
菜蚜茧蜂载体植物系统的构建及其控害效果
2015-12-03 05:55沈嘉炜蔡尤俊张文庆
环境昆虫学报 2015年2期
沈嘉炜,蔡尤俊,张文庆
(中山大学有害生物控制与资源利用国家重点实验室,广州 510275)
载体植物系统(banker plant system)是近年来开发的一种能自我维持并可有效降低目标害虫种群的新型生物防治技术。这一技术集保护与利用天敌、人工繁殖与释放天敌以及异地引进天敌等比较传统的生物防治技术特点于一身,强调生物防治的可持续性以及有效性,能极大改善传统生物防治技术的不足(Nagasaka et al.,2010;Ohta et al.,2010)。载体植物系统主要包括载体植物(banker plant)、替代寄主或替代食物(alternative hosts or food)以及有益生物(beneficial)三个基本要素(Frank,2010;肖英方等,2012)。其中,载体植物是系统中最重要的要素,其最主要作用是作为系统载体,饲养替代寄主或替代食物,或者直接将植物本身作为营养来源应用于有益生物的养殖(Xiao et al.,2011);另一个基本要素替代寄主或食物的主要作用则是提供食物资源给有益生物(通常为寄生性天敌或捕食性天敌),让有益生物能通过这一系统大量地自主繁殖(Wong et al.,2012);最后一个基本要素有益生物则主要通过利用载体植物系统中的替代寄主或食物大量自主繁殖后,自动扩散到目标作物上,对靶标害虫进行有效控制。目前,该系统已经在19种天敌中成功构建,用于11种常见害虫的防治(Frank,2010)。据统计,目前美国约有1%到5%的温室使用载体植物系统,加拿大约有10%-25%种植者使用这一系统。而在欧洲,丹麦使用率达20%,荷兰使用这一系统防治蚜虫的面积约为120 hm2(Sarah et al.,2012)。我国载体植物系统的研究、应用则相对较少,因此开发载体植物系统具有相当广阔的前景。
目前,全世界的蚜虫已鉴定的达4700 多种。蚜虫具有种类多、寄主杂,分布广、数量大、繁殖快、孤雌生殖、世代重叠、危害巨大等特点,每年对我国作物造成的损失很大。菜蚜Lipaphis erysimi 又名萝卜蚜,主要成群集结在蔬菜叶片背面,吸食叶片汁液,危害蔬菜生长(于东坡等,2009)。如何对蚜虫进行有效的防治,一直以来都是困扰广大农林工作者的问题(唐平华,2012)。Mulder 等(1999)利用谷类植物为载体植物,以粗脊蚜茧蜂Aphidius colemani Viereck 为有益生物、禾谷缢管蚜Rhopolosiphum padi L.为替代寄主,构建的载体植物系统成功用于棉蚜Aphis gossypii Glover 防治。在我国,利用烟蚜茧蜂Aphidius gifuensis Ashmaed 防治桃蚜Myzus persicae Sulzer(又称烟蚜)的技术已比较成熟。李明福等(2006)在云南玉溪以1∶50 的雌蜂、桃蚜比在烟田释放烟蚜茧蜂,桃蚜寄生率达到85%-92%,僵蚜羽化率在85%-90%,防治效果达到61.2%-89.3%;龙宪军等(2012)在湖南湘西以1∶100的雌蜂、桃蚜比在烟田释放烟蚜茧蜂,35 d 后防平均治效率达到83.0%。迄今,利用载体植物系统防治菜蚜尚未见报道。
