
二氧化碳气调对锯谷盗的毒力及其能源物质利用的影响
2015-08-02 03:59:22曹宇闫玉芳杨文佳熊正利王丽娟李灿
浙江大学学报(农业与生命科学版) 2015年6期
曹宇,闫玉芳,杨文佳,熊正利,王丽娟,李灿*
(1.贵阳学院生物与环境工程学院,有害生物控制与资源利用贵州省高校特色重点实验室,贵阳550005;2.重庆市渝东南农业科学院,重庆涪陵408000)
二氧化碳气调对锯谷盗的毒力及其能源物质利用的影响
曹宇1,闫玉芳2,杨文佳1,熊正利1,王丽娟1,李灿1*
(1.贵阳学院生物与环境工程学院,有害生物控制与资源利用贵州省高校特色重点实验室,贵阳550005;2.重庆市渝东南农业科学院,重庆涪陵408000)
为探讨二氧化碳(CO2)气调对锯谷盗(Oryzaephilus surinamensis Linne)的控制作用,研究了不同CO2浓度对锯谷盗的毒力效果,以及CO2气调胁迫与致死作用对锯谷盗能源物质含量和利用的影响,以期从生理生化角度探讨CO2气调的作用机制.试验在温度(25±1)℃、相对湿度(70±5)%的条件下进行,CO2气调设为10%、30%、50%、70%和90%5个梯度,每隔3h观察其对锯谷盗的致死情况;另外,利用各浓度CO2胁迫2、4、6和8 h及致死后,测定锯谷盗体内多糖、可溶性蛋白质及脂肪含量,计算其利用率.结果表明:不同CO2气调浓度处理对锯谷盗具有不同的毒力作用,随着其浓度的升高,毒力逐渐增强,在10%CO2处理下,50%锯谷盗致死时间和99%锯谷盗致死时间分别为52.23和131.45h,在90%CO2处理下分别为12.52和32.44h;在气调胁迫下,同浓度CO2处理随着时间的延长以及胁迫相同时间随着CO2浓度的升高,锯谷盗体内多糖、可溶性蛋白质及脂肪含量均降低,利用率逐渐提高,其对3种能源物质的利用率为可溶性蛋白质≈脂肪>多糖.在气调致死下,不同能源物质剩余量均随着气调浓度的升高逐渐增多,但均低于其气调胁迫不同时间后的含量;同时,锯谷盗对3种能源物质的利用率逐渐降低,但均高于气调胁迫不同时间后的利用率,其对三者的利用率为多糖>可溶性蛋白质>脂肪.因此,不同CO2气调浓度对锯谷盗不同的毒力可能与昆虫对能源物质的不同响应有关,昆虫对能源物质利用的显著差异可能与其抗气性的形成有关.
锯谷盗;二氧化碳气调;能源物质;多糖;可溶性蛋白质;脂肪
Journal of Zhejiang University(Agric.&Life Sci.),2015,41(6):631-640
SummaryThe application of controlled atmosphere(CA),is an effective,safe,and residue-free alternative compared with chemical fumigants and protectants against pests and mites,and using of carbon dioxide(CO2)hasreceived considerable attention in disinfesting stored foodstuffs,particularly durable products.There are reports about the toxic effects of CO2-enriched atmospheres on several insect pests of stored products,but the data on the toxicity of CO2against Oryzaephilus surinamensis Linne are limited.Oryzaephilus surinamensis is one of the major pests of stored spices and spice products found in many districts and countries,causing huge losses to stored products especially economical crops and food products.Oryzaephilus surinamensis management in stored food commodities is facing a challenge resulting from pesticide residues in foodstuffs and the increasing resistance of insects to conventional chemical pesticides.It is essential to develop alternative pest control techniques for protecting stored commodities.Therefore,the toxicities of different CO2-enriched atmospheres to O.surinamensis were studied,and in order to explore the action mechanism of CO2controlled atmosphere at physiological and biochemical levels,the energy substance content and its utilization rate in O.surinamensis were also determined and calculated after stress of several hours under different CO2concentrations or until dead.
The experiments were conducted at the constant temperature of(25±1)℃and relative humidity of(70± 5)%.Oryzaephilus surinamensis were exposed under different CO2concentrations(10%,30%,50%,70%and 90%),and the mortality of every three hours were observed until all of the insects died.In addition,the contents of polysaccharide,soluble protein and lipid in O.surinamensis were determined after the CO2stress for 2,4,6,8 h or until dead under the different CO2concentrations,and the energy substance contents of O.surinamensis in normal atmosphere were determined to calculate the utilization rate of each energy substance.
The results showed that different concentrations of CO2had different toxicity on O.surinamensis,and the toxicity became stronger with higher CO2concentration.The lethal time of 50%(LT50)and 99%(LT99)O.surinamensis were 52.23and 131.45hunder 10%CO2,while they were 12.52and 32.44hunder 90%CO2,respectively.The contents of polysaccharide,soluble protein and lipid decreased,but the utilization rate increased,with increasing CO2concentration under the same stress time or with the extension of stress time under the same CO2concentration,and the consumptions of the three kinds of energy substances were soluble protein≈lipid>polysaccharide in all the stress treatments.The residual contents of energy substance in O.surinamensis performed a gradual increase with the increasing CO2concentration after the CO2stress until dead,but decreased in its utilization rate.And the O.surinamensishad less residual content of energy substances under the CO2long stress until dead than that under the CO2short stress for 2,4,6and 8h,but it had higher utilization rate with the consumption of polysaccharide>soluble protein>lipid.
Comparison of the utilization rates showed that soluble protein and lipid were the main energy substances,followed by polysaccharide,to deal with the short CO2stress time;but polysaccharide was the prior energy substance of O.surinamensis to respond to the long stress period until dead,followed by soluble protein,and lipid was the last.So the different toxicity of different CO2concentrations to O.surinamensis may have some relationship between different responses of insects to the energy substances and the CO2stress,revealing that the significantly different utilization rates of energy substances in insect probably have some associations with its resistance development to CO2.
