
寡照对设施黄瓜光合特性的影响研究
2015-08-23 01:42张继波薛晓萍李鸿怡
海洋气象学报 2015年3期
张继波,薛晓萍,李鸿怡
(山东省气候中心,济南 250031)
寡照对设施黄瓜光合特性的影响研究
张继波,薛晓萍,李鸿怡
(山东省气候中心,济南 250031)
为研究寡照对设施黄瓜苗期及花果期叶片光合速率的影响,于2012年2—4月份在章丘伟丽种苗基地设计不同程度寡照处理试验。结果表明:随着寡照天数的增加,黄瓜叶片PRD-V,LsD-V,TrD-V和CD-V均呈上升趋势,差值越大,寡照对黄瓜叶片光合速率、气孔限制值、蒸腾速率和气孔导度的影响越大。寡照处理1~7d,黄瓜叶片PRD-V,TrD-V和CD-V迅速增大,寡照处理7d后,黄瓜叶片PRD-V,TrD-V和CD-V缓慢增大,并趋于稳定。寡照7d可视为设施黄瓜发生寡照胁迫的临界值,苗期黄瓜叶片光合特性受寡照影响大于花果期。
寡照;黄瓜;光合特性;日光温室
引言
黄瓜是中国北方节能型日光温室的主栽蔬菜之一,冬、春季普遍存在的亚适温寡照天气对黄瓜植株的光合作用影响较大,致使黄瓜植株矮小,细弱,发育迟缓,成熟期推迟,甚至减产。关于寡照对作物生理参数、生长发育及果实品质的影响,国内外学者已经做了大量研究。据报道,寡照胁迫显著降低水稻[1]、宝盖草属[2]、拟南芥[3]等作物的叶绿素含量,并减少植株枝条、叶片、花蕾数量及叶片厚度[4-7],却使得植株高度和单株叶面积有所增加。弱光条件下,辣椒的净光合速率、蒸腾速率、羧化效率、光补偿点和 CO2补偿点均显著降低,而表观量子效率升高[8]。此外,寡照胁迫严重时可造成黄瓜植株叶绿体发育不良,排列紊乱,超微结构遭到破坏,叶绿体数量减少,叶绿素的降解加剧,叶绿素含量降低[9],最终造成作物产量降低[10]。近年来,国内外关于寡照对黄瓜[11]、番茄[12]、辣椒[13]、甜椒[14]和茄子[15]等主栽设施蔬菜生长发育影响的研究较多,但大多是于人工气候箱条件下,研究单因素寡照对设施作物苗期的光合生理特性及形态的影响;而北方日光温室实际生产条件下,寡照对黄瓜苗期及花果期光合特性影响的研究却少见报道。
文章拟以日光温室黄瓜品种“津优35”为研究对象,在日光温室条件下设计寡照处理试验,充分模拟生产条件下设施黄瓜在苗期及花果期分别遭遇不同等级寡照气象灾害后光合特性的变化情况,以期为日光温室黄瓜生产中寡照气象灾害的等级确定及科学防御提供依据。
1 材料与方法
1.1试验设计
试验于2012年2—4月在章丘伟丽种苗基地进行,供试日光温室坐北朝南,单坡式结构,东西长45m、南边宽9m,温室顶部覆盖白色聚乙烯薄膜(0.08mm),夜间覆盖草帘;供试黄瓜品种为“津优35”,待幼苗生长至第4片真叶时定植,大行距55cm,小行距35cm,每垄36株,缓苗一周后进行寡照处理,寡照处理通过遮阳网遮蔽实现,将遮阳网内光合有效辐射控制在 100~200μmol·CO2·m-2·s-1,与多云、阴天时日光温室内光合有效辐射基本一致。
