
木间韧皮部的定义、分布、发育和生理功能
2015-09-09 21:36张玉秀刘培卫
湖北农业科学 2015年15期
关键词:生理功能
张玉秀+刘培卫

摘要:木间韧皮部是植物异常次生结构的一种。针对木间韧皮部的定义和分布存在着很大分歧以及它的生理功能还不清楚的现状,对木间韧皮部的定义、分布和发育方式等方面进行了简要阐述,并对其可能具有的生理功能进行了推测。旨在为木间韧皮部术语规范使用提供帮助,为进一步探索含有木间韧皮部的植物资源提供参考,为深入研究木间韧皮部的功能奠定基础。
关键词:木间韧皮部;内涵韧皮部;异常次生结构;生理功能
中图分类号:Q944.3+2 文献标识码:A 文章编号:0439-8114(2015)15-3589-04
DOI:10.14088/j.cnki.issn0439-8114.2015.15.002
Abstract: Interxylary phloem is one of the anomalous secondary structures. There are different opinions on definition and distribution of interxylary phloem, and its physiological function is also unclear. So the definition, distribution and development were reviewed, and the possible physiological function of the interxylary phloem was speculated in this paper to provide a reference for scientific use of the term and further exploration of plant species containing interxylary phloem, and to lay the foundation for further study of the function of interxylary phloem.
Key words: interxylary phloem; included phloem; anomalous secondary structure; physiological function
收稿日期:2014-12-16
基金项目:中央级公益性科研院所基本科研业务专项(2013HNB01);海南省自然科学基金项目(314183);协和青年基金项目(33320140078)
作者简介:张玉秀(1984-),女,山东德州人,研究实习员,硕士,主要从事珍稀南药植物病虫害防治及栽培研究,(电话)18876005027(电子信箱)
313884523@qq.com;通讯作者,刘培卫,助理研究员,从事珍稀南药植物资源利用和开发研究,(电子信箱)brucelpw@aliyun.com。通常情况下,维管植物的次生加粗是由于形成层的分裂,向内产生木质部,向外产生韧皮部的结果。但有一些植物的次生加粗偏离了人们所熟知的这种加粗方式,从而出现了各种各样的结构,称为异常次生结构(Anomalous secondary structure)[1],木间韧皮部(Interxylary phloem)就属于此类。关于如何定义木间韧皮部,哪些植物类群中含有此种异常结构,至今已有大量的研究报道,但这些报道中相互矛盾的地方太多[1,2]。针对此种情况,本文对木间韧皮部的定义及其演变过程、哪些植物类群中含有木间韧皮部、木间韧皮部发育的方式、木间韧皮部的生理功能等方面进行了简要概述。
1 木间韧皮部的定义及演变过程