锯谷盗(Oryzaephilus surinamensis Linne)隶属于鞘翅目(Coleoptera),锯谷盗科(Silvanidae),是一种世界性储藏物害虫,可对粮食、药材、烟草、干果等许多储藏物造成不同程度的危害[1-2].对于锯谷盗的生物学、生态学特性已有许多报道:研究发现在不同的温、湿度下,锯谷盗在麦麸上的生长发育历期不同[3],对不同温度储存下的面粉为害不同[4],在不同处理面粉上的种群动态变化也不同[5];锯谷盗对红、黄、绿光等具有不同的趋性,且环境温度对其运动轨迹具有重要影响[6-7],因此,在不同环境下锯谷盗的分布差异显著,发生程度不同.锯谷盗危害范围广泛,尤其对经济作物和粮食等储藏物可造成巨大的经济损失.对其防治主要包括化学药剂[8-9]与生物源药剂控制[10-12]、微波辐射[13-14]、电子束辐射[15-16]和甲酸乙酯熏蒸等[17].且长期以来,对锯谷盗的防治主要依赖磷化氢和溴甲烷等熏蒸防治[18],而锯谷盗对磷化氢极易产生抗性,溴甲烷可破坏臭氧层,两者在我国将于2015年底被淘汰使用.另外,在害虫防治中,某些储藏物涉及食品安全问题,若任意使用化学药剂将存在安全隐患.因此,探索绿色、安全、环保的储藏物害虫防治措施具有重要的现实意义.
二氧化碳(CO2)气调控制是一种有效的储藏物害虫防治措施,无毒、无残留,具备良好的开发前景.CO2气调能够有效控制药材甲(Stegobium paniceum)、烟草甲(Lasioderma serricorne)、嗜卷书虱(Liposcelis bostrychophila)等仓储害虫的为害[19-21],因其安全环保,引起了各国学者的关注.唐培安等[22]利用氮气与甲酸乙酯联合来防治锯谷盗,但尚未见CO2气调对锯谷盗控制的研究报道.因此,本文研究了不同CO2浓度对锯谷盗的毒力作用,测定了在短时气调胁迫及致死下,锯谷盗体内多糖、可溶性蛋白质及脂肪等能源物质的含量变化及其利用率,为从生理生化角度探讨气调对锯谷盗的控害机制及昆虫的抗气性机制提供理论依据,为进一步探讨锯谷盗的可持续有效控制奠定基础.
1 材料与方法
1.1 供试虫源
锯谷盗采自贵阳市各类储藏粮食上,在人工气候实验箱(25±1)℃、相对湿度(70±5)%、光照(昼夜比)10h∶14h条件下饲养,以m(面粉)∶m(酵母粉)=10∶1的混合物为食料.试验时已人工饲养5代以上,以羽化2~3d的成虫作为试虫.
1.2 主要试剂及仪器设备
CO2气体(纯度高于99%)购于贵阳当地气体供应站;蒽酮为天津市科密欧化学试剂有限公司产品;考马斯亮蓝G-250、牛血清白蛋白(albumin bovine serum,ABS)为美国Sigma公司产品.
Allegra X-30高速冷冻离心机(美国Beckman Coulter公司);754PC紫外-可见分光光度计(上海菁华科技仪器有限公司);KL-RO-20自动双纯水系统(成都唐氏康宁科技发展有限公司);BSA124S电子天平(赛多利斯科学仪器北京有限公司).
1.3 试验方法
1.3.1锯谷盗的生物测定与气调处理
1.3.1.1CO2气调对锯谷盗的毒力测定.采用课题组前期气调处理方法[23],取锯谷盗成虫60头置于试验用培养盒中,放入适量面粉和酵母粉混合物,加盖密闭,通过气调混配仪,用导管通入设定浓度的气调气体,待气体浓度配比达到要求后,停止通气.将培养盒的进出气口导管对接,放入人工气候实验箱培养观察,设定试验相对湿度(70±5)%、温度(25±1)℃.CO2气调体积分数分别设置为10%、30%、50%、70%和90%.每隔3h观察和记录在不同气调浓度处理后试虫的存活情况,直至全部死亡,然后测定死亡试虫各能源物质的含量.昆虫死亡确定标准:将试虫移出处理环境5h后,以小毛笔刺激,不动视为死亡.以相同条件下的空气处理作为对照.试验重复3次,以Abbott公式校正死亡率[24].1.3.1.2CO2气调对锯谷盗的胁迫处理.采用
1.3.1.1节的气调处理方法,在相对湿度(70± 5)%、温度(25±1)℃条件下,分别在不同CO2气调浓度胁迫下处理2、4、6和8h后进行相关生化测定;以自然条件下的相关测定作为对照.每组处理50头昆虫,重复3次.
1.3.2CO2气调处理下锯谷盗能源物质测定
1.3.2.1多糖含量测定.参考程伟霞等[25]的蒽酮比色法进行测定.取CO2气调胁迫致死后的锯谷盗30头,充分匀浆后,于15 000r/min、25℃条件下离心10min,取其上清液加入0.2%蒽酮试剂,混合均匀后沸水浴10min,于620nm处测定其吸光度值.利用葡萄糖制作标准曲线,根据标准曲线将测定结果转换成μg/头.以同样条件下在空气中自然生长的活体锯谷盗多糖含量值作为对照.试验重复3次.多糖利用率/%=(对照含量-气调处理下含量)/对照×100[19].1.3.2.2可溶性蛋白质含量测定.采用程伟霞等[25]和Bradford[26]的考马斯亮蓝G-250法进行测定.取锯谷盗30头,加入pH 7.0、0.04mol/L磷酸缓冲液3mL于冰水浴中匀浆,匀浆液于10 000r/min、4℃离心15min,取其上清液加入1%考马斯亮蓝G-250反应,在595nm处测定其吸光度值.利用牛血清白蛋白制作标准曲线,根据标准曲线将测定结果转换成μg/头.以同样条件下在空气中自然生长的活体锯谷盗可溶性蛋白质含量作为对照.试验重复3次.可溶性蛋白质利用率/%=(对照含量-气调处理下含量)/对照×100.