菜蚜茧蜂Diaeretiella rapae M'Intosh 属于膜翅目蚜茧蜂科,是专性寄生蚜虫的一种内寄生蜂。菜蚜茧蜂对蚜虫防治具有非常大的潜力,在我国各地均有分布,其中在广州、江西以及西宁等菜地中属于优势种。据刘宏谋(1980)报道,菜蚜茧蜂对油菜蚜的防治率可达48%。
本文拟构建菜蚜茧蜂载体植物系统,应用于蔬菜蚜虫的生物防治。
1 材料与方法
1.1 实验植株
实验用烟草Nicotiana tabacum L.种子由华南农业大学提供,菜薹Brassica campestris L.种子在青县王镇店种子繁育站购买,蚕豆Vicia faba L.种子在广州市大学城贝岗村菜市场购买。所有植物在24℃±2℃温室种植。
1.2 实验蚜茧蜂及蚜虫
实验用菜蚜茧蜂主要通过在广州市白云区人和镇菜地采集僵蚜获得。待成蜂羽化后用10%蜜糖水进行喂养,确认为菜蚜茧蜂后放入网笼中,提供长有菜蚜的菜薹植株进行扩繁。僵蚜出现后,逐日收集僵蚜,装入10 mL 离心管中备用。新羽化的第二代菜蚜茧蜂按雌雄比1∶1 进行配对喂养,雌蜂用于实验备用。
桃蚜由贵州大学提供,挑选若干无翅、无病、未被寄生的成蚜接种到长至5 叶的烟草上进行扩繁备用,用塑料罩以及网笼笼罩,防治蚜虫逃逸及被寄生。
菜蚜在广州市白云区人和镇田间进行采集,采集后将其接种到菜薹植株上进行扩繁备用,用塑料罩以及网笼笼罩,防止蚜虫逃逸及被寄生。
蚕豆蚜Aphis craccivora 由华南农业大学提供,接种到适龄蚕豆苗植株上进行扩繁备用,用塑料罩以及网笼笼罩。
甘蓝蚜Brevicoryne brassicae(Linnaeus)在广州市花都区田间采集,采集后接种到适龄甘蓝植株上进行扩繁备用,用塑料罩以及网笼笼罩。
1.3 替代寄主筛选
根据菜蚜茧蜂的主要寄主范围以及广东地区危害较大、较常见的田间蚜虫种类,挑选出桃蚜、蚕豆蚜、以及甘蓝蚜三种蚜虫作为候选替代寄主,用于载体植物系统的构建。
1.3.1 菜蚜茧蜂对替代寄主的寄生率与羽化率
在培养盆(直径10 cm、深15 cm,下文中所有烟草、菜薹植株均用此规格的培养盆种植)中种植龄烟草、甘蓝Brassica oleracea L.、蚕豆Vicia faba L.等三种植株,当植株长到适龄的时候,每株分别接入100 头3-4 龄无翅、无病、未被寄生的桃蚜、甘蓝蚜或蚕豆蚜,用无底圆柱型塑料罩将各植株罩住,纱网封顶。蚜虫适应1 d,并确认数目无误后分别接入1 头已交配的雌性菜蚜茧蜂,任其自由寄生24 h,24 h 后将雌蜂移出。雌蜂移出后逐日观察各植株,将出现的僵蚜及时收集到10 mL 离心管,直至植株不再出现僵蚜。记录僵蚜数量以及羽化情况。实验重复3 次。根据菜蚜茧蜂对候选替代寄主的寄生率和羽化率,筛选出寄生率高、羽化率高的寄主作为菜蚜茧蜂载体植物系统的候选替代寄主。
1.3.2 替代寄主对目标作物的潜在性危害
一般情况下,载体植物系统对于其自身替代寄主的要求除了必须在有益生物的取食、寄生偏爱范围内之外,对其取食通常也要求是高度专一的,最起码不能对目标作物存在潜在的大危害,否则其迁移到目标作物上只会适得其反。因此,在筛选替代寄主时也必须对替代寄主会否对目标作物存在潜在性危害做验证。