随着科学技术的发展,木间韧皮部的定义也处于不断的演变过程中。所以对木间韧皮部的描述和称谓有多种不同的版本。木间韧皮部被不同的学者称为内涵韧皮部与弥散型内涵韧皮部,甚至错误地称为木内韧皮部等。Fahn[1]认为木间韧皮部或内涵韧皮部是指由植物体内多个连续的形成层发育而来的,分布在次生木质部内部的异常韧皮部,并且认为藜科(Chenopodiaceae)、苋科(Amaranthaceae)和紫茉莉科(Nyctginaceae)等55个科的植物中具有此结构[1,3]。现在人们通常不采用此说法,而把这种由多个连续的形成层形成的结构称为同心环状排列的异常维管组织[4]或连续性形成层(Successive cambia)[2,5]。国际木材解剖学家协会(International Association of Wood Anatomist, IAWA)在1989年规定,将散生在次生木质部中的条带状的异常次生韧皮部的这种异常结构称为弥散型内涵韧皮部[6]。胡正海[4]把包埋在次生木质部中的异常次生韧皮部束称为内涵韧皮部或木间韧皮部。这两个定义仅考虑了异常韧皮部位于次生木质部内部的这种静态的表现形式,却没有考虑这种异常韧皮部的发育过程。所以根据这两个定义很难把木间韧皮部、木内韧皮部和连续性形成层区分开来,所以在某些报道中将它们混为一谈[1,2]。
Carlquist[2]综合前人的研究成果,明确提出木间韧皮部是指由单个维管形成层发育形成的,嵌入式的散生在植物根或茎的次生木质部中的股状或条带状异常次生韧皮部束。该定义不仅明确了木间韧皮部静态的表现形式,而且还特别指出木间韧皮部是由单个维管形成层发育而来的,所以依据此定义可以清楚地将木间韧皮部与其他类似的异常结构区分开来。仅从植物解剖学的角度出发,笔者认为此定义是目前最科学合理的,本文中所提及的木间韧皮部都以此定义为准。随着对木间韧皮部研究的深入,可能会出现更为科学和符合其本质的定义。
2 木间韧皮部在不同植物类群中的分布
笔者在前人研究报道的基础上,对具有木间韧皮部的植物种类进行了整理和归纳[2,5,7],发现至少在17个科约54个属的部分或全部物种中具有木间韧皮部(表1)。当然随着研究的深入,表1中的内容还需要进一步核实和修正。一般说来,如果某一物种中含有木间韧皮部,那么同一属中的其他物种也应该含有同样的异常结构[8],但Van[9]观察了40种风车子属(Combretum)植物,结果只在其中的20种植物中发现含有木间韧皮部,所以在寻找其他含有木间韧皮部的植物时这一规律仅供参考,不可盲信。
3 木间韧皮部的发育方式
虽然木间韧皮部是由单个维管形成层发育而来的,但研究发现它们的发育过程有3种方式[4,10]。
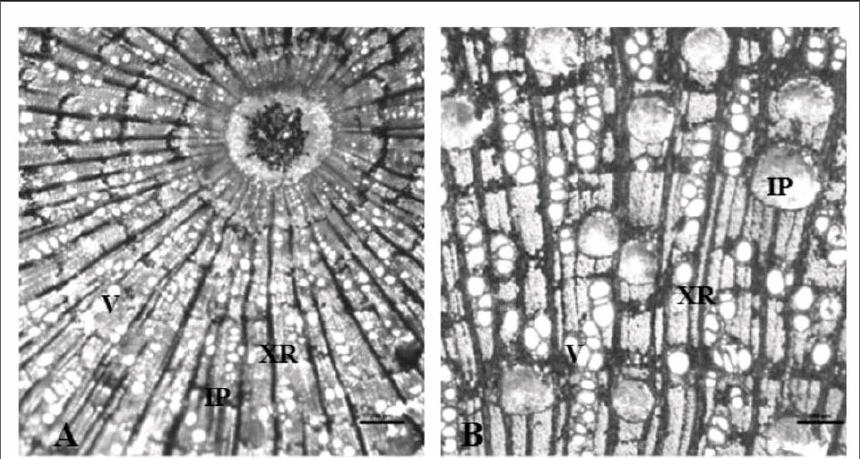
3.1.1 维管形成层异常活动形成的木间韧皮部 此类维管形成层除了向内产生木质部外,还在特定的时间和空间内间断的向内产生一些次生韧皮部的束。风车子属(Combretum)植物的木间韧皮部多以此种方式形成,所以这种方式又被称为风车子型(Combretum-subtype)[10]。沉香属(Aquilaria)的一些种类[23]也属于此种类型(图1A)。
3.1.2 维管形成层正常活动形成的木间韧皮部 马钱属(Strychnos)的木间韧皮部多以此种方式形成(图1B),所以此种类型又被称为马钱型(Strychnos-subtype)[8,10]。此类维管形成层与正常的维管形成层一样,也是向外产生次生韧皮部,向内产生次生木质部,但是在某一特定的时间内,部分形成层单向活动,只向外产生韧皮部,不产生次生木质部,这部分形成层以及它外侧产生的次生韧皮部一起陷入次生木质部内,然后在陷入的次生韧皮部束外方产生弥补形成层,使维管形成层又变成完整的环状,恢复正常的活动,最后形成束状木间韧皮部星散分布在次生木质部中[4]。
3.1.3 次生木质部薄壁组织中形成的木间韧皮部 次生木质部薄壁组织中形成的木间韧皮部[4]又称为刺茉莉型(Azima-subtype)[10]。此类木间韧皮部是由次生木质部内的部分薄壁细胞反分化形成的,在金虎尾科(Malpighiaceae)和柳叶菜科(Onagraceae)的某些属中的木间韧皮部就是以此种方式形成的[4,10]。
4 木间韧皮部的生理功能