1.3.2.3脂肪含量测定.参照Colinet等[27]的方法,将40头锯谷盗置于60℃恒温培养箱中干燥48 h,然后称量虫体干质量(m1).取干燥后的单头成虫于玻璃匀浆器中,加入V(氯仿)∶V(甲醇)=2∶1的提取液5mL,研磨匀浆,转移至离心管中以4 500 r/min离心.转移上清液,在沉淀中再加入6mL三氯甲烷和甲醇提取液,再次离心,将剩余沉淀在60℃恒温培养箱中干燥24h至恒质量(m2).脂肪含量=m1-m2.以自然条件下的锯谷盗脂肪含量作为对照.试验重复3次.脂肪利用率/%=(对照含量-气调处理下含量)/对照×100.
1.4 数据统计与分析
利用SPSS 18.0软件分析锯谷盗的死亡率和存活率,拟合在不同CO2浓度作用下的毒力回归方程;采用邓肯新复极差测验法比较在不同处理下锯谷盗多糖、可溶性蛋白质及脂肪含量的差异显著性.
2 结果与分析
2.1 不同CO2气调浓度处理对锯谷盗的毒力效果
在不同CO2气调浓度处理下,锯谷盗具有不同的敏感性(表1).随着气调浓度的升高,毒力回归曲线斜率增大,说明对锯谷盗的控制作用越强.在10%、30%、50%、70%和90%CO2作用下,50%锯谷盗死亡时间(LT50)分别为52.23、39.76、32.94、20.91和12.52h,99%锯谷盗死亡时间(LT99)分别为131.45、92.06、81.27、61.30和32.44h.随着气调浓度的升高,锯谷盗的LT50和LT99值逐渐减小,说明CO2气调浓度越高,对锯谷盗的毒力越强.
2.2 气调胁迫对锯谷盗能源物质的影响
2.2.1气调胁迫对锯谷盗多糖含量的影响
2.2.1.1气调胁迫下锯谷盗多糖含量的测定结果.在自然条件下,锯谷盗多糖含量为6.48μg/头.气调浓度和胁迫时间是影响其含量的重要因素(表2).在相同气调浓度下,随着胁迫时间的延长,其多糖含量逐渐减少,且浓度越高,减少越多.如经10%CO2胁迫8h后,由对照组的6.48μg/头显著减少为5.90 μg/头;经90%CO2胁迫8h后,减少为5.05μg/头;另外,在10%~90%CO2胁迫下,除90%CO2处理的多糖含量在6与8h之间差异无统计学意义外,其他各浓度在不同胁迫时间下差异均有统计学意义(P<0.05).同样,胁迫相同时间后,随着CO2浓度的升高,其多糖含量也呈现不同程度的显著下降.如胁迫2h后,在10%和90%CO2浓度下多糖含量分别为6.37和5.91μg/头;胁迫8h后,在这2个浓度下多糖含量分别为5.90和5.05μg/头.

表1 不同CO2气调浓度处理对锯谷盗的毒力测定结果Table 1 Toxicity result of different CO2concentrations to O.surinamensis

表2 CO2气调胁迫下锯谷盗的多糖含量Table 2 Polysaccharide content of O.surinamensis under the stress of different CO2concentrations μg/insect
2.2.1.2气调胁迫下锯谷盗对多糖的利用率.随着气调浓度的升高,胁迫时间的延长,锯谷盗对多糖的利用率显著提高(图1).在10%CO2浓度下,多糖利用率在胁迫2、4、6和8h之间差异有统计学意义(F=59.352,P<0.05),且随着胁迫时间的延长,多糖利用率逐渐提高.如在胁迫2h后的利用率为1.24%,8h后达到8.94%;其他浓度下的利用规律相似.在90%CO2的高浓度下,胁迫2h的利用率为9.23%,8h时显著上升为22.19%,但其与6h时差异无统计学意义.同样,在相同胁迫时间下,锯谷盗对多糖的利用率随着CO2浓度的升高而升高,且不同浓度之间差异显著.如在胁迫2h时,在10%、30%、50%、70%和90%各浓度下的多糖利用率分别为1.24%、3.38%、4.31%、6.34%和8.94%(F=62.428,P<0.05);在胁迫8h时,其利用率分别为9.23%、11.98%、15.87%、20.87%和22.19%(F=90.115,P<0.05).
2.2.2气调胁迫对锯谷盗可溶性蛋白质含量的影响
2.2.2.1气调胁迫下可溶性蛋白质含量的测定结果.在自然条件下,锯谷盗可溶性蛋白质含量为95.54μg/头.在各气调胁迫处理下,其含量随着CO2浓度的升高和胁迫时间的延长显著减少(表3),与多糖变化趋势类似.不同CO2浓度处理对蛋白质含量的影响有所不同:如胁迫2h后,可溶性蛋白质含量在10%与30%CO2处理之间差异无统计学意义;6h后,10%与30%、50%与70%CO2处理之间差异无统计学意义;8h后,50%、70%与90% CO2处理之间差异均无统计学意义;胁迫4h后,不同浓度之间的可溶性蛋白质含量差异有统计学意义.在同一浓度胁迫不同时间后,除90%CO2处理下的6与8h之间差异无统计学意义外,其余不同胁迫之间均在统计学上差异显著.
