此启动冬虫夏草的有性阶段。依据目前已知的其他虫生真菌的侵染方式,推测冬虫夏草菌可能存在的潜伏侵染途径,一种可以称之为内向型,即环境中的冬虫夏草菌以幼虫表皮为突破口,通过附生在表皮上,然后基于一系列物理刺入或酶学溶解等过程进入体腔,从组织顺序的角度而言,就是表皮>脂肪体>血淋巴>肠壁;另一种则可称为外向型,即环境中的冬虫夏草菌首先定殖于植物根系,然后随着幼虫摄食过程从口腔进入肠道,并主动穿透或依赖于消化系统侵染虫体,直至进入体腔,实现对寄主的感染,其组织过程可概括为:肠壁>血淋巴>脂肪体>表皮(蒲蜇龙和李增智,1996)。本研究应用Holt 指数平滑法模型评估冬虫夏草菌在幼虫各组织中水平迁移的线性趋势,并依据上述两种潜在侵染模式,分别进行建模评估,比较分析两者存在的可能性和可行性。
1 材料与方法
1.1 样品采集与生境调查
西藏东南部的色季拉山海拔4000-4600 m 间是良好的冬虫夏草发生地,在4156 m 海拔处建立有“中山大学青藏高原特色资源科学工作站”被用于冬虫夏草原生态生物学的科学研究。从海拔3800 m 起往上行,每隔200 m 设一个采样点,每个点采集5 条寄主蒲氏钩蝠蛾幼虫,龄期为3-6 龄,带回备用。
1.2 DNA 的制备
使用DNA 抽提试剂盒提取钩蝠蛾幼虫表皮、脂肪体、血淋巴和肠壁等组织中的基因组DNA,然后结合NanoDrop 2000 超微量分光光度计和琼脂糖凝胶电泳方法检测其浓度、质量和完整性,最终总DNA 被储存于-80℃超低温冰箱中。
所有DNA 提取方法都按照AxyPrep 基因组DNA 小量试剂盒(Axygen 生物技术有限公司)说明书进行。
1.3 qPCR 扩增
基于Real-time qPCR 方法建立了高灵敏度和高特异性的冬虫夏草菌实时定量检测技术体系(R2>0.999),包括物种特异性引物IF2(5'-GCAGTGGCATCTCTCAGTCA-3')和IR2(5'-GCATTTCGCTGCGTTCTT-3'),并以提取的各幼虫表皮、脂肪体、血淋巴和肠壁等组织DNA 为模板,检测其中的ITS 拷贝数,后换算成冬虫夏草菌定殖量。反应体系为:样本基因组DNA 5.0 μL,SYBR Green PCR Master Mix 10 μL,上下游引物各0.5 μL,最后以ddH2O 补至20 μL。每个重复测定3 次(Lei et al.,2013)。
1.4 数据分析与数学建模
为了深入探讨冬虫夏草菌侵染寄主幼虫的途径问题,即表皮侵染途径(外向型outward)或者口腔侵染途径(内向型inward)。需要特别说明的是,表皮、脂肪体、血淋巴和肠道是相互联系但各自独立的组织,冬虫夏草菌在其上的定殖和迭续将遵守方向性的线性关联。本研究采用了Holt线性趋势的指数平滑法模型,该模型适用于具有线性趋势且无季节性的序列,其平滑参数是水平和趋势,不受相互之间的值约束。Holt 模型比Brown 模型更通用,但在计算大序列时要花的时间更长。Holt 指数平滑法与ARIMA 模型极为相似,包含零阶自回归、二阶差分以及二阶移动平均数(Gardner,1958;Gardner,2006)。
2 结果与分析
2.1 不同纬度产区中钩蝠蛾幼虫体内冬虫夏草菌定殖量的比较分析
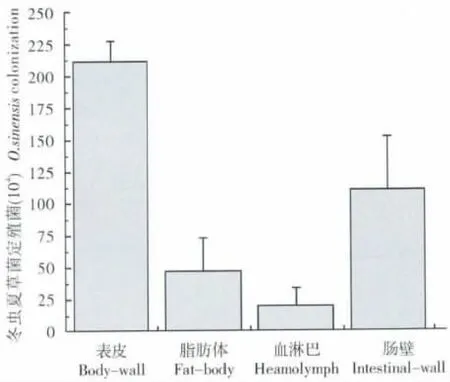
图1 不同纬度产区寄主钩蝠蛾幼虫体内的冬虫夏草菌定殖量对比Fig.1 Colonization of Ophiocordyceps sinensis in the tissues of Thitarodes larvae

