
不同品系条斑紫菜采收期游离氨基酸组成与含量变化特征
2015-08-20 12:08胡传明陆勤勤张美如朱建一马飞周伟沈辉
江苏农业科学 2015年7期
关键词:采收期
胡传明 陆勤勤 张美如 朱建一 马飞 周伟 沈辉

摘要:以7个条斑紫菜栽培品系为材料,利用柱前衍生高效液相色谱法,对其在采收期前期、中期与后期的16种游离氨基酸含量进行测定。结果显示,试样游离氨基酸总含量在48.71~62.05 mg/g之间,其中,Ala、Arg、Glu与Asp 4种呈味氨基酸含量最高,总含量在36.05~53.96 mg/g之间,占总游离氨基酸的69.11%~86.47%,除对照品系外,其他6个品系的呈味氨基酸与游离氨基酸总量均在采收中期达到最高;所有品系的必需氨基酸含量均随采收时间延迟呈增长趋势,在采收后期达到最高;苏通条斑紫菜品系在呈味氨基酸与总游离氨基酸含量组成水平上最高,其次为Y-H002、苏研与苏连,对照品系最低。
关键词:条斑紫菜;游离氨基酸;采收期;栽培品系
中图分类号: S917.3 文献标志码: A 文章编号:1002-1302(2015)07-0334-04
紫菜属(Pyropi)是一类原始大型红藻,生长分布范围广泛,其经济种类条斑紫菜和坛紫菜是我国重要的栽培海藻。条斑紫菜是属于原红藻纲、红毛菜科的一类海洋大型藻类,其生活史由宏观的叶状体(配子体)阶段和微观的丝状体(孢子体)阶段循环完成[1-2]。1953年,Kurogi开始在实验室内培养丝状体获得壳孢子,并由壳孢子得到幼叶状体[3-4]。20世纪50年代末,国外学者陆续提出紫菜的“种”与栽培性状的概念,由此建立了紫菜人工栽培技术,条斑紫菜选育研究也得到广泛发展。1969年,Miura经过多年选育,获得具有稳定遗传性状的栽培品种——奈良轮条斑紫菜(P. yezoensis f. narawaensis)。其后,通过单倍体育种方法成功培育出色泽优、产量高的条斑紫菜新种质晓光,成为20世纪90年代日本盛行的条斑紫菜栽培品种[5]。我国藻类学者在20世纪80年代初,基本延袭这一方法选育成1个长型条斑紫菜品种[2]。目前,单倍体育种技术有了长足发展,并通过种内、种间杂交、定向培育、诱变育种等技术手段,开展了大量的条斑紫菜种质改良、新品系研发等。
条斑紫菜属于一类风味食品,除对产量与形态等基本特征进行必要描述[6]、对其叶绿素荧光特征、脂肪酸与挥发物的组成含量等开展研究外[7-11],评价一个栽培品系的风味特征是工作重点之一,可以反映其食用品质与商业价值。风味物包括游离氨基酸、5′-核苷酸、还原糖与有机酸等一系列物质[12]。条斑紫菜游离氨基酸以Glu、Ala、Asp、Arg、Tau等呈味氨基酸含量最高,以体现其醇厚的风味[13]。游离氨基酸成分与含量不仅与条斑紫菜的种质有关[13],而且与其栽培环境及采收时间也有一定的联系[13-16]。本研究以包括杂交品系、高光强胁迫筛选品系、诱变选育品系、自然选育品系与栽培品系在内的7个条斑紫菜品系为材料,测定其在采收期前期、中期与后期的游离氨基酸含量,分析各品系游离氨基酸组成与含量变化特征,为条斑紫菜各栽培品系的评价研究提供参考。
