
酵母培养物对牦牛瘤胃发酵及甲烷产量的影响
2015-07-31 00:45孙红梅曹连宾郝力壮等
江苏农业科学 2015年3期
孙红梅 曹连宾 郝力壮等
摘要:为研究酵母培养物(YC)对牦牛瘤胃体外培养青藏高原高寒草甸4期牧草发酵产物及甲烷产量的影响,选取3头健康的4岁瘘管牦牛为瘤胃液供体,采用体外产气法,全年4期牧草作为发酵底物,每期牧草中分别添加0、1%、2%、3%、4% YC,筛选YC在每期牧草中的最佳添加量。结果显示,YC可使枯草期产气量下降(P<0.05),对返青期和枯黄期产气量影响不显著(P>0.05),添加2%YC组青草期产气量显著下降(P<0.05);YC对发酵液的pH值和铵态氮浓度影响不显著;在体外培养条件下,在维持牦牛瘤胃正常发酵环境且有改善牧草发酵后能量转化率的情况下,分别在枯草期、返青期、青草期和枯黄期添加3%、2%、1%、2%YC可使其甲烷生成量比对照组显著降低(P<005)。综合考虑各试验组,4期中枯黄期甲烷生成量最少,其中添加3%YC组最低。
关键词:酵母培养物;牦牛;青藏高原;瘤胃;体外产气;牧草;发酵参数;甲烷
中图分类号: S858.236.9 文献标志码: A
文章编号:1002-1302(2015)03-0177-06
酵母培养物(yeast culture,YC)是一种将酵母菌经特定厌氧发酵培养后形成的微生态制品,主要活性成分是酵母菌的各类代谢产物、发酵变异的培养基及少量酵母细胞,包括肽、有机酸、寡糖、氨基酸和一些芳香烃类物质[1]。YC作为反刍动物日粮添加剂的趋势已越来越明显,它有改善动物对饲料的消化率、增加瘤胃微生物蛋白(MCP)[2]、增加泌乳生产性能、提高动物免疫力的优势[3],是继抗生素类添加剂之后作为绿色健康标签而倍受瞩目的新型饲料添加物。基于这些优势,目前很多研究集中在YC对于动物瘤胃发酵、生产性能和饲料消化率的影响方面,而涉及对动物甲烷生成量影响的报道较少,在牦牛研究领域上也是一个空缺。因此,本试验旨在探讨YC对牦牛甲烷生成量的影响和对瘤胃发酵的作用。
1 材料与方法
1.1 试验地点
青海省高原放牧家畜营养与生态国家重点实验室培育基地、青海省高原放牧家畜动物营养与饲料科学重点实验室和青海省高原牦牛研究中心进行牧草营养成分测定和体外产气培养及相关指标评定。
1.2 试验材料
根据青藏高原高寒草地实际情况采集4期牧草样品,在三江源区核心区设置采样样地,采用1.0 m×1.0 m样方,齐地面刈割,挑出不可食部分,称质量并记录,风干称质量带回实验室,粉碎,过40目筛备用[4]。YC为市售商品。
1.3 试验动物管理及日粮
选择3头健康、体质量接近、装有永久性瘤胃瘘管的4岁牦牛作为瘤胃液供体,单独饲喂。试验日粮为燕麦青干草和小麦秸秆组成,全天自由采食,自由饮水,预饲15 d后开始正试期,晨饲前采集瘤胃液,采集至保温瓶中迅速带到实验室。
1.4 牧草常规养分分析法
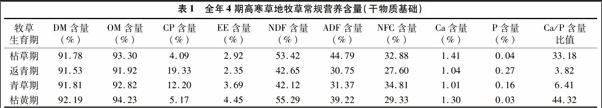
牧草的干物质(DM)、有机物质(OM)、粗蛋白质(CP)、粗脂肪(EE)、酸性洗涤纤维(ADF)、中性洗涤纤维(NDF)、非结构性碳水化合物(NFC)、灰分(Ash)、钙(Ca)、磷(P)含量的测定均参照文献[5]中的推荐方法。
1.5 体外产气法