图2 评估模型的拟合度Fig.2 Evaluation of the degree of fitting of the model
如图1 可知,在寄主钩蝠蛾体内血淋巴中的冬虫夏草菌含量最低。昆虫的血淋巴,兼有血液和淋巴样组织液的特性,其作用是营养物质运输、体腔清洁、温度调节、防御免疫和创伤愈合等,因此对于生理状态完全正常的幼虫而言,血淋巴中的菌含量不可能很高。
同时,我们也发现肠壁定殖有大量的冬虫夏草菌,这来源于进食过程中随根系和土壤到达肠道,而消化系统也能为菌体的生长提供充足的营养和能量物质,所以肠壁就构成了钩蝠蛾幼虫体内的一个菌库(Lei et al.,2011)。
蒲氏钩蝠蛾幼虫表皮携带有相对最多冬虫夏草菌,同时我们在该地土壤中检测到大量目标菌,而且植物根系也被证实含有高丰度的冬虫夏草菌(Peng et al.,2013),这说明钩蝠蛾表皮的带菌状态与周边土壤和植被环境密切相关。
2.2 冬虫夏草菌在寄主幼虫中潜伏侵染的趋向推测
(1)起始于表皮的潜伏侵染途径(内向型,inward)模式

图3 残差自相关函数图Fig.3 Residuals autocorrelation plot

表1 指数平滑法模型参数Table 1 Exponential Smoothing Model parameters

图4 模型趋势图Fig.4 Holt model trend
(2)起始于肠道的潜伏侵染途径(外向型,outward)模式

图5 评估模型的拟合度Fig.5 Evaluation of the degree of fitting of the model

图6 残差自相关函数图Fig.6 Residuals autocorrelation plot

表2 指数平滑法模型参数Table 2 Exponential Smoothing Model parameters
对比上述两个模型,反映了方向相反的两个过程,其中起始于表皮的内向型模型为0.678,起始于肠道的外向型模型为0.271。这两个R2值都不高,说明冬虫夏草菌在幼虫体内的增殖迁移过程并不简单,不是单一地依赖某一种途径,而是还受到其他多方面的影响。相对而言,内向型模式的R2值比外向型模式高出近3 倍,说明这两种途径均有可能存在,但后者发生的过程会伴随更多的影响因素。