1 材料与方法
1.1 供试材料
在江苏南通紫菜栽培区,分别于2013年1月1日(前期)、2月28日(中期)、3月28日(后期)采集各条斑紫菜品系(表1);使用海水清洗藻体,并剔除杂藻和藻体基部,快速过纯水去除盐分;藻体低温快速风干,-20 ℃保存。
1.2 游离氨基酸含量的测定
样品研磨,过100目筛;精确称取藻粉0.30 g,加入体积分数75%的乙醇50 mL,100 ℃水浴15 min,40 ℃真空旋转蒸干, 重复上述步骤1次;用适量pH值为2.2的柠檬酸缓冲液
对照条斑紫菜主产区良种栽培品系江苏省海洋水产研究所
溶解,3 000 r/min 离心5 min;取上清,定容至50 mL,过滤备用;采用Agilent HPLC 1200型自动进样器对氨基酸标准品和样品进行OPA-FMOC自动在线衍生,样品经OPA一级衍生与FMOC二级衍生后上氨基酸专用HPLC色谱柱,根据 Agilent 氨基酸测定手册进行测定,面积归一法进行定量。标准品采用9、90、900 pmol 3个浓度。
1.3 数据分析
采用Excel与SPSS 19.0软件对相关数据进行统计分析。
2 结果与分析
2.1 条斑紫菜游离氨基酸的组成
检测7个条斑紫菜品系采收期前、中、后期共21个试样,均检出16种游离氨基酸,包括Thr、Tyr、Val、Met、Phe、Ile和Leu 7种必需氨基酸,His和Arg 2种半必需氨基酸和Ala、Asp、Glu、Ser、Gly、Cy2、Pro 7种非必需氨基酸(图1)。
由表2可见,21个试样中,游离氨基酸总含量在48.71~62.05 mg/g之间;前期的总游离氨基酸以Y-H002最高,含量为56.87 mg/g,以Y-008最低,含量为50.83 mg/g;中期以苏连最高,含量为62.05 mg/g,以对照最低,含量为 53.69 mg/g;后期以Y-H002最高,含量为58.86 mg/g,以对照最低,含量为48.71 mg/g。采收前、中、后期游离氨基酸总含量的平均值,苏通最高,Y-H002次之,对照最低。
16种游离氨基酸中,以Ala、Arg、Glu与Asp 4种呈味氨基酸含量较高,总含量在36.05~53.96 mg/g之间,占总游离氨基酸的69.11%~86.47%,其中,Ala占总游离氨基酸含量的23.4%~32.79%,Arg占23.51%~32.65%,Glu占951%~18.94%,Asp占6.68%~10.05%。7个品系4种呈味氨基酸总含量中,前期以对照最高,总含量为45.51 mg/g,其次为苏通,总含量为45.05 mg/g,Y-H002最低,总含量为41.70 mg/g;中期以苏通最高,总含量為53.96 mg/g,其次为Y-H002,总含量为51.81 mg/g,对照最低,总含量为42.97 mg/g;后期以苏通最高,总含量43.60 mg/g,其次为Y-2011,总含量为43.84 mg/g,对照最低,总含量为36.05 mg/g。
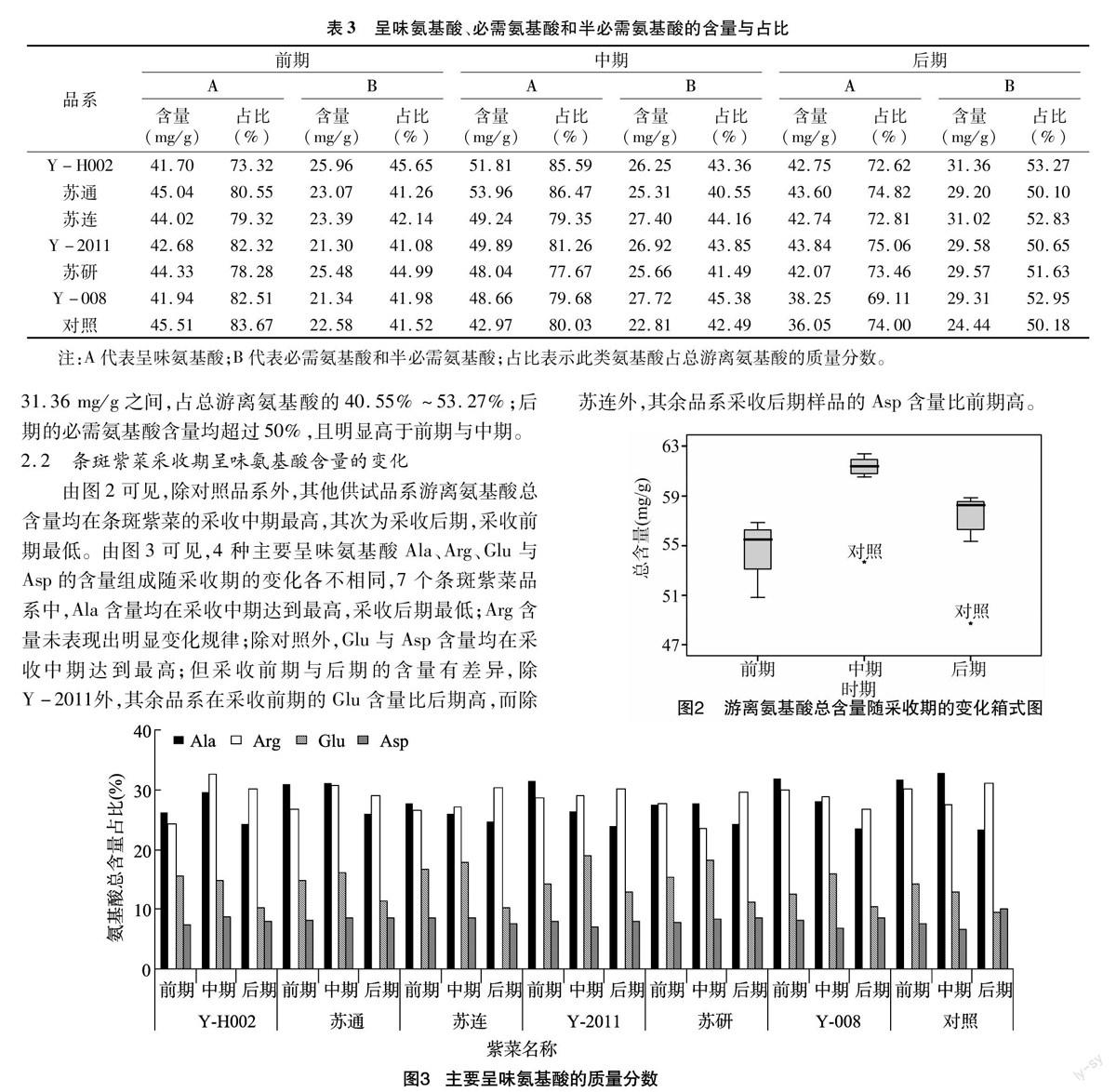
