
雌蛛拖丝在星豹蛛和拟环纹豹蛛物种识别中的作用
2015-10-19 07:16曾阳梁宏合江庆生陈建刘凤想焦晓国
湖北大学学报(自然科学版) 2015年3期
曾阳,梁宏合,江庆生,陈建,刘凤想,焦晓国
(1.行为生态与进化研究中心,湖北生物资源绿色转化协同创新中心,湖北大学生命科学学院,湖北 武汉 430062;2.广西壮族自治区亚热带作物研究所,广西 南宁 530002;3.湖北荆州市农业科学院,湖北 荆州 434000)
有关蜘蛛嗅觉通讯(化学通讯)研究的广度和深度远落后于昆虫[1-3].近年来越来越多的研究表明:在蜘蛛类群中,除了视觉、听觉和触觉等感觉模式外,嗅觉也在通讯中发挥重要作用.蜘蛛化学通讯可分为种间化学通讯和种内化学通讯.通过种间化学信息物质,蜘蛛可寻找猎物、逃避捕食者以及采用化学拟态模拟猎物的性信息素以提高捕食效率[4];通过种内化学通讯,蜘蛛可寻找和定位配偶、发出求偶信号以及利用化学信息物质(变化)评价配偶质量[1-3].蜘蛛雌雄蛛均可释放种内化学信息物质(性信息素),但主要是通过雌蛛释放性信息素,然后被雄蛛步足及触肢上特殊的化学感受器感知,进而雄蛛开始搜寻雌蛛,在雌蛛面前展示求偶动作,进一步完成交配,以达到成功繁殖的目的[1-3,5].
有关蜘蛛性信息素的研究目前集中在生物测定阶段,主要包括性信息素的有无以及推测其可能的生物学功能[2-3],对蜘蛛性信息素的释放腺体及信息素生物合成与转化等知之甚少.目前仅鉴定了5种蜘蛛的性信息素分子[6-10].在狼蛛科蜘蛛中,有关其性信息素的研究虽有报道[10-17],但主要也集中在生物测定研究层次.已有的测定结果表明:狼蛛科蜘蛛性信息素雌性能释放[10-15],雄性也能释放[16];性信息素既有挥发性的[15-16],也有接触性的[10-14,17],但以后者多见[2-3].接触型性信息素主要分布于蜘蛛的体表和拖丝上,而其中又以拖丝上接触型性信息素更为常见[2-3],雄蛛不但可以利用雌蛛释放的拖丝上的性信息素远距离定位雌蛛,也可以在种内性别、日龄和配偶生殖状态的鉴别中发挥重要作用[11,14].
狼蛛科蜘蛛性信息素的研究主要集中于裂腹蛛属[2,12-14,17],在豹蛛属中研究鲜见[2,11,15].本研究以狼蛛科豹蛛属两种游猎型蜘蛛——星豹蛛(Pardosaastrigera)和拟环纹豹蛛(Pardosapseudoannulata)为对象,研究星豹蛛和拟环纹豹蛛雌蛛释放的拖丝分别对星豹蛛和拟环纹豹蛛雄蛛求偶行为的影响,探讨两种豹蛛雌蛛拖丝上接触型性信息素在蜘蛛种内识别中的作用.
1 材料与方法
1.1 实验材料2009年3月于湖北省武汉市华中农业大学棉花试验田采集越冬代星豹蛛和拟环纹豹蛛亚成蛛,单头分装于玻璃试管(长10 cm,直径2 cm)中,管底用吸足水的海绵保湿,脱脂棉封口,以防蜘蛛逃逸.蜘蛛采回后置于光照培养箱中饲养,培养条件控制为25℃、70%~90%的RH和14L:10D光照.以室内饲养的黑腹果蝇(Drosophilamelanogaster)饲养蜘蛛,每只蜘蛛每次喂食20~30只黑腹果蝇,每3天1次.每两周更换一次试管,以保证试管的清洁.每天早晚观察试管中蜘蛛蜕皮情况,记录蜘蛛的蜕皮时间及成熟时间,成熟后备用.实验用蜘蛛都为成熟未交配的蜘蛛.
1.2 实验方法我们已对星豹蛛和拟环纹豹蛛雄蛛求偶行为进行了详细的研究.星豹蛛雄蛛面对雌蛛时,第一对步足同时上扬,最高处与身体长轴垂直,然后下放,接触地面,同时整个身体伴随“俯卧撑”式动作[18].拟环纹豹蛛雄蛛面对雌蛛时,第一对步足同时水平前伸,伴随高频颤动,然后下放,接触地面[19],同时雄蛛触肢同步上扬,然后同步下放,伴随触肢向外打开和收拢,如同船夫的“划桨”动作.整个身体也伴随“俯卧撑”式运动[19].为了研究方便,我们以两种豹蛛雄蛛第一对步足伸展和“俯卧撑”为求偶评价动作.