图7 模型趋势图Fig.7 Holt model trend
在相关工作中,我们已经证实在冬虫夏草发生地的土壤和植物等环境中都生活着大量冬虫夏草菌,因此幼虫表皮接触和感染该菌的机会较大,但表皮的防护结构和功能较强,对菌群的侵入会造成一定的限制;而肠道里的定殖菌只能通过摄食带菌植物根系的方式进入,途径和来源比较有限,而且还与植物本身的带菌丰度、肠道里的pH值、消化系统里的多种溶菌抑菌活性物质等有关。总之,基于本研究的模拟研究结果,虽然依据真菌侵染昆虫的传统观念,冬虫夏草菌的表皮接触侵染方式具有可行性;但起始于肠道定殖菌群,并自肠壁侵入和感染体腔的模式也具有一定的潜在可能性,应在冬虫夏草发生机理的研究中被重点关注。
3 结论与讨论
内生冬虫夏草菌的侵染途径和寄生进程被认为是冬虫夏草发生发育的关键影响因素,但由于实验手段的缺乏和培养条件的严苛,这一过程尚无法实现人工复制(Zhang et al.,2008;Shrestha et al.,2011),给冬虫夏草的发育机理研究和产业化培植带来巨大困难,而当前最大挑战是揭示冬虫夏草菌如何侵入宿主并造成感染的病理性进程(Li et al.,2000;Dong et al.,2010)。
冬虫夏草作为一种虫生真菌,与已被深入研究的金龟子绿僵菌Metarhizium anisopliae 和球孢白僵菌Beauveria bassiana 等类似,也被认为具有典型的双型现象。冬虫夏草菌对寄主钩蝠蛾幼虫的带菌生长、侵染、寄生和致病等重要环节都依赖于该菌的定殖和增殖,所以要解析冬虫夏草发生的过程必须要量化冬虫夏草菌种群在寄主和生态环境中的变化。由于冬虫夏草菌形态不定,生长速度极其缓慢,培养条件又非常苛刻,而且至今仍无法从钩蝠蛾幼虫、植物和土壤等环境中直接分离获得,因此无法像其它病原菌那样通过分离和纯培养来进行定量分析,故基于现代分子生物学手段,建立和开发冬虫夏草菌实时定量检测技术将非常必要。
本文采用real-time qPCR 技术检测了冬虫夏草适生地蒲氏钩蝠蛾幼虫体内冬虫夏草菌的定殖量,探讨冬虫夏草菌群的寄生量及其种群变化趋势的时空模式,从而证实了幼虫带菌生长理论(齐丽丽,2010)。在此基础上,分别建立了起始于表皮的潜伏侵染途径和起始于肠道的潜伏侵染途径两种数学模型。经过比较分析后,推论冬虫夏草菌随口腔摄食侵染寄主幼虫的方式具有一定合理性和可行性,表明冬虫夏草菌可以通过进食途径实现其对钩蝠蛾幼虫的带菌生长、侵染和寄生,从而进一步支持了高寒宿主植物在冬虫夏草发生过程中的作用(雷桅,2012),完善了冬虫夏草菌与寄主昆虫之间的关系及冬虫夏草的生活史,为进一步揭示冬虫夏草的发生机理提供了重要依据。
References)
Dong CH,Yao YJ.On the reliability of fungal materials used in studies on Ophiocordyceps sinensis[J].Journal of Industrial Microbiology&Biotechnology,2010,38:1027-1035
Gardner ES Jr.Exponential smoothing:The state of the art[J].Journal of Forecasting,1958,4(4):1-28.
Gardner ES Jr.Exponential smoothing:The state of the art—Part II[J].International Journal of Forecasting,2006,22(4):637.
Hu X,Zhang YJ,Xiao GH,et al.2013.Genome survey uncovers the secrets of sex and lifestyle in caterpillar fungus[J].Chinese Science Bulletin,58(23):2846-2854.
Lei W.Interdependent Relationship between Host Insects,Plants and Hirsutella sinensis in Alpine Habitat[D].Guangzhou:Sun Yat-sen University,2012.[雷桅.高寒生境中国被毛孢与宿主植物及寄主昆虫依存关系研究[D].广州:中山大学,2012]
Lei W,Chen H,Zhang GR,et al.Molecular identification and food source inference of constructive plants,native to the Ophiocordyceps sinensis habitat[J].African Journal of Biotechnology,2011,10(2):159-167.
Lei W,Li SS,Peng QY,et al.A real-time qPCR assay to quantify Ophiocordyceps sinensis biomass in Thitarodes larvae[J].Journal of Microbiology,2013,51(2):229-233.
Li ZZ,Huang B,Fan MZ.Molecular evidence for anamorph determination of Cordyceps sinensis(Berk.)Sacc[J].Mycosystema,2000,19:60-64.
Lu LX,Liu SX,Shi LG.Advance on Ophiocordyceps sinensis studies[J].Bulletin of Biology,2002,37(6):4-6.[陆龙喜,刘淑梅,时连根.冬虫夏草研究的新进展[J].生物学通报,2002,37(6):4-6.
Peng QY,Zhong X,Lei W,et al.Detection of Ophiocordyceps sinensis in soil by quantitative real-time PCR[J].Canadian Journal of Microbiology,2013,59(3):204-209.
Pu ZL,Li ZZ.Insect Mycology[M].Hefei:Anhui Science &Technology Publishing House,1996.[蒲蜇龙,李增智.昆虫真菌学[M].合肥:安徽科学技术出版社,1996]
Qi LL.Study on Parasitism of Developmental Stages of Hepialus pui by Hirsutella sinensis[D].Guangzhou:Sun Yat-sen University,2010.[齐丽丽.蒲氏蝠蛾不同发育阶段被中国被毛孢寄生的动态研究[D].广州:中山大学,2010]
Shrestha G,Zhang WM,Zhang YJ,et al.What is the Chinesecaterpillar fungus Ophiocordyceps sinensis(Ophiocordycipitaceae)?[J].Mycology,2011,1:228-236.
Yue K,Ye M,Lin X,et al.The artificial cultivation of medicinal caterpillar fungus,Ophiocordyceps sinensis(Ascomycetes):A review[J].International Journal of Medicinal Mushrooms,2013,15(5):425-434.
Zhang YJ,Liu XZ,Wang M.Cloning,expression,and characterization of two novel cuticle-degrading serine proteases from the entomopathogenic fungus Cordyceps sinensis[J].Research in Microbiology,2008,159(6):462-469.
Zhong X,Peng QY,Qi LL,et al.rDNA-targeted PCR primers and FISH probe in the detection of Ophiocordyceps sinensis hyphae and conidia[J].Journal of Microbiological Methods,2010,83(2):188-193.
猜你喜欢
罕少疾病杂志(2021年4期)2021-07-28
世界最新医学信息文摘(2021年12期)2021-06-09
发明与创新·小学生(2021年12期)2021-01-06
军事文摘(2020年22期)2021-01-04
中国医学影像技术(2020年3期)2020-04-02
影像研究与医学应用(2019年1期)2019-01-21
汉语世界(2016年3期)2016-11-16
胃肠病学和肝病学杂志(2016年2期)2016-06-01
听力学及言语疾病杂志(2015年5期)2015-12-24
哈尔滨医药(2015年3期)2015-12-01
