
庙岛群岛南五岛灌木群落结构及其对环境因子的响应
2015-07-05 07:03:12齐婷婷王晓丽石洪华
西北植物学报 2015年5期
齐婷婷,王晓丽,冯 炘*,石洪华
(1 天津理工大学 环境科学与安全工程学院,天津300384;2 国家海洋局第一海洋研究所,山东青岛266061)
群落结构是植物群落的基本属性,也是认识群落的组成、变化和发展趋势的基础[1]。自然界的植物群落是植物与环境相互作用的产物,群落结构的空间格局分布受环境因素、空间因素的梯度规律影响[2]。研究环境因子与植物群落分布格局作用机理,对深入研究群落生态学规律,解读群落与环境间复杂性关系具有重要意义[3]。植被数量分类和排序能较为简单地揭示植被群落与环境之间的复杂生态关系[4]。目前在国内外的相关研究中,双向指示种分析法(TWINSPAN)是群落生态学使用最广泛的等级分类方法之一,而典范对应分析(CCA)是植被梯度分析与环境解释最为常用的方法。
庙岛群岛南部海岛作为一种特殊的陆地生态系统,生物群落和环境与大陆基本相似,但其生态结构与功能比大陆简单[5-6],林木立地条件差,土壤贫瘠,生态系统脆弱,时常受到大风、寒潮、海岸侵蚀等自然灾害的影响。灌木因其根系发达,枯落物丰富,对海岛地区的水源涵养、水土保持以及生态安全具有重要意义[7-8]。国内外对海岛地区灌木的研究多集中于植被种属的区系分析及物种多样性的研究[9],近年来也有涉及灌木群落的研究但仅限于群丛的划分[10],而对灌木群落分布格局及影响因子的研究鲜有报道。为此,本研究运用TWINSPAN 和CCA 分析方法,对庙岛群岛南部岛群灌木植被进行数量分类与排序,并利用向前引入法识别出影响海岛森林灌木种群格局的关键环境因子,以明确不同生态适应型的植物受环境异质性影响程度的差异,揭示该地区灌木林群落分布规律,以期为海岛地区森林生态保护提供理论基础和依据。
1 研究区概况
庙岛群岛南部岛群位于山东省烟台市长岛县境内,地处渤海海峡黄、渤海交界处,包括南长山岛、北长山岛、庙岛、大黑山岛、小黑山岛,多为山坡和平地,地理坐标为37°53.690′~37°59.517′N、120°35.942′~120°45.470′E。调查区域土地总面积为32.81km2,海拔3~165m,自然坡度为0~67°。该地区属暖温带季风大陆性气候区,年平均温度为11.9 ℃。历年年平均降水量为555.5mm,59%的降水集中在夏季,年日照时数2 542h。土壤主要分为棕壤、褐土和潮土3类,棕壤土主要分布在岛屿中上部,褐土主要分布在岛屿中下部和部分滨海平缓地,还有少面积潮土,主要分布于滨海平缓地[11]。
2 研究方法
2.1 野外调查
2012年7月根据庙岛群岛南五岛植被分布特征及面积的大小,依据代表性和可达性原则,在各岛分别设置1~3 条100 m 样带,相邻样带间隔300 m,样带方向根据各海岛地形设定,平均每隔50 m设置5m×5m 的调查样地,共设置44个样地(图1)。样地调查内容包括:乔木层盖度、灌木层出现的每一物种的植物名称、多度、盖度、高度、胸径。同时在每个样方中,样方的正中心使用罗盘测定样方的坡度、坡向,使用全球定位系统(GPS)测定经纬度、海拔高度。

图1 庙岛群岛南五岛灌木调查样方分布Fig.1 Distribution of investigative plots in southern five islands of Miaodao Archipelago

表1 坡向的等级划分Table 1 Rank bipartition of aspect