分别以全年4期天然混合牧草作为发酵底物,4期牧草培养为4个单因素独立试验,底物为220 mg,另外设置空白组(仅瘤胃液和培养液),作为产气量校正。YC添加水平为0、1%、2%、3%、4%,每个水平设3个重复。通过瘤胃瘘管在晨饲前抽取牦牛瘤胃液混合后立即放入保温瓶中备用,防止微生物区系发生变化。采用Menke 等的方法[6]准备缓冲液,并将缓冲液与瘤胃液以2 ∶1的比例混合,将混合好的培养液在发酵前通入CO2保证厌氧环境后,分液至对应编号的培养管内(30 mL),放入水浴(39±0.5) ℃的摇床上开始培养。
1.6 产气量的记录
从培养管放入培养箱中开始计时,在0、2、4、6、8、12、14、16、24、36、48 h快速读数记录。当读数超过80 mL时,为了防止气体超过刻度而无法读数,需要排气,收集气体到对应编号集气袋中,记录排气后的刻度值。
1.7 发酵参数测定
体外培养48 h后,将培养管取出放入冰水混合物中停止发酵。将发酵液排到50 mL玻璃管内,测定pH值;取部分发酵液经离心(4 000 r/30 min),取上清液进行氨态氮(NH3-N)浓度测定,测定方法参照改进的比色法[7]进行。
产气量(mL)=该时间段内培养管产气量(mL)-对应时间段内空白管平均产气量(mL);短链脂肪酸含量SCFA(mmol/L)=0.022 2×GP-0.004 25(r2=0.94)[8];有机物质消化率OMD(%)=0.904 2×GP+0.049 2×CP+0038 7×CA+16.49(n=85,r2=0.93);代谢能ME(MJ/kg,干物质)=0136×GP+0.005 7×CP+0.000 286×CP2+220(n=200,r2=0.94);泌乳净能NEL(MJ/kg,干物质)=0.096×GP+0003 8×CP+0.000 173×CP2+0.54(n=200,r2=0.93)[6]。其中:GP为24 h净产气量,mL;CP为粗蛋白含量,%;CA为灰分含量,%。
发酵气体甲烷含量的测定委托兰州中科安泰分析科技有限公司进行,分析条件[9]为仪器GC 3420 (北分瑞利气相色谱仪);检测器FID(氢火焰检测器);柱长30 m,内径 0.53 mm,膜厚20 μm;柱温80 ℃;汽化温度120 ℃;检测温度130 ℃;进样量10 μL。
1.8 产气动力学参数
1.9 数据统计分析
采用Excel 2003进行数据整理,SAS 9.1统计软件中单因素试验统计分析采用Duncans新复极差法进行多重比较,在0.05水平进行差异显著分析。
2 结果与分析
2.1 4期牧草常规营养成分
全年4期高寒草地牧草常规营养含量(干物质基础)见表1。
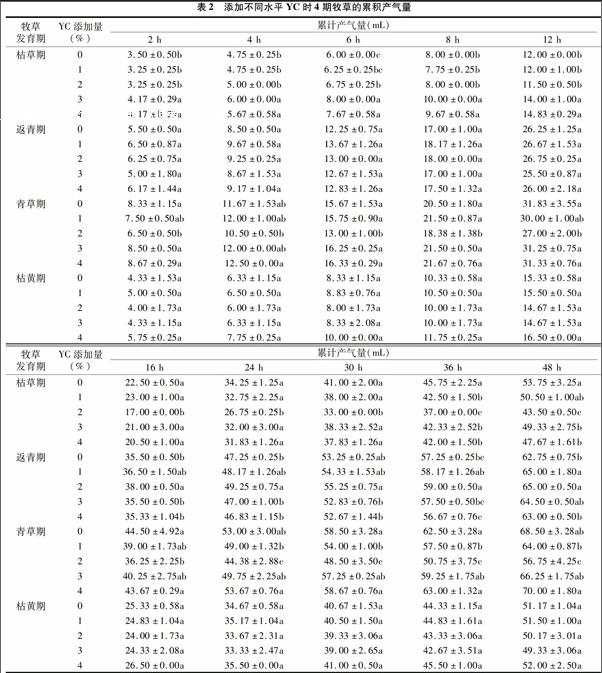
2.2 添加不同水平YC时4期牧草的累积产气量
体外产气试验结果(表2)显示,以枯草期牧草为发酵底物,添加YC降低了48 h累积产气量,添加1%YC组与对照组差异不显著(P>0.05),当添加量加大时,48 h累计产气量显著下降(P<0.05),但在发酵前12 h,枯草期添加3%、4%YC组,返青期添加1%、2%YC组,枯黄期添加4%YC组的产气量均较其对照多(P<0.05)。在返青期,添加YC可以提高牧草的48 h累积产气量,1%、2%YC组较对照组显著提高(P<005),前12 h,YC对产气量影响较小,之后出现显著变化趋势。在青草期,2%YC添加组各时间点的产气记录显著低于对照(P<0.05)。枯黄期试验未发现YC对累积产气量影响显著。