在温室中24℃±2℃准备用培养盆种植的适龄烟草植株,在其上接入各龄桃蚜1000 头左右,任其自由繁殖,以烟草植株为圆心,在其半径0.5 m、1 m、3 m、5 m 的圆周处各放置4 盆菜薹植株,确保植株上没被感染蚜虫,逐日观察各菜薹植株上是否有桃蚜出现,统计其数量。根据各半径圆周上菜薹植株出现的桃蚜数量以及龄期,初步判定桃蚜是否对菜薹植株造成危害,以及其危害扩散范围为多少。实验重复3 次。
1.4 载体植物筛选
载体植物的筛选主要根据替代寄主的取食偏爱性以及对植株的适应性进行选择。
根据替代寄主的筛选结果,以替代寄主的取食范围以及广东地区常见的蔬菜作为候选载体植物。
候选载体植物:烟草、玉米Zea mays、生菜Lactuca sativa、甘蓝B.oleracea L.、蚕豆V.faba L.、菜薹B.campestris L.。
替代寄主:桃蚜。
1.4.1 替代寄主取食偏爱性(选择性试验)
截取6种候选载体植物的嫩叶,并将每种植株嫩叶剪成半径为2 cm 的圆形,并将其以圆形等距分布铺在有湿润滤纸和棉花的培养皿(直径20 cm)中,在培养皿中心放置50 头3-4 龄桃蚜,用纱网笼罩圆盆,防止蚜虫逃逸。分别于24 h以及48 h 后统计各叶片上桃蚜数量以及死亡虫数。实验重复9 次。
1.4.2 替代寄主适应性(非选择性试验)
分别在烟草、玉米、生菜、甘蓝、蚕豆、菜薹等6种候选载体植物植株上接入替代寄主(桃蚜)50 头,并用纱网将其笼罩,防止其逃逸。每24 h 统计6种植物上的蚜虫数量。实验重复3 次。
1.5 菜蚜茧蜂载体植物系统蜂(♀)蚜比确定
在5 棵高约50 cm 的烟草植株上分别接入3-4 龄无翅、无病且未被寄生的桃蚜25、50、100、150 以及200 头。在其适应一天,并确认数目无误后分别接入一头已交配的雌性菜蚜茧蜂,形成1:25、1∶50、1∶100、1∶150 以及1∶200 五个蜂(♀)蚜比梯度,任其自由寄生24 h,24 h 后将雌蜂移出。雌蜂移出后逐日观察各植株,将出现的僵蚜及时收集到10 mL 离心管,直至植株不再出现僵蚜。记录僵蚜数量以及羽化情况。实验重复3 次。
根据菜蚜茧蜂对桃蚜的寄生率、羽化率以及出蜂量等综合考虑,遴选出寄生率高、羽化率高、出蜂量大的蜂蚜比作为载体植物系统的实施蜂蚜比。
1.6 菜蚜茧蜂载体植物系统构建
在3 株高约30 cm、有5、6 片嫩叶的烟草植株上接入4-5 龄无翅、无病且未被寄生的桃蚜100 头,分别置于3个50 cm×50 cm×50 cm 的网笼中,让桃蚜自由繁殖。当3 株烟草上的起始桃蚜量分别达到1500 头、2000 头和2500 头时,按1∶100 的蜂(♀)蚜比,分别接入15、20 和25 头已交配的雌性菜蚜茧蜂。每天浇水以保证烟草植株持续存活,7 d 后开始记录各烟草植株上的僵蚜数量,每7 d 记录1 次,直到桃蚜被全部寄生。此实验重复2 次,用于确定最佳桃蚜起始数量并构建菜蚜茧蜂载体植物系统。
1.7 菜蚜茧蜂载体植物系统对目标害虫的控制作用
1.7.1 系统繁殖的菜蚜茧蜂对菜蚜的寄生率及僵蚜羽化率
在株高约10 cm 菜薹植株上接入3-4 龄无翅、无病、未被寄生的菜蚜100 头。在其适应1 d,并确认数目无误后接入1 头由载体植物系统繁殖所得的已交配的雌性菜蚜茧蜂,任其自由寄生24 h,24 h 后将雌蜂移出。雌蜂移出后逐日观察各植株,将出现的僵蚜及时收集到10 mL 离心管,直至植株不再出现僵蚜。记录僵蚜数量以及羽化情况。实验重复6 次。
1.7.2 菜蚜茧蜂载体植物系统对温室菜蚜的防治效果