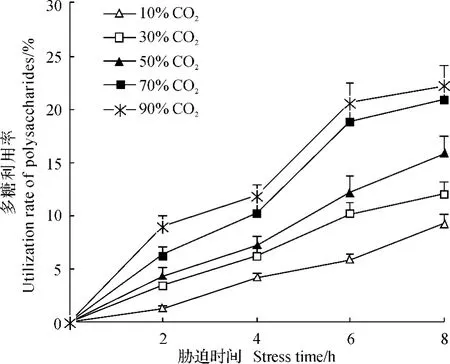
图1 在不同CO2浓度胁迫下锯谷盗对多糖的利用率Fig.1 Utilization rate of polysaccharides in O.surinamensis under the stress of different CO2concentrations

表3 CO2气调胁迫下锯谷盗的可溶性蛋白质含量Table 3 Soluble protein content of O.surinamensis under the stress of different CO2concentrations μg/insect
2.2.2.2气调胁迫下锯谷盗对可溶性蛋白质的利用率.相较于多糖利用率,锯谷盗对可溶性蛋白质的利用率曲线变化有所不同(图2),相似的是随CO2浓度的升高,胁迫时间的延长,利用率增大.10% CO2处理2h后,可溶性蛋白质利用率最低(3.39%),显著低于胁迫4、6和8h后的利用率(F=65.016,P<0.05).同样,在30%、50%、70%和90%CO2处理下,除90%CO2胁迫6与8h之间的利用率在统计学上无显著差异外,其他各浓度处理下的利用率均随时间延长显著升高.在胁迫相同时间下,锯谷盗对可溶性蛋白质的利用率随CO2浓度升高而升高,但不同浓度之间差异显著性有所不同.胁迫2h时,蛋白质利用率在10%与30% CO2处理之间差异无统计学意义,但显著低于50%、70%和90%CO2处理下的利用率,后三者之间差异有统计学意义(F=69.542,P<0.05);胁迫4h时,各浓度间在统计学上均差异显著(F=76.873,P<0.05);胁迫6h时,10%与30%、50%与70%CO2处理之间差异无统计学意义,四者显著低于90%CO2处理下的利用率(F=53.116,P<0.05);胁迫8h时,10%CO2处理下的利用率最低(15.24%),90%CO2处理下的利用率最高(27.05%),但与50%(24.47%)、70%(26.72%)CO2处理下的利用率在统计学上无显著差异(F=51.719,P>0.05).
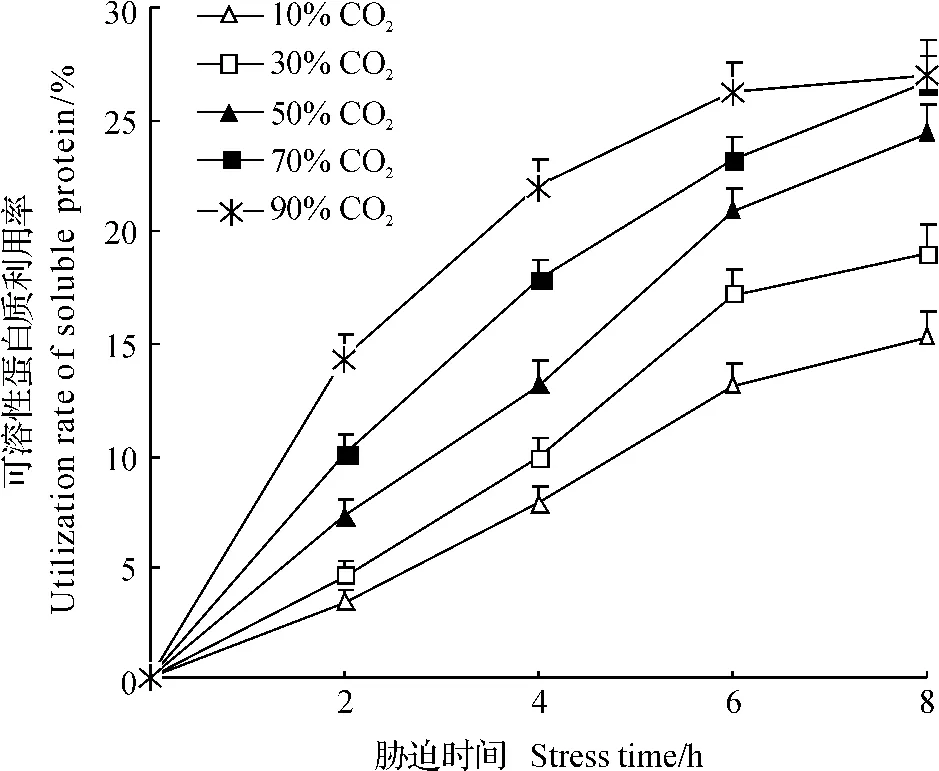
图2 在不同CO2浓度胁迫下锯谷盗对可溶性蛋白质的利用率Fig.2 Utilization rate of soluble protein in O.surinamensis under the stress of different CO2concentrations
2.2.3气调胁迫对锯谷盗脂肪含量的影响
2.2.3.1气调胁迫下脂肪含量的测定结果.在自然条件下,锯谷盗的脂肪含量为31.65μg/头.在气调胁迫下,其含量也呈现出不同差异(表4).在10%~90%CO2处理下,除90%CO2在胁迫6和8h之间锯谷盗体内的脂肪含量差异无统计学意义外,其余浓度处理下的脂肪含量在2、4、6和8h之间均逐渐减少,差异有统计学意义.在10%CO2处理下,胁迫2h的脂肪含量为30.59μg/头,显著高于胁迫4h后的28.51μg/头,6h的27.29μg/头和8h的26.58μg/头;在90%CO2处理下,胁迫2h的含量为27.08μg/头,显著高于胁迫4h(24.68μg/头)、6 h(23.30μg/头)和8h(23.09μg/头)的含量.同样,在胁迫相同时间下,随着CO2浓度的升高其含量也呈不同程度的减少.胁迫2h后,在10%CO2处理下脂肪含量为30.59μg/头,在30%、50%、70%和90%CO2处理下的含量分别为29.73、29.55、27.54和27.08μg/头,其中30%与50%CO2处理之间差异无统计学意义,其他处理浓度间在统计学上差异显著;胁迫4h时脂肪含量在各浓度下的变化与2h一致;胁迫6与8h的脂肪含量变化一致,各浓度之间在统计学上差异显著;在胁迫8h后,各浓度下的脂肪含量均降为最低,分别为26.58、26.07、25.27、23.67和23.09μg/头.