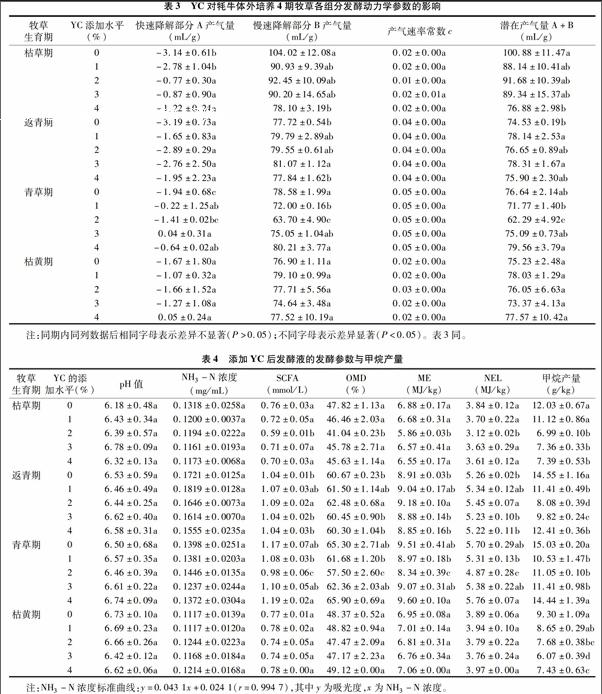
由表3可知,当在枯草期添加≥2%YC时,快速降解部分的产气量A显著增大(P<0.05),而慢速降解部分的产气量B减小,其中添加4%YC组显著减小(P<0.05),A+B下降,与48 h累积产气量的下降吻合。YC对返青期A影响较小,因此在发酵前12 h各组间累积产气量差异不显著(P>005),B增加导致各组48 h累积产气量升高;青草期A除添加2%YC组外均显著升高(P<0.05),而B在添加1%、2%YC时显著下降(P<0.05),添加2%YC组的A+B显著下降(P<0.05),相应48 h累积产气量也显著减少(P<0.05);YC对枯黄期各部分发酵影响不显著(P>0.05)。各期YC不同添加水平对产气速率常数c影响不显著(P>0.05)。
2.3 添加不同量YC时4期牧草的发酵参数与甲烷产量
由表4可知,在4期牧草体外培养试验中添加YC对pH值和NH3-N浓度影响在各期内均不显著(P>0.05);SCFA、OMD、ME、NEL的值在各期试验统一添加水平时的变化趋势一致,枯草期除添加2%YC组较对照组显著下降(P<0.05)外,其余添加量组变化较小(P>0.05),随添加水平的增加呈先减小后增大的趋势;返青期这4个值受YC添加量的影响,随添加量加大呈先增大后减小趋势,添加2%YC组的各值较对照显著增大(P<0.05);在青草期添加2%YC时,这4个值比对照显著小(P<0.05),当YC的添加量>2%时回升,但与对照组差异不显著(P>0.05);在枯黄期添加YC后,这4个值均与对照差异不显著(P>0.05)。YC的存在可减少各期牧草体外发酵的甲烷产量,其中在枯草期添加≥2%YC组的甲烷产气量显著减小(P<0.05);返青期YC各添加水平均能显著降低甲烷产量(P<0.05);青草期添加1%、2%、3%YC组的甲烷产量显著减小(P<0.05),添加4%YC组的甲烷产量也低于对照,但差异不显著(P>0.05);枯黄期添加>1% YC能使甲烷产量显著减小(P<0.05)。
经双因素方差分析考察甲烷生成量,得到青草期>返青期>枯草期>枯黄期,且两两之间差异显著(P<0.05)。其中,对照组的甲烷产量显著高于各YC添加组(P<0.05),添加1%、4%YC组仅次之,差异不显著(P>0.05),与生成甲烷最少的添加2%、3%YC组也均差异显著(P<0.05),不同期与不同YC添加水平间存在互作(P<0.05),即不同期要求不同的YC添加量达到最佳的减少甲烷产量效果。经考核发现,枯黄期添加3%YC时的甲烷生产量最少。
3 结论与讨论
关于YC在反刍动物瘤胃内的作用机理存在控氧理论、小肽营养代谢扳机理论和营养理论等3种理论。控氧理论认为,培养物质仍存在一些酵母菌,它作为好氧菌将瘤胃中的氧气消耗掉后有利于瘤胃保持厌氧环境,对瘤胃微生物厌氧菌群(如纤维菌)生长发酵形成有利条件;小肽营养代谢扳机理论认为,培养物中所含的活性类似于小肽结构物质会促进瘤胃微生物活性而改变瘤胃发酵;营养理论认为,培养物中代谢产物包括有机酸、维生素、矿物质等,有利于微生物繁殖代谢而增强其发酵活性[12]。
3.1 YC对4期牧草体外发酵产气量的影响