在高约30 cm 的烟草植株(小钵直径10 cm)上接入4-5 龄无翅桃蚜100 头,让桃蚜自由繁殖(10 d 左右),当桃蚜达到2000 头左右时,接入20 头已交配的雌性菜蚜茧蜂,构建菜蚜茧蜂载体植物系统。选取接蜂7 d 后的菜蚜茧蜂载体植物系统备用。在温室中24℃±2℃选取株高约20 cm 的菜薹植株30 盆,分别置于两个1 m×1 m×1 m 的白色网笼中,每个网笼15 盆。分别向每株菜薹上接人菜蚜20 头,自由繁殖15 d 后,统计各菜薹上菜蚜的数量,并在处理组网笼中央置入1 株菜蚜茧蜂载体植物系统,对照组网笼中央置入1 株大小相似的纯净烟草植株。之后每7 d 统计各菜薹植株上的菜蚜数量,分析载体植物系统防控菜蚜的效果。实验重复15 次。
1.8 数据统计分析
数据为平均数±标准误,采用t-test、单因素方差分析(one-way ANOVA)等方法检验平均数之间的差异显著性,所有步骤均在软件EXCEL 2007 中进行。
2 结果与分析
2.1 替代寄主筛选
2.1.1 菜蚜茧蜂对替代寄主的寄生率与羽化率
在3种候选替代寄主中,菜蚜茧蜂对桃蚜和甘蓝蚜的寄生率分别为69.00%和60.33%,两者均显著高于蚕豆蚜的26.33%(F3,8=31.68,P<0.01);桃蚜和甘蓝蚜僵蚜的羽化率分别为92.75%和96.13%,均显著高于蚕豆蚜僵蚜的羽化率62.03%(F3,8=33.09,P<0.01)。综合两者考虑,桃蚜更适合作为菜蚜茧蜂的替代寄主,易于茧蜂繁殖。但其能否作为系统替代寄主仍需要通过对目标作物的潜在性危害评估才能确定。
2.1.2 替代寄主对目标作物的潜在危害评价
关于菜蚜茧蜂的替代寄主对目标作物潜在危害的观测结果如表1 所示。在实验设置后的第五天,距离烟草0.5 m 以及1 m 的部分菜薹植株上开始出现桃蚜,但虫量非常少,分别只有0.67 头/株、1.00 头/株。到了第10 天和第15 天,这两距离内的菜薹植株上桃蚜数量均有增加,但总体仍然很少,最多只有30.67 头/株菜薹以及22.00 头/株菜薹。另外,距离接种有桃蚜的烟草植株3 m,5 m 的菜薹植株上,桃蚜迁移量更少,只有到了第15 天才有极少数桃蚜。这说明桃蚜不会对目标蔬菜(菜薹)构成大的潜在危害。因此,我们选择桃蚜作为菜蚜茧蜂的替代寄主。
2.2 载体植物筛选
2.2.1 替代寄主对载体植物的取食偏好性(选择性试验)
在6种候选载体植物(烟草、玉米、生菜、甘蓝、蚕豆和菜薹)中,桃蚜对烟草的取食偏好性明显高于其他5种植物,有49.00%的受试桃蚜倾向于取食烟草,而只有16.30%倾向于取食生菜,菜薹为10.00%,甘蓝为8.40%,蚕豆为5.00%,玉米仅为1.80%(图1)。24 h、48 h 分别累计有3 头和5 头桃蚜死亡。综合24 h 和48 h两个调查时间段来看,桃蚜更倾向于在烟草上进行生长繁殖。

图1 桃蚜对候选载体植物的偏好性Fig.1 The preference of Myzus persicae to candidate blanker plants
2.2.2 替代寄主在载体植物上的取食适应性(非选择性试验)
