
Cr(Ⅵ)盐胁迫下小麦种子中淀粉酶活性的响应
2015-07-31 19:38张艳杰李桂玲王金水等
江苏农业科学 2015年3期
张艳杰 李桂玲 王金水等

摘要:研究重金属离子Cr(Ⅵ)在不同浓度下对小麦种子萌发及其幼苗生长的影响。结果表明:Cr(Ⅵ)对小麦种子萌发和幼苗生长影响的临界值为18 mg/L,即浓度低于18 mg/L的Cr(Ⅵ)促进小麦种子的萌发和幼苗的生长,高于 18 mg/L则抑制小麦种子的萌发和幼苗的生长;Cr(Ⅵ)对根生长的抑制作用大于对芽生长的抑制作用;高浓度的 Cr(Ⅵ) 对小麦种子萌发和幼苗生长的抑制是通过抑制淀粉酶的活性来实现的。
关键词:Cr(Ⅵ);小麦种子;萌发;淀粉酶;同工酶
中图分类号: Q946.5;Q945.78 文献标志码: A
文章编号:1002-1302(2015)03-0073-03
铬(chromium,Cr)是自然环境中存在的1种重金属元素,同时也是1种重要的环境污染物,被列为工业“五毒”之一。铬在自然界中主要以三价铬(Cr3+)、六价铬[Cr(Ⅵ)]的化合物形式存在,且后者的毒性比前者更强[1-2]。已有研究证明,Cr(Ⅵ)不是植物生长发育的必需元素,在植物体内过量积累对植物的生长发育有明显的抑制作用[3]。近年来,随着印染、电镀、化工、电子等行业的发展,越来越多的含铬废水、废渣被排放到水体中,导致农田灌溉用水污染严重,进而对农作物的生长发育造成了巨大伤害,这不仅降低了农产品的产量和品质,而且能够通过食物链进入人体,危害人类健康[4]。
小麦(Triticum aestivum Linn.)是我国重要的粮食作物,对于小麦的研究具有重要的经济价值和现实意义,其产量和品质也越来越受到关注[5]。有关调查显示,受重金属污染的农田耕地面积占总耕地面积的1/5,其中受铬污染的尤为严重[6],因此研究铬污染对小麦种子生长发育的影响极其重要。
目前,关于Cr(Ⅵ)盐对小麦生长发育影响的研究多数是集中在对萌发及幼苗生长阶段形态学指标的检测分析方面,而关于Cr(Ⅵ)盐对小麦不同发育阶段形态指标的影响是如何实现的研究则鲜有报道。本研究围绕这一问题,以小麦为试验材料,采用室内水培法对小麦施以不同浓度梯度的Cr(Ⅵ)溶液,研究Cr(Ⅵ)盐对小麦种子萌发及幼苗生长期间关键酶——淀粉酶活性的影响,以期为重金属Cr(Ⅵ)对小麦生长毒害机理的研究,以及耐、抗重金属Cr(Ⅵ)盐品种的选育提供理论支持和依据。
1 材料与方法
1.1 材料与试剂
小麦选用温麦6号,产自河南省黄泛区农场。试剂为华美生物工程公司生产的重铬酸钾(K2Cr2O7,分析纯)。
1.2 试验方法