采用培养皿滤纸法分别收集成熟后(未交配)5~7日龄星豹蛛和拟环纹豹蛛雌蛛的拖丝(样本数n=50).收集每种雌蛛拖丝时,把单头雌蛛置于底部垫有滤纸的培养皿(直径9 cm)中24 h,每只雌蛛仅使用1次.培养皿盖内侧正上方放一蘸水的棉球保湿,实验室温度控制25℃,14L:10D光照,24 h后把培养皿中雌蛛移走.然后随机选择成熟后(未交配)3~5日龄星豹蛛或拟环纹豹蛛雄蛛各50只,每只雄蛛也仅使用1次.采用小试管把单头雄蛛轻轻引入上述收集有不同类型拖丝的培养皿中.从引入开始计时,持续5 min,分别记录下述指标:1)雄蛛第一对步足伸展潜伏时间(latency to leg raising),即从引入雄蛛开始到雄蛛在培养皿中开始伸展第一对步足的时间;2)雄蛛身体“俯卧撑”式震动潜伏时间(latency to body shaking),即从引入雄蛛开始到雄蛛开始在培养皿中身体“俯卧撑”式震动时间;3)雄蛛第一对步足伸展强度(foreleg raises),即从引入雄蛛开始,雄蛛在5min内第一步足同时向前上方伸展次数(从第一对步足离开地面到其在空中伸展后接触地面为伸展1次).4)雄蛛身体“俯卧撑”式震动强度(body shakes),即从引入雄蛛开始,雄蛛在5min内作“俯卧撑”式震动的次数.如果雄蛛引入后5min内无求偶动作,则该数据剔除,不参与统计.
1.3 统计方法在不同雌蛛释放的拖丝对雄蛛求偶行为影响实验中,雄蛛5min内潜伏时间、第一对步足伸展强度和身体震动强度等数据在进行方差分析前先检验数据正态性和方差齐性.检验结果发现当前研究数据均非正态分布,因此我们采用非参数Kruskal-WallisH分析方法.如差异显著,不同处理间差异采用Mann-Whitney U测验进行两两比较.所有数据均以平均值±标准误表示,采用SPSS 11.5统计软件进行分析.
2 结果与分析
不同雄蛛在不同雌蛛拖丝上第一对步足伸展潜伏期比较结果见图1 A.拟环纹豹蛛和星豹蛛雄蛛在同种雌蛛释放的拖丝上第一对步足伸展潜伏期都显著短于在种间雌蛛拖丝上的潜伏期(Kruskal-Wallis testχ2=65.599,df=3,P<0.001).不同雄蛛在不同雌蛛拖丝上身体震动潜伏期比较结果见图1B.拟环纹豹蛛和星豹蛛雄蛛在同种雌蛛释放的拖丝上身体震动潜伏期都显著短于在种间雌蛛拖丝上的潜伏期(Kruskal-Wallis testχ2=69.989,df=3,P<0.001).
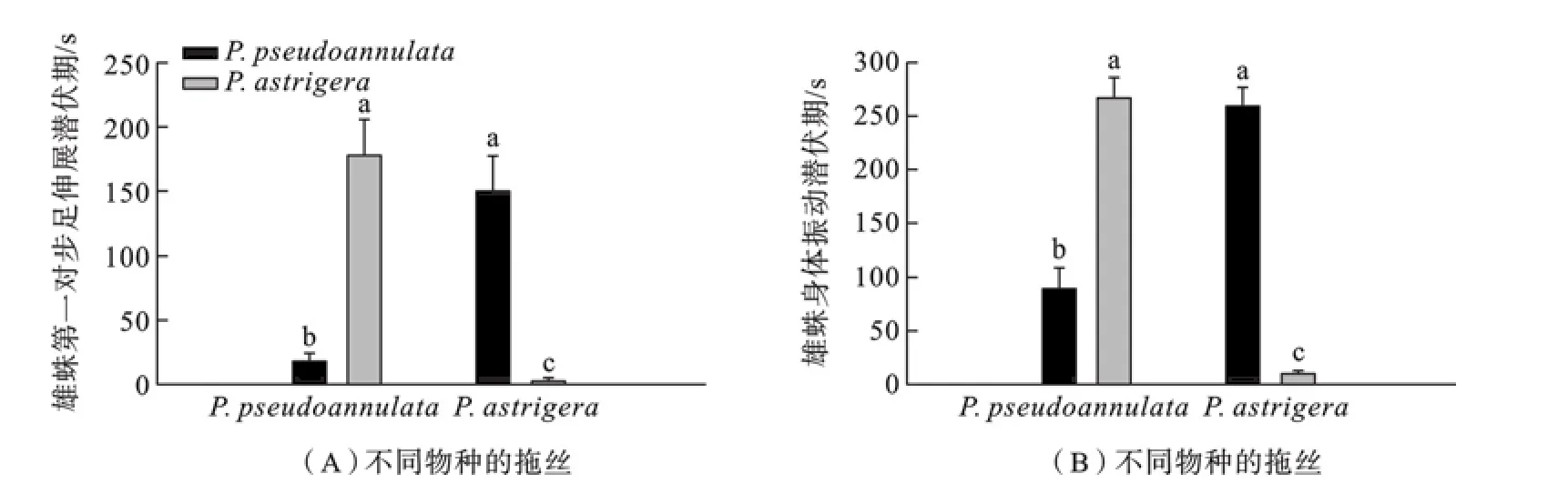
图1 星豹蛛和拟环纹豹蛛雌蛛的拖丝对雄蛛第一对步足伸展潜伏期(A)和身体震动潜伏期(B)的影响(不同字母表示差异显著)

图2 星豹蛛和拟环纹豹蛛雌蛛的拖丝对雄蛛第一对步足伸展强度(A)和身体震动强度(B)的影响(不同字母表示差异显著)