在温室中均匀选取四垄长势良好的黄瓜幼苗,三垄做寡照处理,一垄做对照(CK)。试验开始后,将做寡照处理的黄瓜幼苗用遮阳网遮蔽,分别于寡照处理第1,3,5,7,10,15d由北向南揭开遮阳网,每次露出6株幼苗,分别标记为S1,S3,S5,S7,S10,S15;CK与寡照处理相对应也分为6组,以减少日光温室内南北向温、湿度及光线差异的影响。试验期间各处理水分和养分均保持在适宜水平。
待黄瓜生长至花果期,另选取4垄长势良好的花果期黄瓜作为供试样本,进行如上寡照及恢复处理。
1.2光合参数的测定
采用便携式光合作用测定系统(Li-6400,USA)于上午9:00—11:00测定各处理黄瓜叶片的净光合速率(Pn)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、气孔导度(Cond)、叶片温度(Tleaf)、光合有效辐射(PAR)、空气相对湿度(RH)等光合参数;控制叶室中CO2浓度为375μmol/mol,叶室温度为25℃。气孔限制值由公式(1)计算:
Ls=1-(Ci/Co)…(1) Co表示样本室中CO2浓度(μmol/mol),Ls表示气孔限制值。
1.3数据分析
采用Microsoft Excel 2003处理试验数据并制图,SAS 9.0进行方差分析,应用邓肯新复极级差法分析不同处理间的显著性差异。
2 结果与分析
2.1寡照对黄瓜叶片光合速率的影响
图1为不同寡照处理对苗期和花果期黄瓜叶片光合速率的影响情况。此研究在北方日光温室实际生产条件下进行,太阳辐射波动较大,为去除辐射因素对光合速率的影响,对CK与寡照处理的光合速率做差值,记为PRD-V,差值越大,寡照对黄瓜叶片光合速率的影响越大。由图1可知,随着寡照天数的增加,苗期及花果期黄瓜叶片PRD-V均呈上升趋势,寡照处理1~7d,苗期及花果期黄瓜叶片的 PRD-V迅速增大,而寡照处理7d后,苗期及花果期黄瓜叶片的PRD-V则缓慢增大,并趋于稳定,表明寡照处理对黄瓜叶片光合速率影响显著,而寡照 7d可视为黄瓜叶片光合速率遭受胁迫的临界指标。苗期黄瓜叶片的 PRD-V显著大于花果期,黄瓜在苗期时光合速率受寡照影响更大,而花果期黄瓜对寡照的抗逆性更强。
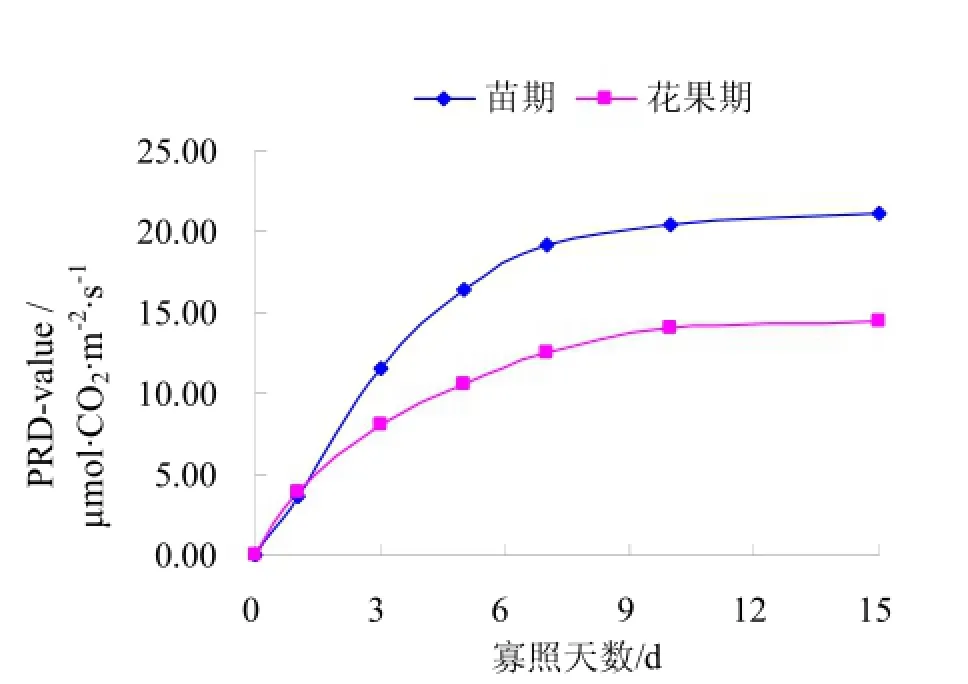
图1 寡照对黄瓜叶片光合速率的影响
2.2寡照对黄瓜叶片气孔限制值的影响
图2为不同寡照处理对苗期及花果期黄瓜叶片气孔限制值(Ls)的影响。对CK与寡照处理的气孔限制值做差值,记为LsD-V,差值越大,寡照对黄瓜叶片Ls的影响越大。由图2可知,随寡照处理天数的增加,苗期及花果期黄瓜叶片LsD-V均呈上升趋势,寡照处理 1~7d,LsD-V缓慢增长,而寡照处理7d后LsD-V增长速率显著增加,花果期黄瓜叶片的 LsD-V显著大于苗期,寡照处理7d后对黄瓜叶片气孔限制值影响更大,花果期气孔限制值受寡照影响更为显著。
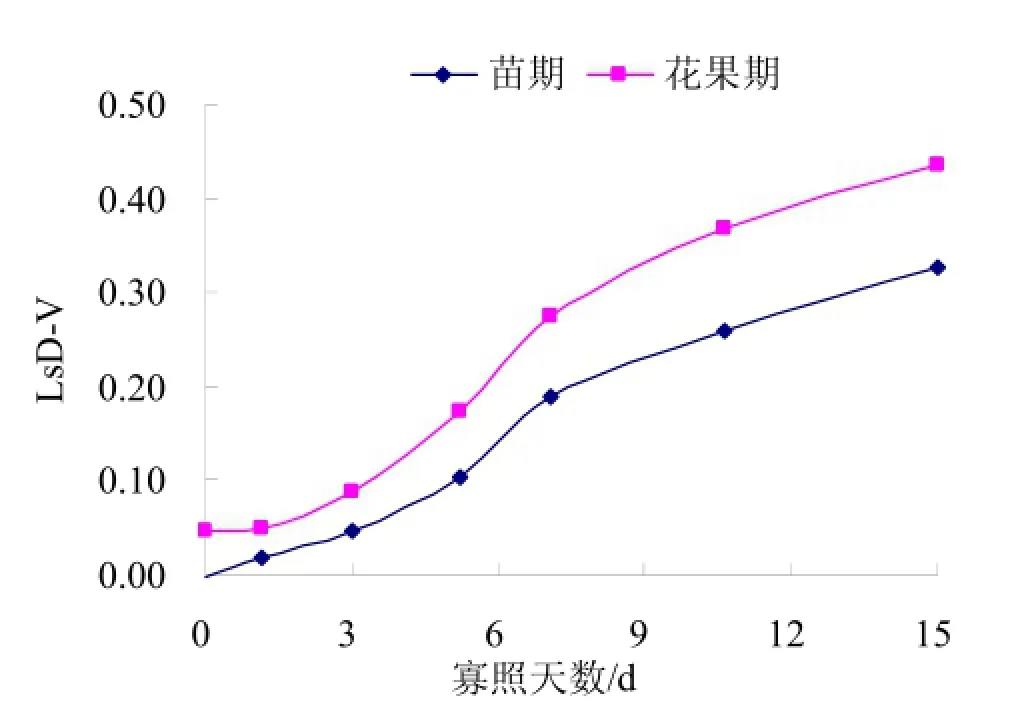