非选择性试验的结果表明(图2),桃蚜在烟草上的生存繁殖能力明显高于其他植株,蚜虫接种7 d 后其数量从50 头上升至237 头,而除菜薹外其他几种植物在蚜虫接入后的第二天就开始相继出现死亡现象,到第5 天玉米、生菜以及蚕豆植株上的蚜虫基本全部死亡。相比之下,虽然桃蚜接入到菜薹植株上之后也出现死亡现象,但经过两天的适应期,桃蚜开始在菜薹上进行繁殖,后期其数量开始逐渐回升,说明桃蚜对于菜薹是有一定危害的。但也可看到桃蚜在菜薹上的生长繁殖速率非常缓慢,不会造成大危害。
综合替代寄主桃蚜对各载体植物的取食偏好性以及生长适应性结果,选择烟草作为系统的载体植物。
2.3 蜂蚜比的确定以及菜蚜茧蜂载体植物系统的构建
2.3.1 蜂蚜比的确定
在实验设置的5个蜂(♀)蚜比中,其寄生率与羽化率均以1∶25 的处理组最高,分别达到96.00%和98.55%;1∶50 和1∶100 两个蜂(♀)蚜比之间相差不大,其寄生率分别为71.33% 和69.00%,羽化率分别为93.33% 和92.92%;蜂(♀)蚜比下降到1∶150 和1∶200 时,寄生率均有较明显的下降,分别为59.11%和58.83%,两者相差不大。1∶200 处理组的羽化率则明显低于其他4 组,只有82.03%(图3)。

图2 桃蚜在候选载体植物上的数量变化Fig.2 The number of Myzus persicae in candidate blanker plants
综合考虑寄生率、羽化率以及用蜂量,选择1∶100作为该载体植物系统构建的蜂(♀)蚜比。
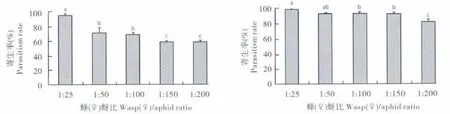
图3 不同蜂(♀)蚜比下菜蚜茧蜂对桃蚜的寄生率和羽化率Fig.3 The parasitism and eclosion rates of Diaeretiella rapae to Myzus persicae in different wasp(♀)aphid ratios
2.3.2 菜蚜茧蜂载体植物系统的构建
以1500 头、2000 头和2500 头三个不同数量梯度的起始桃蚜量构建载体植物系统,结果表明,1500 头、2000 头和2500 头最终获得的桃蚜僵蚜数量分别是1907.50、2770.00 和3297.50 头,可见在一定范围内起始桃蚜数量越多,系统最终产生的僵蚜数量也越多;起始桃蚜数量为1500 头和2000 头时,其产僵蚜的时间更长,一直持续到接入菜蚜茧蜂的第28 天,2500 头组只有21 d。随机抽取各组50 头僵蚜测羽化率,重复3 次,得到三组的羽化率分别为85.33%、83.33%、84.72%,无显著差异(F3,8=0.84,P>0.05)。综合考虑系统僵蚜数量、持续时间、前期起始桃蚜的繁殖时间以及数量庞大的桃蚜影响烟草存活,我们选取起始桃蚜量2000 头来构建载体植物系统。

表2 以不同的桃蚜起始数量构建载体植物系统Table 2 The banker plant systems for Diaeretiella rapae with different beginning amount of Lipaphis erysimi
2.4 菜蚜茧蜂载体植物系统对目标害虫的控制作用
2.4.1 系统繁殖的菜蚜茧蜂对菜蚜的寄生率及僵蚜羽化率
在株高约10 cm 菜薹植株上接入3-4 龄无翅、无病、未被寄生的菜蚜100 头,1 d 后接入1 头由载体植物系统繁殖所得的已交配的雌性菜蚜茧蜂,任其自由寄生24 h,24 h 后将雌蜂移出。结果表明(表3),1 头菜蚜茧蜂对目标害虫菜蚜的寄生率达71.17%,能有效抑制菜蚜种群的发展与扩大繁殖。该菜蚜茧蜂寄生的菜蚜僵蚜的羽化率为82.78,略低于寄生桃蚜的僵蚜羽化率(92.93%)。