不同雄蛛在不同雌蛛拖丝上第一对步足伸展强度比较结果见图2 A.拟环纹豹蛛和星豹蛛雄蛛在同种雌蛛释放的拖丝上第一对步足伸展强度都显著高于在种间雌蛛拖丝上的伸展强度(Kruskal-Wallis testχ2=73.670,df=3,P<0.001).不同雄蛛在不同雌蛛拖丝上身体震动强度比较结果见图2B.拟环纹豹蛛和星豹蛛雄蛛在同种雌蛛释放的拖丝上身体震动强度都显著高于在种间雌蛛拖丝上的震动强度(Kruskal-Wallis testχ2=81.040,df=3,P<0.001).
3 讨论
狼蛛科蜘蛛采用多种通讯方式进行种内和种间信息交流,它们已逐渐成为动物通讯的重要研究类群[1].但研究重点主要集中于狼蛛视觉和听觉通讯方式,化学通讯研究不甚深入.在自然条件下,视觉和听觉主要在近距离范围起作用,远距离通讯仍需借助化学信息[2-3].根据Trivers的理论,雌性在生殖上的投资通常大于雄性,导致配偶选择主要集中于雌性对雄性的选择[20].但近年来研究发现,雄性的生殖行为也需付出相当代价,如精子的产生、搜寻和竞争配偶、求偶和交配行为等[21].雄性的配偶选择越来越引起学者的重视,特别是化学信息在雄性配偶选择中的作用[22].化学信息在雄性择偶中的作用由低到高分为3个层次,物种识别、配偶识别和配偶质量评价[22].其中物种识别是两性动物进行生殖的基础,可以避免因种间杂交而导致的不育.
在自然条件下,游猎型蜘蛛雌雄个体之间的空间分布距离远远超过其视力范围,如果雄蛛随机搜寻雌蛛,必然浪费时间和能量,也更容易遭受捕食者的攻击,对其生存和繁殖都不利[23].因此,雄蛛必须利用其他信息准确高效定位雌蛛[23].越来越多的研究结果表明,对于游猎型蜘蛛,雄蛛主要借助雌蛛释放的拖丝上的物理或化学信息定位雌蛛.例如,蟹蛛(Misumenavatia)雄蛛主要通过雌蛛释放的拖丝物理性状(直径和弹性强度等)定位雌蛛,但不能辨别雌蛛的交配状态[24].已有的研究表明,狼蛛科雄蛛主要通过雌蛛释放于拖丝上的化学信息物质定位雌蛛[11-14,17].当前的研究发现,拟环纹豹蛛和星豹蛛雄蛛在同种雌蛛释放的拖丝上求偶潜伏期都显著短于在异种雌蛛拖丝上的潜伏期,而两种雄蛛在同种雌蛛拖丝上求偶强度都显著高于在异种雌蛛拖丝上的求偶强度,表明星豹蛛和拟环纹豹蛛雄蛛能通过同种雌蛛释放的拖丝上性信息素进行种内识别.可见,星豹蛛和拟环纹豹蛛雌蛛释放的拖丝上化学信息物质具有种的特异性,这与狼蛛科裂腹蛛属中报道一致[11-14,17].可以推测雄蛛对雌蛛拖丝上化学信息物质的识别是一种重要的交配前隔离机制.
化学信息在配偶质量评价中也具重要作用[22].已有研究发现,狼蛛科裂腹蛛属雌蛛交配后其性接受行为下降[14],同类相食行为上升.同种雄性具有通过雌性拖丝上化学信息物质鉴别雌性交配状态的能力,优先选择追踪成熟没有交配的雌蛛释放的拖丝[11,14].加之雌蛛狼蛛通常一生只交配1次[18-19],因此雄性狼蛛偏好选择成熟没有交配过的雌蛛释放的拖丝,不仅可以获得交配的机会,而且可以避免被同种已交配雌蛛的捕杀.