1.2.1 种子处理与试验设计 挑选大小均一、籽粒饱满的温麦6号种子,先用5%H2O2处理5 min进行表面消毒,然后用蒸馏水冲洗3~5遍,再加蒸馏水没过种子最上层2 cm左右,浸种12 h。将小麦种子腹沟向下排列在铺有双层滤纸的培养皿中,每皿40粒,使每粒种子间留有均匀的间距,以防发霉种子对健康种子的感染。在培养皿中分别加入5 mL不同梯度浓度(3、5、10、15、18、20、25、30 mg/L,以Cr(Ⅵ)计)的K2Cr2O7溶液;对照(CK)用蒸馏水培养,每个浓度梯度均设置6个重复,置于室温、自然光照下培养萌发,处理时间分别为3、7 d。所设的6个重复中3个重复用于培养3 d的形态学和生理指标测定;另外3个重复用于培养7 d的形态学和生理学指标测定。
1.2.2 形态学指标的测定 测定培养3、7 d小麦的芽长、根长。记录的数据用Excel 2003进行成组数据t检验的统计分析。
1.2.3 生理学指标测定 待种子培养3、7 d后,分别去掉种皮,称取0.5 g置于预冷的研钵中,加入少量石英砂和2 mL蒸馏水,研磨匀浆后转入10 mL离心管中,用蒸馏水定容至10 mL,4 ℃、10 000 r/min条件离心15 min,上清液即为粗酶液。取上清液,4 ℃下保存备用。
采用聚丙烯酰胺凝胶电泳法(PAGE)测定萌发种子淀粉酶同功酶活性,分离胶浓度为7.2%,浓缩胶浓度为3.2%[7]。于日光灯下观察记录酶谱,用数码相机进行拍照。须要测量的数据有脱色前后胶长、指示剂游动距离、酶带长。计算迁移率相关公式为V=mE。式中:V为溶液中带电粒子的移动速度,cm/s;m为粒子的有效迁移率,cm3/(V·s);E为电场,V/m。同时,采用分光光度法(3,5-二硝基水杨酸法)测定萌发种子的α-淀粉酶同工酶活性[8]。
2 结果与分析
2.1 Cr(Ⅵ)溶液对小麦主要形态学指标的影响
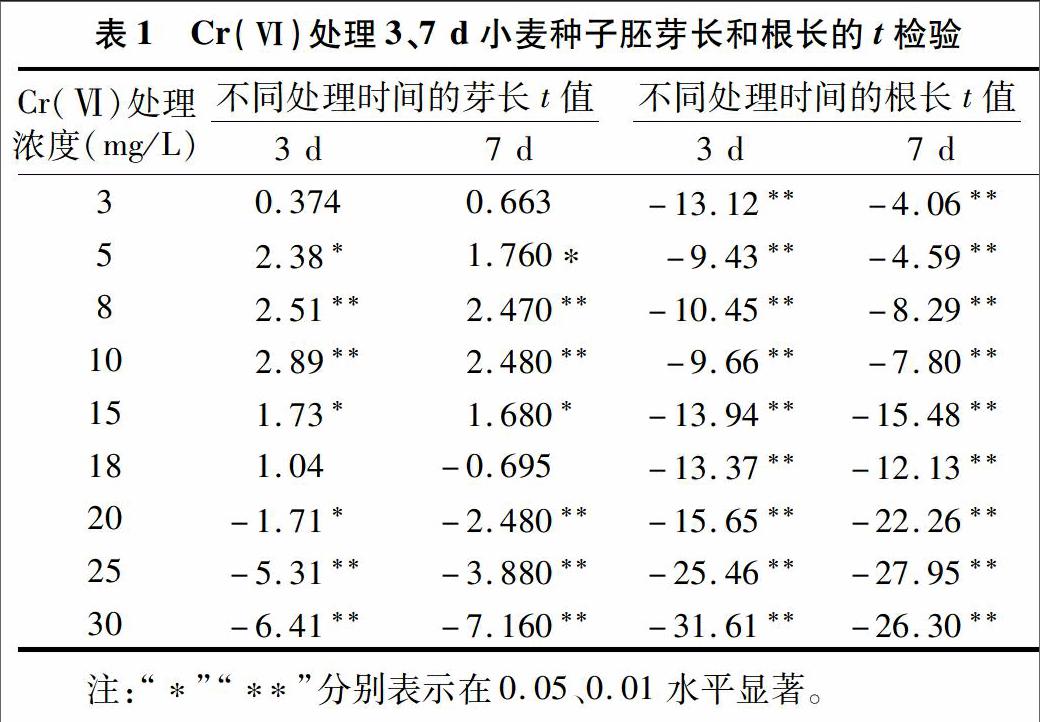
2.1.1 Cr(Ⅵ)溶液对小麦胚芽的影响 如图1所示,用不同浓度梯度Cr(Ⅵ)盐处理后,萌发3 d的小麦胚芽芽长在低浓度处理时略高于对照;相反,在高浓度处理时比对照低。与对照相比,Cr(Ⅵ) 处理浓度为3~15 mg/L时,可以刺激小麦胚芽的生长,对小麦胚芽生长起到了促进作用,其中处理浓度在 10 mg/L 时对小麦胚芽的生长有极显著的促进作用;当 Cr(Ⅵ) 处理浓度提高至18 mg/L时,小麦胚芽的长度开始呈现下降趋势,芽长与对照基本相同;处理浓度为20 mg/L时,小麦种子胚芽的生长受到了极显著的抑制;随着Cr(Ⅵ)盐浓度的继续升高,受抑制程度更为严重,结果见表1。处理7 d的小麦胚芽生长规律与处理3 d的小麦胚芽生长规律总体趋势表现一致,结果如图2所示。因此推测,18 mg/L的Cr(Ⅵ)处理浓度为影响小麦种子萌发和幼苗生长的临界值。