为便于建立环境数据矩阵,将坡向和坡度数据以等级制表示。将坡向按照45°的夹角,以北为0°,顺时针方向旋转分为8个坡向等级,以数字表示各等级(表1)。数字越大,表示坡向越向阳[12-13]。采用等差坡度分级方法,将坡度按照极差5依次分级,所调查区域的坡度范围为3°~67°,共将坡度划为14个等级[14]。
2.2 土壤样品采集及分析
在每个样地分别选取3个取样点,多点表层土(0~30cm)均匀混合取得样方土样,带回实验室后自然风干,除去草根、大石砾等杂质,过2mm 钢筛,磨细待测。土壤理化性质按鲁如坤编写的《土壤农业化学分析方法》进行[15]。
2.3 数据处理
重要值计算[16]:草本层重要值=(相对频度+相对盖度)/2
群落划分采用双向指示种分析法(TWINSPAN)在WinTWINS软件包上进行;环境因子对样方的排序采用典型对应分析(canonical correspon-dence analysis,CCA),在Canoco for Windows 4.5软件包上进行。
样方44因石块较多,故没有进行土壤养分测定,灌木排序时剔除样方44,剔除样方中重要值小于5的偶见种,整理后形成43×20的重要值矩阵,便于CCA 排序分析。分析前,灌木物种数据量和环境因子(除pH 外)均转换成ln(x+1)形式。首先对灌木植物重要值进行除趋势对应分析(detrended correspondence analysis,DCA),以确定其属于单峰型分布或线型分布[17]。DCA 结果表明,所有轴中梯度最长的为4.657,大于3,因此运用典型对应分析(CCA)进行海岛灌木植被与环境因子之间的关系分析。同时采用向前引入法逐步筛选出显著的环境变量,每一步都采用Monte Carlo置换检验。
3 结 果
3.1 南五岛森林灌木
庙岛群岛南五岛的44个样方中共发现灌木27种,隶属于15科23属(表2);其中种数最多的科是豆科,包含6个种;其次是桑科,包含4个种。该调查样地灌木的种属区系组成以泛热带、北温带成分为主,如灌木中的荆条、酸枣、小叶朴、雀儿舌头、花椒、迎春属于泛热带植被,榔榆、麻栎、蒙古栎、桑、紫荆等属于北温带植被[18]。
从表2可以看出,在整个研究区域内,荆条频度最高达66.67%,是灌木群落中的建群种;其次是酸枣和扁担木,各自出现的频度也都达50%以上;柘树的频度为31.11%,分布在除北长山岛以外的其余四岛;其余灌木的频度都小于20%(表2)。

表2 庙岛群岛南五岛灌木种类的组成Table 2 Composition of shrub species in southern five islands of Miaodao Archipelago
3.2 群落结构
运用TWINSPAN 对群落灌木层的主要优势种进行了类型划分,结果表明44个样地可以划分为7组(图2),其中S2、S5、S17样方包含的灌木群落种类型与样方S11、S24、S27、S29、S34包含的灌木群落种相接近,结合实际生态意义,依据《中国植被》的分类和命名原则[19],最终将灌木划分为6 个群丛,各群丛主要特征见表3。
(Ⅰ)桑群丛,包括样地S12。位于南长山岛海拔46 m 处的刺槐林,坡向为阳坡,接受光照时间长,含水量较高,水热条件良好,群落总盖度为100%,乔木生长茂盛,但林下灌木分布稀疏,只分布有桑,为桑单优势群丛,该群落土壤属弱碱性土壤,土壤贫瘠,有效养分含量低。
(Ⅱ)刺槐-柘树群丛,包括样地S18、S26、S31、S38。位于13~40m 的中低海拔处,坡度0°~67°,坡向为半阳坡或半阴坡,主要分布在黑松-刺槐混交林下或黑松林下,群落总盖度为95%~100%,灌木盖度仅4%,灌木植被零星分布,主要以刺槐幼苗为优势种,柘树为次优势种。