由表3可见,16种游离氨基酸中必需氨基酸含量丰富,包括7种必需氨基酸和2种半必需氨基酸,含量在21.3~
表3 呈味氨基酸、必需氨基酸和半必需氨基酸的含量与占比
品系前期中期后期
ABABABABABAB
含量(mg/g)占比(%)含量(mg/g)占比(%)含量(mg/g)占比(%)含量(mg/g)占比(%)含量(mg/g)占比(%)含量(mg/g)占比(%)
Y-H00241.7073.3225.9645.6551.8185.5926.2543.3642.7572.6231.3653.27
苏通45.0480.5523.0741.2653.9686.4725.3140.5543.6074.8229.2050.10
苏连44.0279.3223.3942.1449.2479.3527.4044.1642.7472.8131.0252.83
Y-201142.6882.3221.3041.0849.8981.2626.9243.8543.8475.0629.5850.65
苏研44.3378.2825.4844.9948.0477.6725.6641.4942.0773.4629.5751.63
Y-00841.9482.5121.3441.9848.6679.6827.7245.3838.2569.1129.3152.95
对照45.5183.6722.5841.5242.9780.0322.8142.4936.0574.0024.4450.18
注:A代表呈味氨基酸;B代表必需氨基酸和半必需氨基酸;占比表示此类氨基酸占总游离氨基酸的质量分数。
31.36 mg/g之间,占总游离氨基酸的40.55%~53.27%;后期的必需氨基酸含量均超过50%,且明显高于前期与中期。
2.2 条斑紫菜采收期呈味氨基酸含量的变化
由图2可见,除对照品系外,其他供试品系游离氨基酸总含量均在条斑紫菜的采收中期最高,其次为采收后期,采收前期最低。由图3可见,4种主要呈味氨基酸Ala、Arg、Glu与Asp的含量组成随采收期的变化各不相同,7个条斑紫菜品系中,Ala含量均在采收中期达到最高,采收后期最低;Arg含量未表现出明显变化规律;除对照外,Glu与Asp含量均在采收中期达到最高;但采收前期与后期的含量有差异,除Y-2011外,其余品系在采收前期的Glu含量比后期高,而除苏连外,其余品系采收后期样品的Asp含量比前期高。
3 结论与讨论
3.1 条斑紫菜游离氨基酸含量与组成特征
海区间水温、营养盐水平的差异会对条斑紫菜的生长产生不同的影响[14-15],再加上种质与采收期各不相同,条斑紫菜游离氨基酸总量差异较大,如日本报道紫菜中游离氨基酸总量在25~50 mg/g之间[15],而国内有报道在20~50 mg/g之间[17-18]。本试验7个品系的条斑紫菜试样均采自海门辐射沙洲同一个海区,避免了因海区不同造成的游离氨基酸含量差异,测得的游离氨基酸总量在48.71~62.05 mg/g之间,与李瑞霞等的试验结论[17]差别较大,可能与样品的处理方法有关。