表4 CO2气调胁迫下锯谷盗的脂肪含量Table 4 Lipid content of O.surinamensis under the stress of different CO2concentrations μg/insect
2.2.3.2气调胁迫下锯谷盗对脂肪的利用率.锯谷盗对脂肪的利用率高于多糖,略低于蛋白质,但与两者的变化规律相似(图3).在10%CO2处理下,胁迫2、4、6和8h之后,其利用率显著升高(F=63.029,P<0.05),分别为3.35%、9.92%、13.78%和16.02%;在30%、50%、70%和90%CO2处理下,胁迫不同时间后,脂肪的利用规律与在10% CO2处理下的胁迫相似,其中在90%高浓度下的利用率均高于其他浓度下的相应处理,胁迫2~8h后的利用率分别为14.44%、22.02%、26.38%和27.05%(F=57.323,P<0.05).胁迫相同时间后,脂肪利用率随着气调浓度的升高而升高.在胁迫2 h时,30%与50%CO2处理之间的脂肪利用率在统计学上无显著差异,其余浓度之间差异显著,10%~90%CO2处理下的利用率分别为3.35%、5.98%、6.66%、12.93%和14.44%(F=78.015,P<0.05);胁迫4h后,各浓度下脂肪利用率的差异显著性与2 h时一致(F=66.034,P<0.05);6和8h时,各浓度之间的脂肪利用率均差异有统计学意义(F=84.228,86.162,P<0.05),其中胁迫8h后,各浓度下脂肪的利用率均达到最高,分别为16.02%、17.55%、20.18%、25.17%和27.05%.

图3 在不同CO2浓度胁迫下锯谷盗对脂肪的利用率Fig.3 Utilization rate of lipid in O.surinamensis under thestress of different CO2concentrations
在不同CO2浓度胁迫不同时间处理下,锯谷盗对3种能源物质的利用率存在一定的差异(图1~3).其在30%CO2处理6h后,锯谷盗对3种能源物质的利用率为可溶性蛋白质(17.24%)>脂肪(13.85%)>多糖(10.12%)(F=29.638,P<0.05),表现为显著降低;在50%CO2处理8h时具有一致的规律,可溶性蛋白质(24.47%)>脂肪(20.18%)>多糖(15.87%)(F=48.127,P<0.05).除此之外,在其他各处理下锯谷盗对蛋白质与脂肪的利用率间差异无统计学意义,但两者显著高于对多糖的利用率.总体上,在不同浓度、不同时间的气调胁迫下,锯谷盗对能源物质的利用率为可溶性蛋白质≈脂肪>多糖.
2.3 气调致死对锯谷盗能源物质的影响
2.3.1气调致死下锯谷盗能源物质含量测定结果
在不同CO2气调浓度致死下,锯谷盗体内可溶性蛋白质含量在70%与90%CO2处理下差异无统计学意义,其余浓度之间差异显著;在各浓度致死后,锯谷盗体内的脂肪含量在统计学上的差异显著性与可溶性蛋白质相似,而多糖含量则在各浓度之间差异显著.总体上,不同CO2浓度致死后,锯谷盗体内3种能源物质的剩余量随CO2浓度的升高而显著增加(表5).如10%CO2致死后,多糖、可溶性蛋白质及脂肪含量分别为1.47、28.98和9.84μg/头,分别为对照组的22.69%、30.33%和31.09%;在90% CO2致死下,其含量分别为2.43、44.53和16.64 μg/头,分别为对照的37.50%、46.64%和52.58%.

表5 CO2气调致死下锯谷盗多糖、可溶性蛋白质及脂肪含量Table 5 Polysaccharide,soluble protein and lipid contents of dead O.surinamensis caused by different CO2concentrations μg/insect
2.3.2气调致死下锯谷盗对能源物质的利用率
在气调致死作用下,锯谷盗体内多糖、可溶性蛋白质及脂肪3种能源物质的利用率均随着气调浓度的升高而降低(图4),且呈现不同程度的差异.多糖利用率随气调浓度的升高显著降低(F=43.276,P<0.05),在10%和90%CO2处理下的利用率分别为77.32%和62.50%.可溶性蛋白质的利用率也随气调浓度的升高而显著降低(F=33.542,P<0.05),但在70%和90%CO2处理之间差异无统计学意义.在CO2致死下锯谷盗对脂肪的利用率与对可溶性蛋白质的利用规律一致(F=36.149,P<0.05).
在相同浓度致死下,锯谷盗对3种能源物质的利用也表现出不同的显著性差异.在10%CO2致死下,锯谷盗对多糖、可溶性蛋白质及脂肪的利用率均最高,分别为77.32%、69.67%和68.91%,对多糖利用率显著高于后两者,且后两者之间差异无统计学意义(F=49.324,P>0.05);在30%CO2致死下,锯谷盗对3种能源物质利用率的差异显著性与10%CO2处理相似(F=31.193,P>0.05);在50%、70%和90%CO2致死下,锯谷盗对3种能源物质的利用率均呈现显著差异,其利用关系均为多糖>可溶性蛋白质>脂肪;在90%CO2致死下,锯谷盗对三者的利用率均为最低,分别为62.50%、53.39%和47.42%(F=53.518,P<0.05).