图2 寡照对黄瓜叶片气孔限制值的影响
2.3寡照对黄瓜叶片蒸腾速率和气孔导度的影响
图3a和图3b分别为不同寡照处理对苗期及花果期黄瓜叶片蒸腾速率(Tr)和气孔导度(Cond)的影响。对CK与寡照处理的蒸腾速率和气孔导度做差值,记为TrD-V和CD-V,差值越大,寡照对黄瓜叶片Tr和Cond的影响越大。由图3a和图3b可知,随寡照天数的增加,苗期及花果期黄瓜叶片 TrD-V和 CD-V均呈上升趋势,寡照处理1~7d,苗期及花果期黄瓜叶片的TrD-V和CD-V迅速增大,而寡照7d后,苗期及花果期黄瓜叶片的TrD-V和CD-V则缓慢增大,并趋于稳定;苗期黄瓜叶片TrD-V和CD-V均显著大于花果期,表明苗期黄瓜叶片的 Cond 和Tr受寡照影响更大。
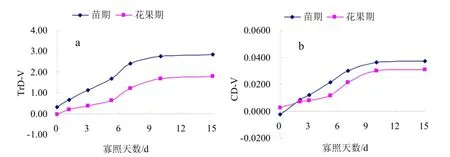
图3 寡照对黄瓜叶片蒸腾速率(a)及气孔导度(b)的影响
3 讨论与结论
此研究在北方日光温室实际生产条件下设计苗期及花果期黄瓜寡照处理试验,研究寡照对黄瓜叶片光合特性的影响。光合作用是作物生物产量的生理基础,作物对环境胁迫响应的共同特征主要体现在叶片光合速率的降低。试验时段太阳辐射变化幅度较大,不能将自然光条件下获取的作物光合速率直接进行比较。为去除辐射波动对作物光合速率产生的影响,对CK与寡照处理的光合速率、气孔限制值、蒸腾速率和气孔导度分别做差值,记为PRD-V,LsD-V,TrD-V和CD-V,差值越大,寡照对黄瓜叶片光合速率、气孔限制值、蒸腾速率和气孔导度的影响越大。寡照处理1~7d,苗期及花果期黄瓜叶片的PRD-V,TrD-V和CD-V均迅速增大,寡照处理7d后,苗期及花果期黄瓜叶片的PRD-V,TrD-V和CD-V缓慢增大,并趋于稳定;而LsD-V则在寡照处理1~7d的增长速率显著低于寡照处理7d后,因此,寡照7d可以视为黄瓜光合特性产生胁迫的临界值。除LsD-V外,苗期黄瓜叶片的PRD-V,TrD-V和CD-V均显著大于花果期,表明苗期黄瓜光合特性更易遭受寡照胁迫影响。Gunderson et al.(1994)研究认为黄瓜最大光合速率随寡照胁迫时间增加逐渐降低,是作物为适应弱光环境,逐渐调整自身功能,从而逐步丧失在强光下具有较强光合作用的潜能,即出现光合适应现象[16]。
文章系统研究了日光温室适温条件下不同程度寡照处理对苗期及花果期黄瓜光合特性的影响,但温室实际生产条件下存在着黄瓜植株生长周期较长,破坏性试验成本较高,连作障碍等问题,无法获取连续多年大量观测资料,在今后的研究中应进一步加强观测进行佐证。
[1] Yamazaki J Y. Is light quality involved in the regulation of the photosynthetic apparatus in attached rice leaves[J]. Photosynthesis Research,2010,105:63-71.