瘤胃发酵产气主要来自饲料中OM的降解[6],这些气体包括一些挥发性酸类、CO2、H2及甲烷等[13],产气量反映饲料的可发酵程度,可用以衡量饲料营养价值[14]。枯草期前8 h产气量升高,促进牧草OM中易降解部分发酵,A增大。有研究证实,YC能增加纤维菌的数量和活性[3,15],酵母菌是好氧菌,其代谢过程会减少瘤胃内的氧气,加强形成瘤胃厌氧环境,促进纤维分解菌等厌氧菌的代谢。饲料碳水化合物被纤维分解菌快速降解产生大量气体,而随微生物迅速增殖加大了对瘤胃内挥发性营养物质的消耗,因此累积产气逐渐减少,同时A的增大弥补了B减少的量,因此仅添加4%YC组的产气量显著下降;在返青期,低浓度YC对牦牛瘤胃微生物的刺激作用较强,与OMD的增大一致,累积产气量增多;在青草期期添加2%YC时,底物发酵被抑制,因为OMD显著下降(P<0.05),发酵自始至终产气显著低于对照(P<0.05),说明添加2%YC对以青草期为底物的瘤胃微生物有抑制作用,其原因可能是青草期牧草营养相对均衡,P含量和Ca含量比例合理,而YC中也含矿物质和一些酶类,添加2%YC破坏了这种平衡,对微生物活性和对饲料降解不利;而添加YC对枯黄期牦牛瘤胃体外发酵产气和OMD无影响,各添加水平下的变化与对照组差异均不显著(P>0.05),A+B的变化趋势与 48 h 累积产气量趋势一致。4期试验添加YC对累积产气量的影响变化不一致,主要受各期牧草营养成分影响,主要受饲料中ADF和NDF含量差异影响[16]。
3.2 YC对4期牧草体外培养48 h后各发酵参数的影响
通常情况下,反刍动物采食后瘤胃正常的pH值在5.5~7.5之间变动[17],瘤胃酸碱环境与动物消化机能密切联系,大至瘤胃原虫,小至诸多作用酶等都对瘤胃内pH值很敏感,维持瘤胃正常的酸碱环境对机体正常代谢尤为重要[18]。氨态氮是饲料中蛋白氮和非蛋白氮等含氮物质被微生物分解代谢形成的终产物,大部分会被瘤胃微生物利用合成MCP[19],成为反刍动物的蛋白质来源[20],其浓度是饲料被分解程度和瘤胃微生物合成利用情况的平衡动态值,最适为0.065 8~0367 0 mg/mL[21],即0.06~0.30 mg/mL[22]。分别进行的4期试验结果显示,与对照组相比,YC不同添加水平对pH值和NH3-N浓度影响均不显著(P>0.05),与姜艳美等的结论[23]一致,说明YC不会改变动物瘤胃的正常运作机能,可使牦牛瘤胃内适宜酸碱环境和氮的循环保持良好平衡。虽然按照理论推测,因YC可促进纤维的快速降解,会加速有机酸累积,而使瘤胃pH值下降,但诸多试验并未发现pH值下降,而是维持在恒定范围内,这是因为YC中含有的二羧酸可通过促进瘤胃内乳酸利用菌的活性增强对乳酸的循环利用[24-25],不致瘤胃形成过酸环境,可达到稳定pH值的效果,调节瘤胃内对酸环境敏感的菌群,增加瘤胃微生物菌群多样性[26],因此可添加YC防止动物因日粮中添加精料而导致酸中毒的现象。据报道[27],YC能使饲料中的CP降解率提高,同时促进MCP合成,瘤胃细菌数增加是添加YC最显著的变化,这是YC可提高动物生产力的源动力[28]。瘤胃微生物加快增殖,生成更丰富的MCP为机体提供蛋白质,因此YC可改善瘤胃内氮循环。本试验中的NH3-N浓度均未受影响,但蒋小军等的试验结果显示,YC添加后NH3-N浓度降低,因MCP合成迅速增加[29],与微生物数量增加的报道一致,同样是YC的积极效应;而Yoon等发现,YC能刺激蛋白分解菌增长[30],一方面导致NH3-N生成增多,另一方面MCP合成加快,因此本试验NH3-N浓度无变化。