木间韧皮部在植物中究竟具有什么样的生理功能至今还不清楚[2]。因为相对于整个植物界来说,只有少数的几种植物中具有木间韧皮部[2],即使在同一个属中,如风车子属[2,9],有些物种含有木间韧皮部,而有些却没有,但它们在生境或大小方面却没有明显的差异[2],所以阐明木间韧皮部的功能是一件复杂的事情。另外,目前发现的含有木间韧皮部的植物的经济价值都比较低,导致人们对其的研究较少。总之,要完全阐明木间韧皮部的功能还有很长的路要走。
虽然木间韧皮部在植物生长过程中究竟起到什么样的作用目前还没有定论,但人们根据木间韧皮部的解剖结构推测木间韧皮部在光合产物的运输和贮藏等方面具有重要的作用[2,24]。下面列举了木间韧皮部可能具有的几种生理功能。
4.1 光合产物的运输通道
通过解剖发现木间韧皮部在结构组成上与正常的次生韧皮部相同,所以大多数学者认为它的主要功能和正常的次生韧皮部类似,主要功能就是向下运输养料和光合产物[2,26]。人们对含有木间韧皮部的白木香剥除树皮后,白木香还能正常生长;风车子属(Combretum)中含有木间韧皮部植物与不含有木间韧皮部的植物相比,前者正常的次生韧皮部中几乎不含有筛管[10];马钱属(Strychnos)的老茎中,木间韧皮部所占的比例可达整个韧皮部的75%[9];具有木间韧皮部的植物往往具有巨大的果实或花朵[2]。根据这些研究结果,人们推测木间韧皮部不仅是辅助次生韧皮部向下运输养料和光合产物的通道,而且是养料和光合产物向下运输的高速公路[2,24]。
4.2 韧皮部的保护模式
木间韧皮部束位于次生木质内部,周围被木质化的纤维包裹,这种结构明显表明与正常次生韧皮部相比,木间韧皮部更不容易受到刺吸式昆虫或者其他外界伤害的影响。另外使君子科(Combretaceae)的风车子属(Combretum)、柳叶菜科(Onagraceae)的所有属以及瑞香科(Thymeleaceae)沉香属(Aquilaria)中的木间韧皮部都富含晶体,这些晶体能够对抗动物,避免植物被吃掉,这也间接地表明木间韧皮部不容易被破坏。因此木间韧皮部可以看作是处于保护模式下的韧皮部[2],从这个层面来看,木间韧皮部可能是更为进化的一种表现形式。
4.3 贮藏作用
淀粉粒在植物体内是一种很普遍的贮藏营养物质,人们发现沉香属木间韧皮部中分布着大量淀粉粒(图1A),而在柳叶菜科(Onagraceae)的木材中,淀粉粒一般不分布在木间韧皮部中,而是分布在木间韧皮部周围的木薄壁细胞中[2],所以人们推测某些植物中的木间韧皮部具有贮藏和运输两种功能,而在某些植物中可能是以运输功能为主[2]。
4.4 其他生理功能
除了上面列举的3种明显生理功能以外,木间韧皮部可能还在存储水分[25]、阻碍伤害扩大[23,25]和维持顶端优势[24]等方面具有重要的作用。
5 展望
木间韧皮部是相对比较常见的植物异常次生结构之一,目前对它的研究主要集中在结构解剖方面。它在植物的生长过程中具有什么样的生理功能,木间韧皮部的出现究竟是一种退化的表现,还是一种进化的形式,它在系统演化过程中处于什么样的位置等这些问题还尚属空白。
近年来,沉香的价格在国内外市场上一直居高不下,品质稍好的沉香价格堪比黄金。沉香属植物作为生产沉香的最主要的植物资源,随之成为国内外学者关注的焦点。木间韧皮部是沉香属植物普遍具有的结构特征,是沉香树脂产生和累积的主要场所[23],所以人们应该借助沉香热的机会,对沉香属木间韧皮部的生理功能及其在系统演化中的地位等问题运用转录组学、代谢组学和生理生化等技术手段进行深入研究,从而为揭示其他植物类群中木间韧皮部的实质提供理论依据。
致谢:感谢魏建和研究员对本文提出的指导意见和建议。
参考文献:
[1] FAHN A.“The Development of the Secondary Body in Plants with Interxylary Phloem.”Xylorama[M].Basel:Birkhauser Verlag,1985.58-67.
[2] CARLQUIST S. Interxylary phloem: Diversity and functions[J]. Brittonia,2013,62(2): 477-495.
[3] 黄桂玲.植物的异常次生生长[J].植物学通报,1988,5(1):9-13.