[1]Huber BA.Sexualselection research on spiders:progress and biases[J].BiolRev,2005,80:363-385.
[2]GaskettAC.Spider sex pheromones:emission,reception,structures,and functions[J].BiolRev,2007,82:27-48.
[3]Schulz S.Spider pheromones—a structuralperspective[J].JChem Ecol,2013,39:1-14.
[4]Gemeno C,Yeargan K V,Haynes K F.Aggressive chemical mimicry by the bolas spiderMastophorahutchinsoni:identification and quantification ofamajor prey’s sex pheromone components in the spider’s volatile emissions[J].JChem Ecol,2000,26:1235-1243.
[5]Jiao X G,Chen Z Q,Du H Y,et al.Chemoreceptors distribution and relative importance ofmale forelegs and palps in intersexual chemical communication of the wolfspiderPardosaastrigera[J].Chemoecology,2011,21:45-49.
[6]Schulz S,ToftS.Identification ofa sex pheromone from a spider[J].Science,1993,260:1635-1637.
[7]Papke M,Riechert SE,Schulz S.An airborne female pheromone associated withmale attraction and courtship in a desert spider[J].Anim Behav,2001,61:877-886.
[8]Tichy H,Gingl E,Ehn R,et al.Female sex pheromone of a wandering spiderCupienniussalei:identification and sensory reception[J].JComp Physiol A,2001,187:75-78.
[9]Xiao Y H,Zhang JX,Li SQ.A two-component female-produced pheromone of the spiderPholcusbeijingensis[J].J Chem Ecol,2009,35:769-778.
[10]Chinta SP,Goller S,Lux J,etal.The sex pheromone of thewasp spiderArgioperuennichi[J].Angew Chem Int Ed,2010,49:2033-2036.
[11]吴俊,焦晓国,陈建,等.星豹蛛雌蛛性信息素的行为学证据[J].动物学报,2007,53:994-999.
[12]Roberts JA,Uetz GW.Chemicalsignaling in awolfspider:a test of ethospecies discrimination[J].JChem Ecol,2004a,30:1271-1284.
[13]Roberts JA,Uetz GW.Species-specificity of chemical signals:silk source affects discrimination in awolf spider(Araneae,Lycosidae)[J].JInsect Behav,2004b,17:477-491.
[14]Roberts J A,Uetz G W.Information content of female chemical signals in a wolf spiderSchizocosaocreata:male discrimination of reproductive stateand receptivity[J].Anim Behav,2005,70:217-223.
[15]Searcy L E,Rypstra A L,Persons M H.Airborne chemical communication in thewolf spiderPardosamilvina[J].JChem Ecol,1999,25:2527-2533.
[16]Aisenberg A,Baruffaldi L,González M.Behavioural evidence ofmale volatile pheromones in the sex-role reversed wolf spidersAllocosabrasiliensisandAllocosaalticeps[J].Naturwissenschaften,2010,97:63-70.
[17]Baruffaldi L,Costa FG,Rodríguez A,et al.Chemical communication inSchizocosa malitiosa:evidence of a female contact sex pheromone and persistence in the field[J].JChem Ecol,2010,36:759-767.
[18]吴俊,焦晓国,陈建,等.星豹蛛求偶和交配行为[J].动物学杂志,2008,43:9-12.
[19]陈占起,焦晓国,陈建,等.拟环纹豹蛛求偶和交配行为的初步研究[J].湖北大学学报:自然科学版,2009,31:176-179.
[20]TriversR L.Sexualselection and the descen tof man[M].Chicago:Aldine,1972.
[21]Bonduriansky R.The evolution of malematechoice in insects:asyn the sisofideas and evidence[J].Biol Rev,2001,76:305-339.
[22]Johansson BG,Jones TM.The role of chemical communication inmate choice[J].BiolRev,2007,82:265-289.
[23]陈军,宋大祥.狼蛛科蜘蛛的繁殖行为[J].蛛形学报,1999,8:55-62.
[24]Leonard AS,MorseDH.Line-following preferences of malecrab spidersMisumenavatia[J].Anim Behav,2006,71:717-724.
猜你喜欢
作文小学高年级(2021年11期)2021-12-22
红蜻蜓·低年级(2021年9期)2021-11-05
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
红领巾·探索(2020年5期)2020-05-19
昆虫学报(2020年1期)2020-03-03
小朋友·快乐手工(2018年3期)2018-04-22
红领巾·探索(2018年12期)2018-01-26
小学阅读指南·低年级版(2017年6期)2017-06-12
中国马铃薯(2015年3期)2015-12-19