2.1.2 Cr(Ⅵ)溶液对小麦胚根的影响 用不同浓度梯度的Cr(Ⅵ)处理后,与对照相比,对萌发3、7 d的小麦胚根的生长均有抑制作用,且均达到了极显著水平,结果见图1、图2、表1。这说明用不同浓度Cr(Ⅵ)处理后,小麦胚根的生长状况与胚芽不同;高浓度(>18 mg/L)的Cr(Ⅵ)处理时,Cr(Ⅵ)浓度越高,抑制效应越强,且对小麦种子胚根生长的抑制作用大于胚芽。
2.2 Cr(Ⅵ)溶液对小麦生长发育过程中关键酶淀粉酶活性的影响
2.2.1 Cr(Ⅵ)溶液对小麦幼苗生长过程中淀粉酶同工酶活性的影响 如图3所示,不同浓度Cr(Ⅵ)处理7 d 的小麦总淀粉酶同功酶谱带共有6条,依次标为a、b、c、d、e、f。与对照相比,Cr(Ⅵ)处理浓度为3~15 mg/L时,6条同工酶的谱带宽度和亮度均高于对照,且在10 mg/L时的谱带最宽、亮度最强;Cr(Ⅵ)处理浓度为18 mg/L时,6条同工酶谱带宽度、亮度基本和对照相同;Cr(Ⅵ)处理浓度为20~30 mg/L时,a、d、e、f 4条同工酶的表达均受到不同程度的抑制,尤其是处理浓度达到 30 mg/L 时,a、b 2条谱带消失,d、e、f 3条同工酶谱带明显变浅、变窄。结果表明,随着Cr(Ⅵ)浓度的升高,小麦幼苗生长过程中淀粉酶同工酶的活性与表达呈现先增后降特征;总体变化趋势与Cr(Ⅵ)处理后小麦种子胚芽的生长规律基本一致,其中高浓度Cr(Ⅵ)(>18mg/L)处理时对淀粉酶同工酶的抑制作用同于对胚根的抑制作用。总体结果说明,Cr(Ⅵ)处理对小麦生长发育的影响可能是通过影响淀粉酶活性来实现的。
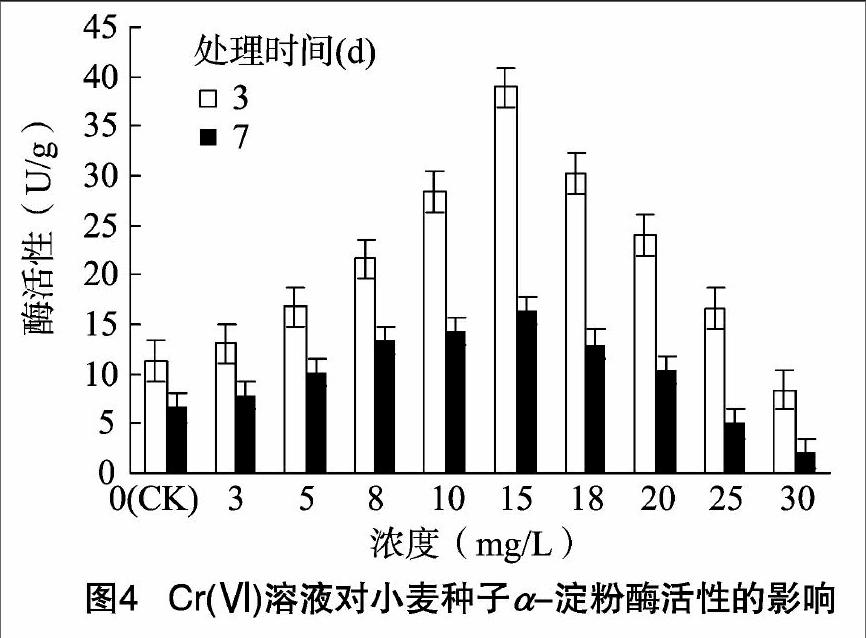
2.2.2 Cr(Ⅵ)溶液对小麦α-淀粉酶活性的影响 α-淀粉酶是植物种子萌发过程中产生的重要酶,因此其活性的高低与种子活力大小、种子是否能够萌发密切相关。如图4所示,Cr(Ⅵ)处理3、7 d的小麦α-淀粉酶活性变化趋势基本一致,都随着Cr(Ⅵ)处理浓度的升高呈先升后降趋势,且活性都在 15 mg/L Cr(Ⅵ) 处理时达到最大值。与对照相比,Cr(Ⅵ)处理3 d 的小麦α-淀粉酶活性在Cr(Ⅵ)处理浓度为30 mg/L时受到抑制,处理浓度为3~25 mg/L均提高了α-淀粉酶活性;Cr(Ⅵ)处理7 d后,小麦幼苗的α-淀粉酶活性在25 mg/L时开始受到抑制,此后继续升高Cr(Ⅵ)处理浓度,α-淀粉酶活性受到更强抑制。这与小麦胚芽生长的受抑制浓度略有差异,但与培养7 d的淀粉酶同工酶垂直电泳检测结果基本一致。
3 结论