(Ⅲ)酸枣群丛,包括样地S2、S5、S11、S17、S24、S27、S29、S34。位于23~68m 处的中低海拔处,坡度3°~24°,多数样方位于缓地和斜坡处,坡向主要为半阴坡,位于黑松或混交林下,土壤养分含量高,含水量高,利于林下植被的生长,灌木盖度为45%,以酸枣为优势种,另有柘树、扁担木、荆条等灌木植被生长,灌木种相对丰富。
(Ⅳ)扁担木群丛,为灌木优势群丛,包括样地S3、S4、S6~S10、S15、S16、S20~S23、S28、S30、S35、S42、S44。样方分布的海拔范围为37~142m,坡度3°~51°,坡向主要为半阳坡。在黑松、刺槐和混交林下均有分布,群落总盖度为65%~98%,土壤呈酸性且有机质、有效氮的含量高,灌木低分布广泛,高海拔地区生态环境脆弱,植被分布少。扁担木为优势种,伴生种为荆条、酸枣、柘树、紫穗槐等。
(Ⅴ)荆条群丛,包括样地S1、S13、S14、S19、S32、S33、S36、S37、S39~S41、S43。位于海拔20~165m 处,坡度14°~65°,坡向为阳坡、半阳坡或半阴坡。主要分布在黑松林下,土壤呈酸性,群落总盖度为52%~100%,灌木层盖度最大,荆条为群丛优势种,伴生种有桑、扁担木、刺槐等。
(Ⅵ)紫穗槐群丛,包括样地S25。位于海拔6 m 的平地处,坡向为阳坡且无乔木生长,近岸土壤常年受海水冲刷,土壤pH 值最大、有机质含量低、含水量低,适合阳性耐旱类植被生长,灌木层盖度30%,以紫穗槐为单一优势种。
3.3 灌木植被与环境因子之间的关系
群落的CCA 排序反映了环境对群落类型和物种分布的影响。表4列出的是灌木层排序中各排序轴(AX)的特征值以及排序轴与环境变量的相关系数。其中排序轴特征值反映的是相应的排序轴所集中的物种多度距阵中的信息量的大小。4个轴的特征值分别为0.535、0.286、0.176、0.158,由于第1轴(AX1)的特征值最大(0.535),第2轴(AX2)次之(0.286),前两轴包含的生态信息较大,显示出重要的生态意义,因此,本研究只对第AX1、AX2轴进行深入分析。
利用向前引入法对环境因子进行逐步筛选,Monte Carlo置换检验结果显示,选取的8 个环境因子中,只有乔木层盖度(F=3.19,P=0.004),pH(F=2.01,P=0.014)达到显著水平,表明这两个环境因子对庙岛群岛南五岛灌木植被分布起到了主导作用。

图2 庙岛群岛南五岛灌木层TWINSPAN 分类树状图D.分级水平;N.样地数量;1~44.样地号;Ⅰ~Ⅵ.群丛类型Fig.2 Dendrogram of the TWINSPAN classification in shrub layer of southern five islands of Miaodao Archipelago D.Hierarchical level;N.The number of samples;1~44.Plot code;Ⅰ~Ⅵ.Association types

表3 6个群丛的环境变量值Table 3 Mean of the environmental variables between species groups in the study area

表4 CCA排序各环境因子与前4个排序轴的相关系数Table 4 Correlations of environmental variables with the first four axes of CCA as well as their eigenvalues
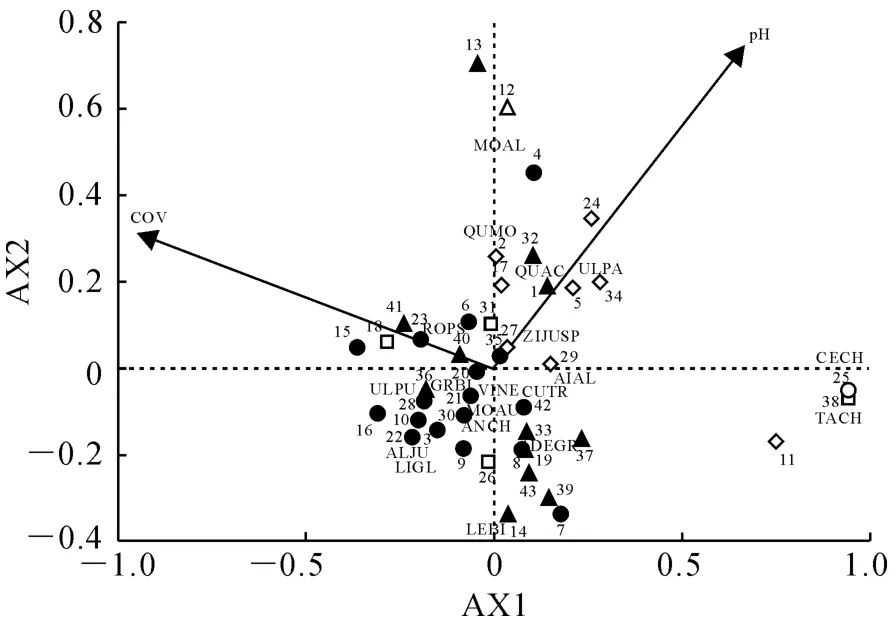