图4 CO2气调致死下锯谷盗对能源物质的利用率Fig.4 Utilization rate of energy substances in deadO.surinamensis caused by different CO2concentrations
3 讨论
CO2气调控害在许多害虫上都有研究报道[28],其安全环保,可媲美生物防治,且效果显著.本研究表明,CO2气调浓度越高,其LT50和LT99值越小,表明其对锯谷盗的控制效果越好.前人研究发现,在25℃用96%氮气处理5d后可完全抑制锯谷盗的危害[29],与本文10%CO2的控害效果相当(LT99=131.45h),但鉴于两者在成本及施用安全系数等方面的差异,何者更具有较好的应用前景,需进一步探讨.相关研究表明,50%CO2处理对药材甲毒力作用的LT99为72h[20],低于本文中同浓度下对锯谷盗的毒力效果,说明CO2气调对不同害虫的控害效果不同,且进一步研究表明其对昆虫的不同虫态也有不同的毒杀作用[14].这种差异也可能与试验条件相关,如温度、湿度等环境因子[30].因此,关于CO2气调的控害研究如何针对不同的昆虫、结合不同的外界环境等来充分发挥其有效性,需要进一步探索.
昆虫死亡是对气调应答的宏观表现,是在气调下体内发生的一系列生理生化反应,如昆虫体内能源物质的变化等是导致昆虫死亡的主要原因[19,31].本研究表明,CO2气调后,相较于对照,锯谷盗体内的多糖、可溶性蛋白质及脂肪3种能源物质的含量都显著减少,说明CO2处理导致了锯谷盗能源物质的大量消耗,可能是引起其死亡的重要因素.这在嗜卷书虱、嗜虫书虱(Liposcelis entomophila)及赤拟谷盗(Tribolium castaneum)等害虫上有类似报道[19,32].短时(2、4、6和8h)气调胁迫后,不管在何种处理下,锯谷盗对3种能源物质的利用率为可溶性蛋白质≈脂肪>多糖;而在CO2气调长时间胁迫至致死作用后,锯谷盗对3种能源物质的利用率为多糖>可溶性蛋白质>脂肪,表明在短时胁迫和长时间致死作用下,锯谷盗对能源物质的利用机制存在差异.这可能与逆境胁迫不同时段下不同能源物质提供三磷酸腺苷(adenosine triphosphate,ATP)的效率高低有关[33-34].另外,在长时间气调胁迫到致死过程中,锯谷盗主要依赖对多糖和可溶性蛋白质的利用,避免了对脂肪的消耗,增加了脂肪的贮存量,可能是为了更好地应对气调胁迫,有利于其对CO2气调的耐受性及抗气性的形成.已有研究表明,昆虫体内的脂肪类化合物含量越高,对高浓度CO2胁迫的抗性越强[19,34],如嗜卷书虱对甘油三酯的不同利用率决定了其对不同CO2浓度胁迫的抗性[19].
目前,关于锯谷盗气调控制的研究较少,对其作用下昆虫生理生化的研究更是缺乏.Wang等[19]研究表明,在CO2作用下,嗜卷书虱能源物质随胁迫时间的延长消耗逐渐增加,与本研究结论一致.在其他逆境胁迫下,昆虫能源物质的变化有所不同.如在化学药剂毒死蜱的短暂胁迫下,嗜卷书虱和嗜虫书虱的多糖、可溶性蛋白质显著提高,随着时间延长再下降[25],在吡虫啉胁迫下,大豆蚜(Aphis glycines Matsumura)体内能源物质有相似变化[35];在低温胁迫下,烟蚜茧蜂(Aphidius gifuensis)的可溶性蛋白质含量显著增加,而脂肪含量降低[31].这些研究说明昆虫对不同逆境有不同的应激反应,不同逆境对昆虫的致死机制不同,当然,也可能与昆虫种类有关.关于逆境下昆虫能源物质代谢的复杂性有待于进一步探讨.
总之,本研究表明,高浓度CO2胁迫可导致锯谷盗能源物质的急剧消耗,是引起其快速死亡的重要因素,是一种值得推广的储藏物害虫防治措施.而锯谷盗在短时气调胁迫与长时间致死作用下,对能源物质不同的利用机制需要进一步探讨;同时,还需要对CO2气调作用下更多能源物质的代谢进行测定,对气调胁迫下昆虫水分散失[33]、各类解毒酶和靶标酶等酶学变化[21,23]及其他分子水平的研究,都是阐明CO2气调控害机制及害虫抗气性形成机制的重要组成内容,有待深入探究.
致谢 河南农业大学闫凤鸣教授对本文的写作给予了大力帮助和指导,谨致谢意.
(References):
[1] Toews M D,Arthur F H,Campbell J F.Distribution,abundance,and seasonal patterns of stored product beetles in a commercial food storage facility.Journal of Stored Products Research,2014,56:21-32.
[2] Sharaf K,Hadid Y,Pavlíˇcek T,et al.Local genetic population divergence in a saw-toothed grain beetle,Oryzaephilussurinamensis(L.)(Coleoptera,Cucujidae).Journal of Stored Products Research,2013,53:72-76.