[4] 胡正海.植物解剖学[M].北京:高等教育出版社,2010.251-253.
[5] CARLQUIST S. Successive cambia revisited:ontogeny,histology, diversity,and functional significance[J]. Journal of the Torrey Botanical Society,2007,134(2):301-322.
[6] WHEELER E A, BAAS P, GASSON P E. IAWA list of microscopic features for hardwood identification[J]. IAWA Bull, 1989, 10: 219-332.
[7] CARLQUIST S. Comparative Wood Anatomy-Systematic, Ecological, and Evolutionary Aspects of Dicotyledon Wood[M]. New York:Springer-Verlag Berlin and Heidelberg, 2001.271-279.
[8] VAN VEENENDAAL W, DEN OUTER R. Development of included phloem and organization of the phloem network in the stem of Strychnos millepunctata (Loganiaceae)[J].IAWA Journal, 1993,14(3): 253-265.
[9] VAN VLIET G.Wood anatomy of the Combretaceae[J].Blumea.1979,25(2):141-223.
[10] DEN OUTER R., VAN VEENENDAAL W . Development of included phloem in the stem of Combretum nigricans (Combretaceae)[J]. IAWA Journal, 1995,16(2): 151-158.
[11] SINGH B. The origin and distribution of inter- and intraxylary phloem in Leptadenia[J]. Proceedings: Plant Sciences, 1943,18(3): 14-19.
[12] PATIL V, RAJPUT K. Structure and development of inter- and intraxylary phloem in Leptadenia reticulata (Asclepiadaceae)[J]. Polish Botanical Journal, 2008,53(1):5-13。
[13] SCOTT D.On some points in the anatomy of Ipomoea versicolor Meissn[J].Annals of Botany,1891,5(1):173-180.
[14] CARLQUIST S. Wood anatomy of some Gentianaceae: Systematic and ecological conclusions[J]. Aliso,1984,120(2): 573-582.
[15] LENS F,K?魡REHED P,BAAS S,et al. The wood anatomy of the polyphyletic Icacinaceae and their relationships within asteroids[J].Taxon,2008, 57(2): 525-552.
[16] SCOTT D,BREBNER G. On the anatomy and histology of Strychnos[J]. Annals of Botany, 1889,3(2): 275-302.
[17] MENNEGA A. Anatomy of the secondary xylem[M]. Berlin:Duncker & Humblot, 1980.112-161.
[18] PFEIFFER H. Handbuch der Pflanzenanatomie[M]. Berlin:Gebruder Borntraeger, 1926. 241-243.
[19] CHALK L, CHATTAWAY M. Identification of woods with included phloem[J]. Tropical Woods,1937, 50(1): 1-31.
[20] CARLQUIST S. Wood anatomy of onagraceae; further species; root anatomy; signi ficance of vestured pits and other structures in the dicotyledons[J]. Annals of the Missouri Botanical Garden,1983,69(2):755-769.
[21] CARLQUIST S. Wood and bark anatomy of Salvadoraceae: ecology, relationships, histology of interxylary phloem[J]. Journal of the Torrey Botanical Society,2002,129(1):10-20.
[22] CARLQUIST S. Types of cambial activity and wood anatomy in Stylidium (Stylidiaceae)[J]. American Journal of Botany, 1981,68(2): 778-785.
[23] RAO K, DAYAL R. The secondary xylem of aquilaria agallocha (Thymelaeaceae) and the formation of AGAR[J]. IAWA Bulletin,1992,13(2):163-172.
[24] Z?魷ZIMO E,TAMAIO N,VIEIRA C. Development of intraxylary phloem in the stem of Combretum rotundifolium (CombretaCeae)[J]. IAWA Journal, 2011,32(4): 14-24.
[25] ROBERT E,SCHMITZ N,BOEREN I, et al. Successive cambia: a developmental oddity or an adaptive structure[J]. PLOS ONE, 2011:6(1):e16558.
猜你喜欢
国外畜牧学·猪与禽(2022年1期)2022-04-23
昆明医科大学学报(2020年12期)2021-01-26
猪业科学(2018年4期)2018-05-19
广东饲料(2016年4期)2016-12-01
江苏农业科学(2016年2期)2016-04-11
广东饲料(2016年8期)2016-02-27
江苏农业科学(2015年10期)2015-12-23
宠物世界·狗迷(2015年7期)2015-05-30
中国粮油学报(2015年5期)2015-02-06
河南科技(2014年7期)2014-02-27