Glu、Asp、Arg、Ala是条斑紫菜重要的呈味氨基酸[17],其中,Ala与Glu是影响紫菜风味的重要氨基酸[12,19]。试验结果表明,这4种氨基酸在条斑紫菜中含量最高,总含量在3605~53.96 mg/g之间,占总游离氨基酸的69.11%~8647%,符合条斑紫菜强烈的口感特征。Yoshie等對8种日本产条斑紫菜干制品中的游离氨基酸含量进行分析,结果表明,Ala、Glu、Asp含量分别在3.08~16.02、3.22~18.44、066~2.82 mg/g之间[20]。纪明侯等对青岛产条斑紫菜中游离氨基酸进行检测,结果显示,高肥区Ala、Glu、Asp含量分别在7.3~30.2、2.9~9.3、0.5~2.7 mg/g[14]。本试验结果表明,7个品系中,Ala、Glu、Asp含量分别在11.40~19.42、4.63~1163、3.59~5.28 mg/g之间,Asp含量明显高于Yoshie等和纪明侯等的测定结果,Ala与Glu含量与Yoshie等和纪明侯等的测定结果相当,最低值均高于Yoshie等和纪明侯等的测定结果最低水平。
游离氨基酸总含量在采收前期以Y-H002与苏研最高,中期以苏通与苏连最高,后期以Y-H002与苏连最高;每个品系氨基酸总含量的平均值以苏通与Y-H002最高,对照品系最低。4种主要呈味氨基酸含量,前期为对照品系与苏通最高,中期以苏通与Y-H002最高,后期以Y-2011与苏通最高。7个条斑紫菜品系中,苏通在呈味氨基酸与总游离氨基酸含量组成水平上最高,其次为Y-H002、苏研与苏连,对照品系最低。
3.2 游离氨基酸随采收期的变化特征
条斑紫菜的壳孢子采苗、单孢子放散和营养生长都与海区气温及水温有着密切关系,其采收期也随海区水温的差异有着明显的不同,不同条斑紫菜品系中的游离氨基酸含量组成变化也不一致。纪明侯等分析从1—4月采收的条斑紫菜认为,虽然部分游离氨基酸在采收期中段出现1个高峰后下降,但是游离氨基酸总体随采收期延迟呈下降趋势[14]。Sakai等调查11月至翌年1月日本北海道产条斑紫菜中游离氨基酸含量认为,游离氨基酸随采收期延迟呈增长趋势,特别是Asp、Ser、Glu与Ala[19]。Niwa等分析2个条斑紫菜品系HG-4与HG-5采收次数对游离氨基酸的影响发现,Ala、Asp、Glu、Tau 4种游离氨基酸中,只有Glu的含量随采收期延迟而降低[13],本试验对照品系出现同样结论;其他3种氨基酸含量在HG-4纯系的第3次采收时达到最大,后出现下降,本试验从壳孢子采苗至采收前期,条斑紫菜在海中生长70多天,除对照外,其他6个品系中的Ala、Asp、Glu、Arg 4种主要呈味氨基酸含量从前期的41.70~45.51 mg/g上升到中期的42.97~53.96 mg/g,后期下降到36.05~43.84 mg/g,同样出现1个高峰后下降。
与呈味氨基酸含量相反,7个条斑紫菜品系中必需氨基酸含量均随采收时间延迟呈现增长趋势,而必需氨基酸含量的多寡决定其营养的平衡程度与利用价值,呈味氨基酸与必需氨基酸含量的变化为条斑紫菜游离氨基酸评价提供了一个平衡点。
参考文献:
[1]曾呈奎,张德瑞. 紫菜的研究——Ⅲ.紫菜的有性生殖[J]. 植物学报:英文版,1955,4(2):153-166.