表3 系统繁殖菜蚜茧蜂对菜蚜的寄生率和羽化率Table 3 The parasitism and eclosion rates of Diaeretiella rapae on the target pest Lipaphis erysimi
2.4.2 菜蚜茧蜂载体植物系统对温室菜蚜的防治效果
当烟草植株上的桃蚜达到2000 头时,接入20头已交配的雌性菜蚜茧蜂,构建菜蚜茧蜂载体植物系统。选取接蜂7 d 后的菜蚜茧蜂载体植物系统备用。在温室1 m×1 m×1 m 的白色网笼中放置菜薹植株15 盆,分别向每株菜薹上接入菜蚜20头,自由繁殖15 d 后,在网笼中央置入1 株菜蚜茧蜂载体植物系统,结果表明(图4):处理组引入载体植物系统当天,处理组和对照组菜薹植株上的菜蚜平均数量无显著差异;引入菜蚜茧蜂载体植物系统后第7 天和14 天,载体植物系统能显著降低菜薹上的菜蚜数量,减退率分别为83.11%和95.80%,并且在实验中我们观察到处理组的菜薹植株能正常生长,而对照组菜薹则被数量庞大(第7 天时为1299.67 头/株)的菜蚜侵蚀,第14天时已濒临死亡。这说明所构建的菜蚜茧蜂载体植物系统在温室中能很好地防治菜蚜。
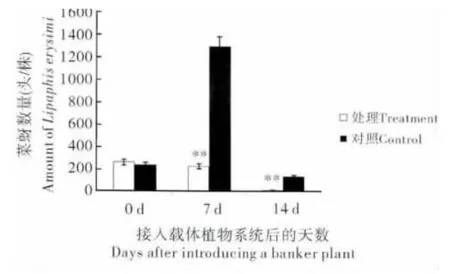
图4 菜蚜茧蜂载体植物系统在温室中防控菜蚜的效果Fig.4 Effectiveness of the Diaeretiella rapae banker plant system on Lipaphis erysimi in a greenhouse
3 结论与讨论
载体系统构建过程中,首先要确定载体植物、有益生物和替代寄主。本文所用的菜蚜茧蜂,已有防治蚜虫方面的报道(刘宏谋,1980)。在筛选替代寄主方面,国内外都少有研究报道,主要的成功案例都是在茧蜂的寄主范围内直接确定一种非目标害虫作为替代寄主(Wong et al.,2012)。本文以常见的三种蚜虫(桃蚜、甘蓝蚜和蚕豆蚜)为备选替代寄主,来筛选最佳替代寄主。通过比较菜蚜茧蜂对这三种蚜虫的平均寄生率、羽化率,最终选择繁殖菜蚜茧蜂数量最多的桃蚜为替代寄主。同时,我们发现桃蚜对目标作物菜薹潜在危害不大(表1)。所以,综合这两方面的研究,确定桃蚜为替代寄主。在载体植物筛选方面,国际上应用成功的载体植物系统大都是直接用替代寄主的目标植物作为载体植物(Frank,2010)。本文为确定载体植物,用包括桃蚜的目标植物烟草在内的6种常见植物进行筛选。由于桃蚜一般都是在烟草植株上进行繁殖,因此在对其进行取食偏好性实验时,存在桃蚜对烟草有习惯性取食倾向的可能,对实验结果造成一定影响。但从桃蚜对各候选载体植物的适应性试验结果可以看出,经过7 d 的适应,桃蚜只能在烟草植株上进行大量繁殖,而在其他植株上则相继死亡或只维持在较低数量水平(图2),因此对候选载体植物进行筛选是有效的。