图3 南五岛灌木植被与环境因子CCA 排序图数字代表样方号;字母为灌木代码,同表2;+.桑群丛;□.刺槐-柘树群丛;◇.酸枣群丛;●.扁担木群丛;▲.荆条群丛;○.紫穗槐群丛;pH.土壤酸碱度;COV.乔木层盖度Fig.3 CCA ordination of environment variables and shrub vegetation in southern five islands of Miaodao Archipelago The number is the plot;Letter is the code of shrub in Table 2;+.Morus alba population;□.Robinia pseudoacacia-Cudrania tricuspidata population;◇.Ziziphus jujuba var.spinosapopulation;●.Grewia biloba var.parviflora population;▲.Vitex negundo var.heterophyllapopulation;○.Amorpha fruticosa population;pH.Hydrogen ion concentration;COV.Coverage of tree
庙岛群岛南五岛森林灌木植被与环境因子之间的关系可以很好地在CCA 排序图中表现出来(图3)。第一排序轴(AX1)与乔木层盖度呈显著负相关,代表林下光照的梯度变化。沿AX1 轴从左到右,随着乔木层盖度由大到小,即林下光照条件由弱到强,灌木植被的种类与数量相应增加;第一排序轴与土壤pH 呈显著正相关,代表土壤酸碱度的梯度变化。沿AX1轴从左到右,即土壤由酸性土壤过渡到碱性土壤,灌木植被的分布逐渐减少。
4 结论与讨论
4.1 南五岛灌木种类及区系
研究发现,因调查区域的坐标和生境的差异,陆域系统灌木的种类有明显的差异,少则几种,多达上百种。庙岛群岛南五岛有灌木植被27 种,隶属于15科23属,以豆科、桑科为主,优势种为荆条、酸枣、扁担木。与同属温带大陆季风气候的太岳山(28种)[20]、北京八达岭地区(25种)[21]所含灌木种数相当,这是因为岛陆部分是海岛陆地生态系统的主体,具有陆域资源特征,生物群落和环境与大陆基本相似,加之对灌木群落的人为干扰较小,导致海岛上有不少灌木植被生长。但与太子山半干旱地区(125种)[22]相比,该地区灌木植被较少。岛陆面积相对较小,生态系统构成较为单一,加之受台风、风暴潮、干旱等频发性的自然灾害的影响及海岛土壤贫瘠等原因,岛陆生态系统稳定性差,生态特征脆弱[23-25],这些条件造成了海岛陆地灌木植被种类相对匮乏,物种多样性较低。灌木的种属区系组成以北温带、泛热带成分为主[18]。导致这种现象的原因可能是庙岛群岛位于东亚暖温带,属暖温带东亚季风大陆性气候,这种区系特点与其环境条件是相一致的;另一方面,种属区系属泛热带的灌木植被在海岛分布广泛,说明该区系具有一定的热带亲缘。
4.2 灌木植被与环境因子的关系
灌木被划分为6个群丛,分别为:酸枣群丛、扁担木群丛、荆条群丛、紫穗槐群丛、桑群丛和刺槐-柘树群丛。不同生态适应型的灌木受环境异质性的影响有所差异,庙岛群岛南五岛灌木的空间分布主要受乔木层盖度与土壤pH 两个环境因子的影响。