[3] 王明洁,国海林,孙明.温湿度对锯谷盗实验种群生长和发育的影响.南京农业大学学报,2001,24(2):57-60.Wang M J,Guo H L,Sun M.The influence of temperature and relative humidity on development of saw-toothed grain beetle,Oryzaephilus surinamensis(L.).Journal of Nanjing Agricultural University,2001,24(2):57-60.(in Chinese with English abstract)
[4] 钟建军,杨杰,吕建华,等.感染害虫的小麦加工后害虫发生动态研究.粮食科技与经济,2012,37(5):24-26.Zhong J J,Yang J,LüJ H,et al.Study on the occurrence dynamic of pests on the wheat infected by pests after processing.Grain Science and Technology and Economy,2012,37(5):24-26.(in Chinese with English abstract)
[5] 吕建华,史雅,钟建军,等.玉米象和锯谷盗在小麦粉中的种群动态研究.粮食科技与经济,2012,37(1):16-17.LüJ H,Shi Y,Zhong J J,et al.The population dynamics of Sitophilus zeamais and Oryzaephilus surinamensis in the wheat flour.Grain Science and Technology and Economy,2012,37(1):16-17.(in Chinese with English abstract)
[6] 姚渭,薛美洲,杜燕萍.八种储粮害虫趋光性的测定.粮食储藏,2005,34(2):3-5.Yao W,Xue M Z,Du Y P.The detection of phototaxis of the 8species stored-product insects.Grain Storage,2005,34(2):3-5.(in Chinese with English abstract)
[7] 王殿轩,安超楠,李兆东,等.温度对锯谷盗成虫水平和垂直运动分布的影响.河南农业大学学报,2013,47(3):330-333.Wang D X,An C N,Li Z D,et al.Influence of temperature on moving and distribution of Oryzaephilus surinamensis(Linnaeus)adults on flat surface and in vertical wheat column.Journal of Henan Agricultural University,2013,47(3):330-333.(in Chinese with English abstract)
[8] Popoola K O K,Adenuga T.Oil palm inflorescence(Elaeis guineensis Jacq.)ash and pirimiphos-methyl dust application for control of Oryzaephilus surinamensis infestation on stored date palm fruit(Phoenix dactylifera).Zoology and Ecology,2013,23(4):330-334.
[9] Rumbos C I,Dutton A C,Athanassiou C G.Comparison of two pirimiphos-methyl formulations against major storedproduct insect species.Journal of Stored Products Research,2013,55:106-115.
[10] 李能威,张晓琳,郭伟群,等.多杀菌素防治储粮害虫的研究进展.中国生物防治学报,2011,27(3):400-403.Li N W,Zhang X L,Guo W Q,et al.Research progress of spinosad as a stored-grain protectant.Chinese Journal of Biological Control,2011,27(3):400-403.(in Chinese with English abstract)
[11] 黄卫,胡飞俊,邓树华,等.三种微生物制剂对锯谷盗成虫的防治效果研究.粮油食品科技,2013,21(6):108-110.Huang W,Hu F J,Deng S H,et al.The control effect of microbial preparations on adults of Oryzaephilus surinamensis(Linnaeus).Science and Technology of Cereals,Oils and Foods,2013,21(6):108-110.(in Chinese with English abstract)
[12] 王晶磊,肖雅斌,李增凯,等.苦皮藤素和硅藻土储粮害虫室内防治效果研究.粮食储藏,2014,43(2):5-8.Wang J L,Xiao Y B,Li Z K,et al.Effect of celangulin and diatomite on stored grain insect indoor.Grain Storage,2014,43(2):5-8.(in Chinese with English abstract)
[13] Deep N Y,Tanupriya A,Monika S,etal.Microwave technology for disinfestation of cereals and pulses:An overview.Journal of Food Science and Technology,2014,51(12):3568-3576.
[14] 王殿轩,刘炎,荆纪东,等.微波对锯谷盗不同虫态的致死效应研究.粮食储藏,2011,40(3):3-7.Wang D X,Liu Y,Jing J D,et al.Lethal efficacy of microwave radiation on Oryzaephilus surinamensis(Linnaeus).Grain Storage,2011,40(3):3-7.(in Chinese with English abstract)
[15] Sehgal B,Subramanyam B,Arthur F H,et al.Variation in susceptibility of laboratory and field strains of three storedgrain insect species toβ-cyfluthrin and chlorpyrifos-methyl plus deltamethrin applied to concrete surfaces.Pest Management Science,2014,70(4):576-587.
[16] 范家霖,陈云堂,郭东权,等.电子束对锯谷盗幼虫和成虫的辐照效应研究.安徽农业科学,2010,38(24):13246-13248,13311.Fan J L,Chen Y T,Guo D Q,et al.Effect of electronicbeam irradiation on larvae and adult of Oryzaephilus surinamensis.Journal of Anhui Agricultural Sciences,2010,38(24):13246-13248,13311.(in Chinese with English abstract)
[17] 唐培安,宋伟,张婷.甲酸乙酯对锯谷盗的熏蒸活性研究.粮食储藏,2010,39(6):3-5.Tang P A,Song W,Zhang T.Fumigation activity of ethyl formate against Oryzaephilus surinamensis.Grain Storage,2010,39(6):3-5.(in Chinese with English abstract)
[18] 陈斌,李隆术.储藏物害虫生物性防治技术研究现状和展望.植物保护学报,2002,29(3):272-278.Chen B,Li L S.Biological control measures for insect pests of stored products:Current status and perspectives.Acta Phytophylacica Sinica,2002,29(3):272-278.(in Chinese with English abstract)
[19] Wang J J,Zhao Z M.Accumulation and utilization of triacylglycerol and polysaccharides in Liposcelis bostrychophila(Psocoptera,Liposcelididae)selected for resistance to carbon dioxide.Journal of Applied Entomology,2003,127(2):107-111.
[20] Gunasekaran N,Rajendran S.Toxicity of carbon dioxide to drugstore beetle Stegobium paniceum and cigarette beetle Lasioderma serricorne.Journal of Stored Products Research,2005,41:283-294.
[21] 李灿,李子忠,曹宇,等.药材甲与烟草甲羧酸酯酶和磷酸酯酶毒理学特性比较.植物保护学报,2013,40(1):78-82.Li C,Li Z Z,Cao Y,et al.A comparative of toxicological characteristics of carboxylesterases and phosphatases in Stegobium paniceum and Lasioderma serricorne.Acta Phytophylacica Sinica,2013,40(1):78-82.(in Chinese with English abstract)
[22] 唐培安,侯晓燕,宋伟,等.氮气与甲酸乙酯混合熏蒸对锯谷盗的毒力研究.粮食储藏,2013,42(5):3-6.Tang P A,Hou X Y,Song W,et al.Efficacy of ethyl formate against Oryzaephilus surinamensis by combination with nitrogen gas.Grain Storage,2013,42(5):3-6.(in Chinese with English abstract)
[23] Li C,Li Z Z,Cao Y,et al.Partial characterization of stressinduced carboxylesterase from adults of Stegobium paniceum and Lasioderma serricorne(Coleoptera:Anobiidae)subjected to CO2-enriched atmosphere.Journal of Pest Science,2009,82(1):7-11.