SCFA也被认为是挥发性脂肪酸(VFA),可被瘤胃上皮细胞和瘤胃微生物直接吸收,是主要来自饲料中碳水化合物的降解产物。试验证明,YC能促进纤维素及木质素的降解,改善瘤胃发酵,在总VFA中乙酸和丙酸占绝大多数,YC活性因子对瘤胃微生物群系有选择性刺激作用,据报道,YC能提高产乙酸菌活性,增强产乙酸菌利用氢,可使乙酸产量增加5倍[31],因为氢更多被用于合成乙酸,可被产甲烷菌利用的氢减少,这是本试验得到添加YC后甲烷生成量减少的原因之一。在枯草期和青草期,随YC添加量加大,SCFA含量先下降后上升,添加2%YC时显著下降(P<0.05),而返青期的变化为先上升后下降,添加2%YC时显著上升(P<0.05),这些变化与产气量和OMD变化紧密相关,都源自YC对瘤胃微生物的刺激作用,进而造成饲料被分解程度的差异。在枯黄期添加YC后,SCFA与对照差异不显著。这与Williams等的结论[32]一致。不同水平YC对4期牧草产生的SCFA量影响变化不一致,主要受牧草营养类型差异的影响,有研究指出,VFA量会随YC添加时间及日粮类型变化而变化[33]。有研究显示,活性酿酒酵母能提高真菌对纤维的分解能力,显著提高甲酸、乳酸、乙酸等的浓度[34],因此SCFA浓度增大;但有报道,酵母培养物对总VFA无影响[32]。
3.3 YC对4期牧草体外培养生成甲烷量的影响
联合国专委会表示,每年仅反刍动物向大气中排放的甲烷量达85 Tg,占全球温室效应危害的2%左右[35],与此同时,反刍动物瘤胃甲烷的产生也浪费掉2%~12% 的饲料总能[36],面临高寒牧区天然草场日渐退化的压力,在可持续发展和减排温室气体的胁迫下,有效抑制甲烷的研究就变得尤为重要。据报道,产乙酸菌生成乙酸的过程需要H2的参与,YC能刺激瘤胃中产乙酸菌利用H2,甚至添加活酵母菌能使乙酸产量及产乙酸菌利用H2效率提高5倍,这样增强与产甲烷菌竞争氢的作用,会有效减少甲烷的生成。改变瘤胃发酵方式,乙酸含量/丙酸含量的值减小[25],丙酸大量形成消耗氢,可减弱甲烷的生成。但这种效果与日粮类型关联,因此出现不一致的结论[37]。甲烷的减少还与原虫数量减少相关,YC可减少瘤胃中45%的原虫[38]。枯草期添加≥2% YC可显著降低甲烷生成量,但添加2%YC组的SCFA、OMD、ME、NEL显著减小(P<0.05);返青期添加YC各组的甲烷生成量均下降(P<005),其中添加2%YC组对底物发酵程度的促进作用更大,能量转化率显著增大(P<0.05);青草期添加4%以下YC组的甲烷生成量显著下降(P<0.05),对添加1%YC组的饲料能量转换无影响,而添加2%YC组的能量转化效率下降,虽然4%YC使SCFA含量升高,OMD、ME、NEL均提高(P<005),但甲烷未见减少;在不影响能量转化时,添加2%YC可显著减少枯黄期的甲烷生成量(P<0.05)。
枯草期产气量因YC的添加而下降(P<0.05),对返青期和枯黄期产气量影响不显著(P>0.05),添加2%YC组的产气量在青草期显著下降(P<0.05)。在维持牦牛瘤胃正常发酵环境和有效改善饲料能量转化率的情况下,分别在枯草期、返青期、青草期、枯黄期添加3%、2%、1%、2%YC可使甲烷生成量显著降低(P<0.05)。其中,枯黄期的甲烷生成量最少,且该生育期添加3%YC组为各试验组的最低值。
参考文献:
[1]张爱忠. 酵母培养物对内蒙古白绒山羊瘤胃发酵及其它生理功能调控作用的研究[D]. 呼和浩特:内蒙古农业大学,2005.