[2] Haliapas S,Yupsanis T A,Syros T D,et al. Petunia hybrida during transition to flowering as affected by light intensity and quality treatments[J]. Acta Physiol Plant,2008,30:807-815.
[3] Tanaka R,Tanaka A. Effects of chlorophyllide a oxygenase overexpression on light acclimation in Arabidopsis thaliana[J]. Photosynthesis Research,2005,85: 327-340.
[4] Barisic N,Stojkovic B,Tarasjev A. Plastic responses to light intensity and planting density in three Lamium species[J]. Plant Systematics and Evolution,2006,262:25-36.
[5] Seidlova L,Verlinden M,Gloser J,et al. Which plant traits promote growth in the low-light regimes of vegetation gaps[J]. Plant Ecology,2009,200:303-318.
[6] Shen W Q,Zhang G P,Xi L F,et al. Effects of light and temperature conditions on flowering in Mat Rush[J]. Journal of Zhejiang University Science,2002,3(5):611-616.
[7] Hou J L,Li W D,Zheng Q Y,et al. Effect of low light intensity on growth and accumulation of secondary metabolites in roots of Glycyrrhiza uralensis Fisch[J]. Biochemical Systematics and Ecology,2010,38:160-168.
[8] 眭晓蕾,张宝玺,张振贤,等. 弱光条件下不同基因型辣椒幼苗光合与生长的差异[J].农业工程学报,2005,21(增刊):41-44.
[9] 沈文云,马德华,侯锋,等. 弱光处理对黄瓜叶绿体超微结构的影响[J]. 园艺学报,1995,22(4):397-398.
[10] Baltzer J L,Thomas S C. Physiological and morphological correlates of whole- plant light compensation point in temperate deciduous tree seedlings[J]. Oecologia,2007,153:209-223.
[11] 艾希珍,郭延奎,马兴庄,等. 弱光条件下日光温室黄瓜需光特性及叶绿体超微结构[J].中国农业科学,2004,37(2):268-273.
[12] 黄伟,任华中,张福墁. 低温弱光对番茄苗期生长和光合作用的影响[J]. 中国蔬菜,2002,(4):15-17.
[13] 张志刚,尚庆茂. 低温、弱光及盐胁迫下辣椒叶片的光合特性[J]. 中国农业科学,2010,43(1):123-131.
[14] Jan Pilarski,Maciej Kocurek. The content of photosynthetic pigments and the light conditions in the fruits and leaves of sweet pepper[J]. Acta Physiologiae Plantarum,2005,2(27):173-182.
[15] 郁继华,舒英杰,吕军芬,等. 低温弱光对茄子幼苗光合特性的影响[J]. 西北植物学报,2004,24(5):831-836.
[16] Gunderson C A,Wullschleger S D. Photosynthetic acclimation in trees to rising atmospheric CO2:A broader perspective[J]. Photosynthesis Research,1994,39:369-388.
S16
B
1005-0582(2015)03-0026-04
2014-08-12
山东省2013年度农业重大应用技术创新项目“新型双屋面日光温室及高效种植模式研究”和山东省气候中心创新项目(2015qhzx04)共同资助
张继波(1987—),男,汉族,山东青州人,助工,主要从事设施环境调控方面的研究。
猜你喜欢
农业知识(2022年9期)2022-10-13
美食(2022年5期)2022-05-07
农业工程技术(2022年1期)2022-04-19
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
中国农业科技导报(2020年12期)2020-03-15
河北果树(2020年1期)2020-02-09
现代园艺(2018年2期)2018-03-15
现代园艺(2018年3期)2018-02-10
创新作文(小学版)(2018年6期)2018-01-26