林下植被的生长不仅与土壤性质、地形有关,而且还受生物因子(林冠郁闭度)的影响[26]。在海岛地区,乔木层盖度在灌木物种分布格局的形成中明显起着主导作用。乔木层盖度即成树树冠的遮蔽程度反映了林下光照的变化状况[27],能够影响林下气温、湿度、土壤温度等小气候因子,说明在海岛地区光照是影响灌木群落分布的主要因素,过高或过低都会影响森林植物的生长发育。由于对光照和降水的阻挡作用,乔木层盖度与林下灌木呈显著负相关(r=-0.675 4),从图3可以看出,沿AX1轴从左到右,乔木层盖度由大到小,光照由弱到强,适宜于空气湿度较大、土壤湿润的扁担木、刺槐-柘树群丛位于排序图左部,喜温暖干燥的酸枣、桑、紫穗槐群丛位于排序图右侧,荆条群丛生境范围广,位于排序图中间位置。多数灌木物种集中分布在排序图的中间部位,即光照条件适中、温暖而湿润的生境下。这是因为乔木盖度大的群落,林下光照条件差,林内环境荫蔽湿润,不利于植物的光合作用进行,植被生长缓慢;相反乔木盖度小的群落,林下光线充足,生境干燥,植物蒸腾作用加快,植被容易枯萎。此外,不同灌木植被对光照条件的响应也与它们各自的生活习性相关。刺槐(AMFR)、榆(ULPU)等中性耐荫、喜土壤湿润的灌木分布在乔木盖度相对大的环境;酸枣(ZIJUSP)、臭椿(AIAL)等耐旱阳性物种分布在乔木盖度小的环境;紫荆(CECH)、柽树(TACH)因不耐遮荫,要求光线充足,分布在远离其他植被AX1轴的最右端,即无乔木分布太阳直射的环境下。本结论与Aarrestad等研究的地面层植被的分布受乔木层盖度影响较大[28]的结论相一致。
植被的分布与生长发育往往受土壤理化性质的影响[29-30],贺强等研究了黄河口盐沼植被分布与土壤pH 呈显著负相关[31]。作为衡量土壤酸碱强度的指标,土壤pH 在本研究中显著影响灌木植被的分布。沿AX1 轴从左到右,即土壤pH 值由低到高,灌木植被逐渐减少,灌木植被主要分布在pH 较低的弱酸性土壤(图3),如扁担木群丛、荆条群丛、刺槐-柘树群丛。这是因为在酸性条件下,土壤微生物种类和数量少,微生物活性低,有机质分解缓慢,有机质含量相对较高,利于灌木植被生长。岛上陆地四面临海,由于海风、海雾、海浪冲刷等原因,部分土壤呈现盐碱化,土壤pH 值升高有利于微生物活动,加速有机质分解;此外,碱解氮是衡量土壤肥力重要指标之一,有研究表明土壤碱解氮含量与有机质含量关系密切,pH 值升高时有机质含量降低导致碱解氮减少[32],土壤肥力降低,土壤板结、通气性、透水性差,继而限制植被的生长发育[33],pH 值较高的区域只有少量耐盐碱灌木生长,如紫荆、柽树位于排序图pH 值高的最右侧。因此,在海岛陆地森林应大力种植耐盐碱的植被以抵制土壤盐碱化的威胁。本研究与邵方丽的研究结果相似,她发现在冀北山典型森林中,pH 值为5.9~6.1的弱酸性土壤适于灌木的生长[34]。
另外,灌木群丛中的优势种的分布格局与群丛类型的分布格局有很大的相似性,且优势种均位于对应群丛的中心位置,如以扁担木(GRBI)为优势种的群丛与扁担木均位于CCA 排序图的左下方;同样以酸枣(ZIJUSP)为优势种的群丛与酸枣均位于排序图的右上方。由此可以反映出植物群丛中优势种的分布格局在很大程度上决定着植物群丛类型的分布格局。由灌木排序图(图3)与分类图(图2)可知,CCA 二维排序结果与TWINSPAN 分类结果较为一致,这表明庙岛群岛南五岛群灌木群落在空间地理分布上具有一定的规律性,酸枣、扁担木、荆条优势群丛位于光照条件适中、温暖而湿润的弱酸性土壤下,碱性土壤生长的灌木植被种类单一,在海岛地区应大力种植抗盐碱的灌木植被。
[1] SHI Z M(史作民),CHENG R M(程瑞梅),LIU SH R(刘世荣),et al.Study on species diversity of plant communities in Bao Tianman[J].Scientia Silvae Sinicae(林业科学),2002,38(60):17-23(in Chinese).