试验结果表明,低浓度Cr(Ⅵ)处理对小麦种子的萌发有一定的促进作用,Cr(Ⅵ)浓度低于18 mg/L时,小麦胚芽的芽长以及淀粉酶同工酶活性与对照相比均有所提高;Cr(Ⅵ)浓度高于18 mg/L时,小麦苗芽与淀粉酶同工酶活性都受到了抑制,且随着Cr(Ⅵ)浓度的升高,抑制作用更强。因此可以认为,18 mg/L为临界值,总体表现出“低促高抑”特点。这与李桂玲等在研究Cr3+处理对小麦种子幼苗的影响以及黄辉等研究Cr(Ⅵ)处理玉米幼苗中得到的结果一致[9-10]。此外,高浓度Cr(Ⅵ)处理时,小麦种子胚根生长的受抑制程度大于胚芽[11],且对小麦同种器官在不同生长发育阶段抑制作用不同,后期抑制程度更大。原因可能是种子胚根部位最先突破种皮暴露在Cr(Ⅵ)溶液中,随着胚根受Cr(Ⅵ)胁迫时间延长,积累量越多,因此从宏观上表现出受到更严重的伤害。
植物种子萌发需要足够的物质和能量来源,小麦等禾谷类植物种子的主要贮藏物质是淀粉,通过淀粉酶水解淀粉获得能源物质进而完成种子的萌发,这个过程中淀粉酶活性至关重要[12-13]。本试验结果表明,低浓度(3~15 mg/L) Cr(Ⅵ) 处理时,萌发7 d的小麦淀粉酶同工酶活性逐渐升高,使得淀粉的水解速度加快从而促进小麦种子胚芽的生长;当Cr(Ⅵ)盐浓度大于18 mg/L时,淀粉酶同工酶活性开始降低,相应的小麦种子胚芽的生长受到了抑制,芽长均低于对照;高浓度Cr(Ⅵ)处理时,小麦α-淀粉酶活性也受到了不同程度的抑制,表现为小麦胚根胚芽的生长受到抑制。有学者研究了小麦和水稻种子萌发阶段同工酶的表达,结果证明淀粉酶活性变化趋势反映了实生苗的形态学变化[14-15]。因此,Cr(Ⅵ) 盐对小麦种子萌发的影响可能是通过影响小麦种子中淀粉酶活性来实现的,但是关于Cr(Ⅵ)盐如何作用于淀粉酶活性部位,以及高浓度Cr(Ⅵ)盐如何引起淀粉酶活性下降等问题的分子机理值得进行进一步的研究。
参考文献:
[1]Becquer T,Quantin C,Sicot M,et al. Chromium availability inultramafic soils from New Caledonia[J]. Science of the Total Environment,2003,301(1/2/3):251-261.
[2]张义贤. 三价铬和六价铬对大麦毒害效应的比较[J]. 中国环境科学,1997,17(6):565-568.
[3]Shanker A K,Cervantes C,Loza-Tavera H,et al. Chromium toxicity in plants[J]. Environment International,2005,31(5):739-753.