[24] Abbott W S.A method of computing the effectiveness of an insecticide.Journal of Economic Entomology,1925,18(2):265-267.
[25] 程伟霞,王进军,陈志永.杀虫剂胁迫下嗜卷书虱和嗜虫书虱能源物质的代谢比较.动物学研究,2005,26(5):545-550.Cheng W X,Wang J J,Chen Z Y.Comparison of the metabolisms of three energy sources in Liposcelis bostrychophila and L.entomophila(Psocoptera,Liposcelididae)under the impact of insecticides.Zoological Research,2005,26(5):545- 550.(in Chinese with English abstract)
[26] Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Analytical Biochemistry,1976,72(1/2):248-254.
[27] Colinet H,Vernon P,Hance T.Does thermal-related plasticity in size and fat reserves influence supercooling abilities and cold-tolerance in Aphidius colemani(Hymenoptera:Aphidiinae)mummies?Journal of Thermal Biology,2007,32(7):374-382.
[28] Navarro S,Jay E G.Application of modified atmospheres for controlling stored grain insects//Lawson T J.Stored Products Pest Control,British Crop Protection Council Monograph,1987,37:229-236.
[29] 严晓平,宋永成,王强,等.一定条件下96%以上氮气控制主要储粮害虫试验.粮食储藏,2010,39(1):3-5.Yan X P,Song Y C,Wang Q,et al.Test on stored grain insects exposed to over 96%nitrogen.Grain Storage,2010,39(1):3-5.(in Chinese with English abstract)
[30] 李灿,李子忠.不同温度下药材甲对二氧化碳的敏感差异.农药,2006,45(11):748-750.Li C,Li Z Z.The difference of sensitivity of Stegobium paniceum to carbon dioxide at different temperature.Agrochemicals,2006,45(11):748-750.(in Chinese with English abstract)
[31] 宋修超,崔宁宁,郑方强,等.变温贮藏僵蚜对烟蚜茧蜂耐寒能力的影响.应用生态学报,2012,23(9):2515-2520.Song X C,Cui N N,Zheng F Q,et al.Effects of fluctuating thermal regime on cold tolerance of parasitoid wasp Aphidius gifuensis.Chinese Journal of Applied Ecology,2012,23(9):2515-2520.(in Chinese with English abstract)
[32] Donahaye E,Navarro S.Comparisons of energy reserves among strains of Tribolium castaneumselected for resistance to hypoxia and hypercarbia,and the unselected strain.Journal of Stored Products Research,2000,36:223-234.
[33] Friedlander A,Navarro S,Silhacek D L.The effects of carbon dioxide on NADPH production in Ephestia cautella(Walker)pupae.Comparative Biochemistry and Physiology Part B:Comparative Biochemistry,1984,77(4):839-842.
[34] Zhou S,Criddle R S,Mitcham E J.Metabolic response of Platynota stultana pupae to controlled atmospheres and its relation to insect mortality response.Journal of Insect Physiology,2000,46(10):1375-1385.
[35] 王红,徐忠新,赵奎军,等.致死中浓度吡虫啉对大豆蚜体内能源物质含量的影响.中国生物防治学报,2014,30(5):654-659.Wang H,Xu Z X,Zhao K J,et al.Effects of median lethal concentration of imidacloprid on accumulation of energy substances in Aphis glycines(Homoptera:Aphididae).Chinese Journal of Biological Control,2014,30(5):654-659.(in Chinese with English abstract)
Influence of carbon dioxide controlled atmosphere on Oryzaephilus surinamensis Linne and its utilization of energy substances.
Cao Yu1,Yan Yufang2,Yang Wenjia1,Xiong Zhengli1,Wang Lijuan1,Li Can1*
(1.Key &Special Laboratory of Guizhou High College for Pest Control and Resource Utilization,College of Biology and Engineering of Environment,Guiyang University,Guiyang 550005,China;2.Chongqing Yudongnan Academy of Agricultural Sciences,Fuling 408000,Chongqing,China)
Oryzaephilus surinamensis Linne;carbon dioxide controlled atmosphere;energy substance;polysaccharide;soluble protein;lipid
Q 966;S 435.6
A
10.3785/j.issn.1008-9209.2015.04.072
贵州省国际合作项目(黔科合外G字[2013]7047号);贵州省普通高校生物多样性与应用生态学特色重点实验室项目(黔教合KY字[2012]013);贵州省重点学科项目(黔学位合字ZDXK[2013]08).
李灿(http://orcid.org/0000-0002-0536-8555),E-mail:lican790108@163.com
联系方式:曹宇(http://orcid.org/0000-0002-4022-4266),E-mail:yucaosuccess@126.com
2015-04-07;接受日期(Accepted):2015-07-13;< class="emphasis_bold">网络出版日期
日期(Published online):2015-10-27
URL:http://www.cnki.net/kcms/detail/33.1247.S.20151027.1122.002.html
猜你喜欢
四川农业与农机(2019年4期)2019-08-29 06:05:54
中国化肥信息(2019年6期)2019-01-19 13:10:42
经济技术协作信息(2018年5期)2019-01-19 08:39:16
中成药(2018年8期)2018-08-29 01:28:34
消费导刊(2017年24期)2018-01-31 01:29:29
印制电路信息(2015年6期)2015-12-30 12:57:48
医学研究杂志(2015年9期)2015-07-01 17:28:24
中国当代医药(2015年9期)2015-03-01 02:02:13
中国果业信息(2015年6期)2015-01-23 17:38:07
食品科学(2013年24期)2013-03-11 18:30:53