[2]Erasmus L J,Robinson P H,Ahmadi A,et al. Influence of prepartum and postpartum supplementation of a yeast culture and monensin,or both,on ruminal fermentation and performance of multiparous dairy cows[J]. Animal Feed Science and Technology,2005,122(3/4):219-239.
[3]张鹏飞,于安乐,高 巍. 酵母培养物对反刍动物作用的研究进展[J]. 吉林畜牧兽医,2007,28(6):17-20.
[4]郝力壮,王万邦,王 迅,等. 三江源区嵩草草地枯草期牧草营养价值评定及载畜量研究[J]. 草地学报,2013,21(1):56-64.
[5] 张丽英. 饲料分析及饲料质量检测技术[M]. 2版. 北京:中国农业大学出版社,2003.
[6] Menke K H,Steingass H. Estimation of the energetic feed value[J]. Animal Research and Development,1988,28:7-55.
[7]冯宗慈,高 民. 通过比色测定瘤胃液氨氮含量方法的改进[J]. 内蒙古畜牧科学,1993,31(4):40-41.
[8]Makkar H S. In vitro gas method for evaluation of feeds containing phytochemicals—a review[J]. Animal Feed Science and Technology,2005(1):291-302.
[9]郭雪峰. 内蒙古白绒山羊甲烷产生量估测模型的建立及其影响因素的研究[D]. 呼和浩特:内蒙古农业大学,2008.
[10]rskov E R,McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage[J]. Journal of Agricultural Science,1979,92:499-503.
[11]黄雅莉,邹彩霞,韦升菊,等. 体外产气法研究半胱胺对水牛瘤胃发酵参数和甲烷产量的影响[J]. 动物营养学报,2014,26(1):125-133.
[12]Kou V. Mode of action of yeast culture(YEA-SACC1026)for stimulation of rumen fermentation in buffaio calves[J]. Journal of the Science of Food and Agriculture,1998(3):407-413.
[13]Menke K H,Raab L,Salewski A,et al. The estimation of the digestibility and metabolizable energy content of ruminant feeding stuffs from the gas production when they are incubated with rumen liquor in vitro[J]. Journal of Agricultural Science,1979,93:217-222.
[14]王小晶,董国忠,刘智波. 青蒿提取物和大黄对山羊瘤胃发酵和甲烷产量的影响[J]. 畜禽业,2009(4):52-55.
[15]哈斯花. 体外发酵产气技术在饲料营养价值评定中的作用[J]. 当代畜禽养殖业,2013(1):15-19.
[16]郑晓中. 酵母培养物对反刍家畜营养作用的研究[J]. 饲料研究,1996(11):2-5.
[17]Yoon I K,Garrett J E,Cox D J. Effect of yeast culture supplementation to alfafa grass hay diet on microbial fermentation in continuous culture of rumen contents[J]. Animal Science,1997,75:91-98.
[18]National Research Council.Nutrient requirements of beef cattle[S]. 7th ed. Washington D C:National Academy Press,1996:85-97.
[19]Leng R A,Nolan J Y. Nitrogen metabolism in the rumen[J]. Journal of Dairy Science,1984,67(5):1072-1089.