[2] BORCARD D,LEGENDRE P,DRAPEAU P.Partialling out the spatial component of ecological variation[J].Ecology,1992,73:1 045-1 055.
[3] ZHANG W H(张文辉),LU T(卢 涛),MA K M(马克明),et al.Analysis on the environmental and spatial factors for plant community distribution in the arid valley in the upper reach of Minjiang River[J].Acta Ecologica Sinica(生态学报),2004,24(3):552-559(in Chinese).
[4] 宋永昌.植被生态学[M].上海:华东师范大学出版社,2001.
[5] LAGERSTROM A,NILSSON M C,WARDLE D A.Decoupled responses of tree and shrub leaf and litter trait values to ecosystem retrogression across an island area gradient[J].Plant and Soil,2013,367(1-2):183-197.
[6] NOGUE S,NASCIMENTO D L,FERNANDEZ-PALACIOS J M,et al.The ancient forests of La Gomera,Canary Islands,and their sensitivity to environmental change[J].Journal of Ecology,2013,101(2):368-377.
[7] ZHANG Z S,LI X R,ROBERT S,et al.Effect of sand-stabilizing shrubs on soil respiration in a temperate desert[J].Plant and Soil,20133671-2449-463.
[8] YANG Z Y,WANG M G,WANG B W,et al.Experimental research on shrub runoff-plot[J].Research of Soil and Water Conservation,2001,8(9):28-30.
[9] 张 骞.庙岛群岛植物群落结构研究[D].济南:山东师范大学,2008.
[10] XU H CH(许鸿川).Classification of subtropical shrub communities of coastal islands in Fujian Province[J].Journal of Fujian Agricultural University(福建农业大学学报),1999,28(1):121-124(in Chinese).
[11] 《中国海岛志》编纂委员会.中国海岛志[M].北京:海洋出版社,2013:79-81.
[12] ZHANG J T,ZHANG F.Diversity and composition of plant functional groups in mountain forests of the Lishan Nature Reserve,North China[J].Botanical Studies,2007,48(3):339-348.
[13] ZHANG W Q(章皖秋),LI X H(李先华),LUO Q ZH(罗庆州).Spatial distribution of vegetation in Tianmu Mountain Nature Reserve based on RS and GIS data[J].Chinese Journal of Ecology(生态学杂志),2003,22(6):21-27(in Chinese).
[14] TANG G A(汤国安),SONG J(宋 佳).Comparison of slope classification methods in slope mapping from DEMs[J].Journal of Soil
and Water Conservation(水土保持学报),2006,22(2):157-160(in Chinese).[15] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999.
[16] ZHANG Y(张 勇),HU H B(胡海波),HUANG J(黄 进).Plant diversity and its relationship with environment in planning afforestation of Yuntai Mountain in Lianyungang[J].Subtropical Plant Science(亚热带植物科学),2009,38(1):41-45(in Chinese).
[17] LEP J,MILAUER P.Multivariate Analysis of Ecological Data using CANOCO[M].Cambridge:Cambridge University Press,2003:1-269.
[18] WU ZH Y(吴征镒).The areal-types of Chinese genera of seed plants[J].Acta Bot.Yunnan.(云南植物研究),1991,(S IV):1-139(in Chinese).
[19] 中国植被编辑委员会.中国植被[M].北京:科学出版社,1983:143-156.