雌性菜蚜茧蜂接蜂量与接蜂时桃蚜数量的比例(蜂(♀)蚜比)对桃蚜的繁蜂效果有着非常重要的影响。当蚜虫量较少而接蜂量过大时,蚜虫会被过量寄生而失去必要的繁殖增长能力,使得在繁蜂后期因为蚜虫的缺乏而失去持续繁殖菜蚜茧蜂的能力;而当蚜虫量较大而接蜂量过少时,蚜虫的过量繁殖会使得寄生植株的嫩叶生长停滞、植株营养不良或大量形成有翅蚜虫,致使菜蚜茧蜂的繁殖效率低下。而且当少量菜蚜茧蜂面对大量蚜虫进行寄生时,寄生效果也会降低,致使部分菜蚜茧蜂后代营养不良而不利于进一步寄生(邓建华等,2006)。因此,在菜蚜茧蜂载体植物系统构建过程中,为获得较佳的繁蜂效果和蚜虫防治效果,必须对蜂(♀)蚜比进行研究确定。本实验通过设计五个蜂(♀)蚜比梯度进行寄生实验,通过检测寄生率、羽化率,比较产蜂量,确定1∶100 为最适蜂(♀)蚜比。
本文所构建的菜蚜茧蜂载体植物系统,在28 d内能产生大量的菜蚜茧蜂。通过僵蚜数量和调查所得的平均羽化率计算(以2000 头起始蚜虫为例),每株载体植物系统可产生2300 多头菜蚜茧蜂。在1 m3的养虫笼内,可容纳10 株左右的载体植物系统,总共能产生23000 多头菜蚜茧蜂。而且构建的菜蚜茧蜂载体植物在温室中对菜蚜的控害效果高达95.80%,对菜蚜的防治效果很好。但是,这些菜蚜茧蜂载体植物系统在田间的防治效果如何?还有待进一步研究。
在利用载体植物系统繁殖菜蚜茧蜂的过程中,重寄生蜂是限制菜蚜茧蜂大量繁殖的一个重要因素,菜蚜茧蜂的主要重寄生蜂是蚜虫宽缘金小蜂Pachyneuron aphidis(Bouche),周子方等(2011)在田间小棚进行烟蚜茧蜂繁殖时发现,在烟蚜茧蜂大量形成的初期,重寄生蜂的数量较少,仅为1.7%,但到了繁蜂后期,僵蚜的重寄生率已达到11.5%,繁蜂质量大受影响。蚜虫宽缘金小蜂Pachyneuron aphidis 对所构建的系统有一定的危害性,为避免重寄生的威胁,我们进行菜蚜茧蜂接种时严格杜绝重寄生蜂;尽量缩短繁蜂周期,以免重寄生蜂有机可乘;及时清除繁蜂后的烟草植株;在载体植物系统中发现重寄生蜂后,立即隔离此系统并持续捕杀重寄生蜂至整个系统不再产菜蚜茧蜂。而在实际应用中,当整个载体植物系统呈开放式状态,如何防止重寄生蜂对菜蚜茧蜂僵蚜的寄生,还有待进一步的研究。
References)
Deng JH,Wu XF,Song CM,et al.Rearing effect of Aphidius gifuensis with a nylon-net coved cages in tobacco fields[J].Journal of Southwest Agricultural University,2006,28(1):66-69.[邓建华,吴兴富,宋春满,等.田间小棚繁殖烟蚜茧蜂的繁蜂效果研究[J].西南农业大学学报(自然科学版),2006,28(1):66-69]
Frank SD.Biological control of arthropod pests using banker plant systems:Past progress and future directions[J].Biological Control,2010,52:8-16.
Jandricic SE,Frank SD.The effect of banker plant species on the fitness of Aphidius colemani Viereck and its aphid host(Rhopalosiphum padi L.)[J].Biological Control,2010,52:8-16.