[2]张学成,秦 松,马家海,等. 海藻遗传学[M]. 北京:中国农业出版社,2005:184-317.
[3]Kurogi M. Study of the life-history of Porphyra Ⅰ. The germination and development of carpospores[J]. Bull Tohoku Reg Fish Lab,1953,2:67-103.
[4]Kurogi M. On the liberation of monospores from the filamentous thallus(conchocelis-stage)of Porphyra tenera Kjellm[J]. Bull Tohoku Reg Fish Lab,1953,2:104-108.
[5]Miura A. A new variety and a new form of Porphyra(Bangiales,Rhodophyta) from Japan:Porphyra tenera Kjellman var. tamatsuensis Miura,var. nov. and P. yezoensis Ueda form.narawaensis Miura,form.nov.[J]. Journal of the Tokyo University Fisheries,1950,71(1):1-37.
[6]张美如,陆勤勤,陈淑吟,等. 条斑紫菜品系评价方法的探讨[J]. 水产科技情报,2011,38(6):273-280,283.
[7]张 涛,沈宗根,李家富,等. 紫菜不同品系贝壳丝状体叶绿素荧光特性比较[J]. 江苏农业科学,2012,40(11):238-242.
[8]李家富,张 涛,陆勤勤,等. 坛紫菜叶状体营养细胞与生殖细胞叶绿素荧光特性比较[J]. 海洋科学,2013,37(3):82-86.
[9]姚春燕,张 涛,姜红霞,等. 条斑紫菜不同品系藻体光合色素及叶绿素荧光参数比较[J]. 南京师大学报:自然科学版,2010,33(2):81-86.
[10]马 飞,陆勤勤,胡传明,等. 多个条斑紫菜品系采收期内脂肪酸组成、含量的变化分析[J]. 水产学报,2013,37(10):1551-1557.
[11]胡传明,徐继林,朱建一,等. 紫菜特征挥发性物质分析[J]. 海洋科学,2011,35(5):106-111.
[12]Noda H,Horiguchi Y,Araki S. Studies on the flavor substances of‘Nori,the dried laver Porphyra spp.-Ⅱ.Free amino acids and 5′-nucleotides[J]. Nippon Suisan Gakkaishi,1975,41(12):1299-1303.
[13]Niwa K,Furuita H,Yamamoto T. Changes of growth characteristics and free amino acid content of cultivated Porphyra yezoensis Ueda (bangiales rhodophyta) blades with the progression of the number of harvests in a nori farm[J]. Journal of Applied Phycology,2008,20(5):687-693.
[14]紀明侯,蒲淑珠,牛伫庆.不同海区生长的条斑紫菜的氨基酸含量变化[J]. 海洋与湖沼,1981,12(6):522-530.
[15]马家海,大住幸宽,川合正允. 浙江省象山港紫菜轮栽及其品质分析的研究[J]. 中国水产科学,1997,4(1):30-37.
[16]李信书,伏光辉,陈百尧,等. 氮、磷加富对条斑紫菜生长及生化组成的影响[J]. 水产科学,2012,31(9):544-548.
[17]李瑞霞,伊纪峰,沈颂东,等. 条斑紫菜(Porphyra yezoensis)游离氨基酸组成分析[J]. 氨基酸和生物资源,2011,33(1):4-9.
[18]胡传明.紫菜产品品质及安全质量分析研究[D]. 南京:南京师范大学,2008:24-34.
[19]Sakai H,Kasai T. Fatty acids,free amino acids and 5′-nucleotides of dried laver,Hoshi-nori,harvested in different months in Hokkaido and produced under different drying conditions[J]. Nippon Shokuhin Kagaku Kogaku Kaishi,2000,47(4):327-332.
[20]Yoshie Y,Suzuki T,Shirai T,et al. Free amino acids and fatty acid composition in dried nori of various culture locations and prices[J]. Nippon Suisan Gakkaishi,1993,59:1769-1775.
猜你喜欢
中国果树(2023年9期)2023-11-27
中国果树(2023年5期)2023-07-17
中成药(2021年5期)2021-07-21
河北果树(2020年4期)2020-11-26
亚热带植物科学(2020年4期)2020-09-28
食品与发酵工业(2019年13期)2019-07-24
天然产物研究与开发(2018年8期)2018-09-10
中成药(2017年5期)2017-06-13
中成药(2017年4期)2017-05-17
特产研究(2015年1期)2015-04-12