[20] NAN H L(南海龙),HAN H R(韩海荣),MA Q Y(马钦彦).Species diversity of herb and shrub layers in gaps of a mixed conifer broadleaved forest in Taiyue Mountain of Shanxi Province[J].Journal of Beijing Forestry University(北京林业大学学报),2004,28(2):52-56(in Chinese).
[21] DU L X(杜丽侠),XING SH H(邢韶华).Relationship between spatial distribution pattern of shrub community and environmental factors in Badaling of Beijing[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2004,29(3):601-607(in Chinese).
[22] LIU X J(刘晓娟),SUN X G(孙学刚),DONG L X(董力行).Studies on species diversity of vascular plants in the Taizi Mountains forest region[J].Journal of Gansu Agricultural University(甘肃农业大学学报),2004,39(5):548-553(in Chinese).
[23] BUSTAMANTE-SANCHEZ M A,ARMESTO J J.Seed limitation during early forest succession in a rural landscape on Chiloe Island,Chile:implications for temperate forest restoration[J].Journal of Applied Ecology,2012,49(5):1 103-1 112.
[24] KATOVAI E,BURLEY A L,MAYFIELD M M.Understory plant species and functional diversity in the degraded wet tropical forests of Kolombangara Island,Solomon Islands[J].Biological Conservation,2012,145(1):214-224.
[25] INAGAKI Y,KURAMOTO S,FUKATA H.Effects of typhoons on leaf fall in hinoki cypress(Chamaecyparis obtusa Endlicher)plantations in Shikoku Island[J].Bulletin of the Forestry and Forest Products Research Institute,2010,9(3):103-112.
[26] BARBIER S,GOSSELIN F,BALANDIER P.Inflluence of tree species on understory vegetation diversity and mechanism involved:A critical review for temperate and boreal forests[J].Forest Ecology and Management,2008,254:1-15.
[27] FU CH J(符婵娟),LIU Y H(刘艳红),ZHAO B Y(赵本元).Regeneration characteristics and influencing factors of Abies fargesii forests in Shennongjia National Nature Reserve[J].Acta Ecologica Sinica(生态学报),2009,29(8):4 179-4 186(in Chinese).
[28] AARRESTAD P A,MASUNGA G S,HYTTEBORN H,et al.Infuence of soil,tree cover and large herbivores on field layer vegetation along a savanna landscape gradient in Northern Botswana[J].Journal of Arid Environments,2011,75:290-297.
[29] REN G H,DENG B,SHANG Z H,et al.Plant communities and soil variations along a successional gradient in an alpine wetland on the Qinghai-Tibetan Plateau[J].Ecological Engineering,2013,61:110-116.
[30] SOLON J,DEGORSKI M,ROO-ZIELINSKA E.Vegetation response to atopographical-soil gradient[J].Catena,2007,71:309-320.
[31] HE Q(贺 强),CUI B SH(崔宝山),ZHAO X SH(赵欣胜).Relationships between saltmarsh vegetation distribution/diversity and soil chemical factors in the Yellow River Estuary,China[J].Acta Ecologica Sinica(生态学报),2009,29(2):666-687(in Chinese).
[32] JIA K L T(甲卡拉铁),LI G ZH(李桂珍),RAO M Y(尧美英).Correlation between soil pH and the contents of available nutrients in selected soils from mango orchards in Panxi[J].South China Fruits(中国南方果树),2011,40(4):8-12(in Chinese).
[33] TANG K(唐 琨),ZHU W W(朱伟文),ZHOU W X(周文新).Research progress on effects of soil pH on plant growth and development[J].Crop Research(作物研究),2013,27(2):207-210(in Chinese).
[34] 邵方丽.冀北山地典型森林植被与土壤成分的空间异质性关系研究[D].北京:北京林业大学,2012.
猜你喜欢
中国水土保持科学(2023年6期)2024-01-05 05:53:12
水土保持研究(2022年1期)2022-12-27 07:35:24
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
林业调查规划(2020年3期)2020-06-03 07:02:36
乡村科技(2018年8期)2018-02-13 12:16:18
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
山西大学学报(自然科学版)(2014年2期)2014-05-10 06:55:16