[4]李 天,来航线,和文祥,等. Cr(Ⅵ)的土壤酶效应研究[J]. 西北农林科技大学学报:自然科学版,2012,40(8):171-178.
[5]马鸿翔,王龙俊,姚金保,等. 江苏小麦品质现状与提升策略[J]. 江苏农业学报,2013,29(3):468-473.
[6]宋 波,高 定,陈同斌,等. 北京市菜地土壤和蔬菜铬含量及其健康风险评估[J]. 环境科学学报,2006,26(10):1707-1715.
[7]李 勤. 谷物萌发前后淀粉酶活力的比较[J]. 广西轻工业,2007,23(5):9-10.
[8]王学奎. 植物生理生化实验原理和技术[M]. 2版. 北京:高等教育出版社,2006:174-176.
[9]李桂玲,李明泽,李欢庆,等. Cr3+对小麦种子萌发及幼苗生长的影响[J]. 江苏农业科学,2009(4):115-116.
[10]黄 辉,高 峡,王 娟. 六价铬对玉米幼苗生长及抗氧化系统的影响[J]. 农业环境科学学报,2011,30(4):633-638.
[11]李丽君,郑普山,周怀平,等. 铬对玉米种子萌发的影响[J]. 山西农业科学,2001,29(2):32-34.
[12]陈禅友,刘 磊. 长豇豆种子萌发进程中生理生化指标动态变化[J]. 种子,2006,25(9):30-33.
[13]何俊瑜,任艳芳. 镉胁迫对水稻种子萌发、幼苗生长和淀粉酶活性的影响[J]. 华北农学报,2008,23(增刊1):131-134.
[14]Umemura T,Perata P,Futsuhara Y,et al. Sugar sensing and alpha-amylase gene repression in rice embryos[J]. Planta,1998,204(4):420-428.
[15]Cejudo F J,Cubo M T,Baulcombe D C. Amyl expression during wheat seed germination[J]. Plant Science,1995,106(2):207-213.
猜你喜欢
遗传(2023年8期)2023-08-25
宁夏医学杂志(2020年3期)2021-01-21
中国粮油学报(2019年4期)2019-07-12
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
湖北农业科学(2015年17期)2015-10-09
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
食品工业科技(2014年7期)2014-03-11
茶叶通讯(2014年2期)2014-02-27