[20]Srinivas B,Gupta B N. Rumen fermentation,bacterial and total volatile fatty acid(TVFA) production rates in cattle fed on urea-molasses-mineral block licks supplement[J]. Animal Feed Science and Technology,1997,65(1/2/3/4):275-286.
[21]Coleman G S,Sandford D C. Uptake and utilization of bacteria,amino-acids and nucleic-acid components by the rumen ciliate Eudiplodinium maggii[J]. Journal of Applied Bacteriology,1979,47(3):409-419.
[22]王加启,李树聪. 不同日粮类型对奶牛营养代谢影响的研究[C]//动物营养研究进展论文集.北京:中国农业科学技术出版社,2004:124-133.
[23]姜艳美,王加启,邓露芳,等. 酵母菌培养物对瘤胃发酵的影响[J]. 动物营养学报,2008,20(1):92-97.
[24]冯仰廉. 反刍动物营养学[M]. 北京:科学出版社,2004.
[25]朱金凤,张丁华,王艳丰,等. 酵母及酵母培养物对反刍动物瘤胃内环境影响的研究进展[J]. 河南农业,2008,5(10):33-35.
[26]李声永,王加启,龚月生,等. 酵母培养物在反刍动物日粮中的应用研究进展[J]. 中国畜牧兽医,2002,29(5):18-22.
[27]龙 玲. 酵母培养物对肉牛瘤胃发酵及纤维消化的影响[D]. 杨凌:西北农林科技大学,2003.
[28]Wallace R J. Ruminal microbiology,biotechnology,and ruminant nutrition:progress and problems[J]. Journal of Animal Science,1994,72(11):2992-3003.
[29]蒋小军,王康宁. 酵母培养物对山羊瘤胃发酵的影响[J]. 畜禽业,2005(7):26-27.
[30]Yoon I K,Stern M D. Effects of saccharomyces cerevisiae and aspergillus oryzae cultures on ruminal fermentation in dairy cows[J]. Journal of Dairy Science,1996,79(3):411-417.
[31]王志博. 酵母培养物破壁条件的优化及其对底物发酵的影响[D]. 哈尔滨:东北农业大学,2007.
[32]Williams P E V,Tait C A G,Innes G M,et al. Effects of the inclusion of yeast culture (Saccharomyces cerevisiae plus growth medium) in the diet of dairy cows on milk yield and forage degradation and fermentation patterns in the rumen of steers[J]. Journal of Animal Science,1991,69:3016-3026.
[33]刘大程. 不同品质粗饲料日粮及添加酵母培养物对绵羊瘤胃发酵及纤维分解菌的影响[D]. 呼和浩特:内蒙古农业大学,2007.
[34]Chaucheyras E,Fonty G,Benin G,et al. Effects of live Saccharomyces cerevisiae cells on zoospore germination,growth and cellulolytic activity of the rumen anaerobic fungus,Neocallimatix frantalis MCH3[J]. Current Microbiology,1996,31:201.
[35]王嘉厚. 反刍动物甲烷产生与控制措施[J]. 畜禽业,2013(10):46.
[36]Johnson K A,Johnson D E. Methane emissions from cattle[J]. Journal of Animal Science,1995,73(8):2483-2492.
[37]毛宏伟,蒿迈道. 啤酒酵母培养物(BYC)调节牛瘤胃发酵的试验研究初报[J]. 黄牛杂志,1993,04(4):22-26.
[38]Frumholta P P,Newbold J,Wallace R J. Influence of Aspergillus oryae fermentation extract on the fermentation of a basal ration in the rumen simulation technique(Rustic)[J]. Journal of Agriculture Science,1989,113(2):169-172.
猜你喜欢
环境卫生工程(2021年5期)2021-11-20
湖南饲料(2021年4期)2021-10-13
Defence Technology(2019年6期)2020-01-07
西南石油大学学报(自然科学版)(2019年5期)2019-12-20
意林·全彩Color(2019年8期)2019-11-13
扬子江(2019年3期)2019-05-24
录井工程(2017年3期)2018-01-22
领导文萃(2017年10期)2017-06-05
中国石油大学学报(自然科学版)(2015年2期)2015-11-10
石油化工应用(2014年8期)2014-03-11