Li MF,Zhang YP,Wang XZ.Investigation of the effect to prevent and cure to aphids to rearing Aphidius gifuensis[J].Chinese Agricultural Science Bulletin,2006,22(2):343-346.[李明福,张永平,王秀忠.烟蚜茧蜂繁育及对烟蚜的防治效果探索[J].中国农学通报,2006,22(2):343-346]
Liu HM.Aphids occurrence and control technique in rape field[J].Journal of Henan Agricultural Science,1980,4:15-16.[刘宏谋.油菜蚜虫的发生和防治[J].河南农林科技,1980,4:15-16]
Long XJ,Lu Z.Control of Myzus persicae by using Aphidius gifuensis ashmaed[J].Hunan Agricultural Sciences,2012,1:80-82.[龙宪军,卢钊.利用烟蚜茧蜂防治烟蚜的技术研究[J].湖南农业科学,2012,1:80-82]
Mulder S,Hoogerbrugge H,Altena K,et al.Biological pest control in cucumbers in the Netherlands[J].International Organization for Biological Control Bulletin,1999,22:177-180.
Nagasaka K,Takahasi N,Okabayashi T,et al.Impact of secondary parasitism on Aphidius colemani in the banker plant system on aphid control in commercial greenhouses in Kochi,Japan[J].Applied Entomology and Zoology,2010,45(4):541-547.
Ohta I,Honda K.Use of Sitobion akebiae(Hemiptera:Aphididae)as an alternative host aphid for a banker-plant system using an indigenous parasitoid,Aphidius gifuensis(Hymenoptera:Braconidae)[J].Applied Entomology and Zoology,2010,45(2):233-241.
Tang PH.Host-Mediated RNAi to Control Aphids[D].Chongqing:Chongqing University,2012.[唐平华.利用宿主介导RNAi 进行蚜虫防治的研究[D].硕士学位论文,重庆:重庆大学,2012]
Wang Y,Huang W,Siemann E,et al.Lower resistance and higher tolerance of invasive host plants:biocontrol agents reach high densities but exert weak control[J].Ecological Applications,2011,21(3):729-738.
Wong SK,Frank SD.Influence of banker plants and spiders on biological control by Orius insidiosus(Heteroptera:Anthocoridae)[J].Biological Control,2012,63(2):181-187.
Xiao YF,Chen JJ,Cantliffe D,et al.Establishment of papaya banker plant system for parasitoid,Encarsia sophia(Hymenoptera:Aphilidae)against Bemisia tabaci(Hemiptera:Aleyrodidae)in greenhouse tomato production[J].Biological Control,2011,58(3):239.
Xiao YF,Mao RQ,Shen GQ.Bankerplant system:a new approach for biological control of arthropod pests[J].Chinese Journal of Biological Control,2012,28(1):1-8.[肖英方,毛润乾,沈国清.害虫生物防治新技术——载体植物系统[J].中国生物防治学报,2012,28(1):1-8]
Yu DP,Liu JX,Sun XH.Reasons of aphids re-occurrence and comprehensive control technique[J].Modern Agricultural Science and Technology,2009,11:130-132.[于东坡,刘金象,孙秀宏.菜蚜重发的原因及综合防治技术[J].现代农业科技,2009,11:130-132]
Zhou ZF,Ren W,Zhou JH,et al.Technical obstacles and solutions for Myzus persicae control by large-scale reproduction of Aphidius gifuensis[J].Journal of Anhui Agricultural Science,2011,16:9659-9661.[周子方,任伟,周冀衡,等.规模化应用烟蚜茧蜂防治烟蚜的主要技术障碍及应对方法[J].安徽农业科学,2011,16:9659-9661]
猜你喜欢
资源导刊(2022年12期)2023-01-03
果农之友(2022年9期)2022-09-27
小天使·一年级语数英综合(2022年5期)2022-05-25
南方农业(2021年28期)2021-12-25
果农之友(2021年10期)2021-11-03
发明与创新·小学生(2021年5期)2021-08-23
发明与创新(2021年19期)2021-05-07
文苑(2020年3期)2020-04-21
小学生导刊(2018年31期)2018-12-06
湖北植保(2017年6期)2017-12-23
