
柑橘全爪螨对甲氰菊酯和阿维菌素的抗性选育及交互抗性
2015-07-02 01:45何恒果王进军
植物保护 2015年6期
何恒果, 王进军
(1.西华师范大学生命科学学院,南充 637002;2.西南大学植物保护学院,重庆 400715)
柑橘全爪螨对甲氰菊酯和阿维菌素的抗性选育及交互抗性
何恒果1, 王进军2
(1.西华师范大学生命科学学院,南充 637002;2.西南大学植物保护学院,重庆 400715)
通过室内抗性品系选育,研究了柑橘全爪螨对甲氰菊酯和阿维菌素的抗性发展情况,并就其与柑橘园常用11种杀螨剂的交互抗性进行了分析。结果表明:在柑橘全爪螨19代中用甲氰菊酯和阿维菌素分别不连续汰选16次和11次后,柑橘全爪螨对两者的抗性分别为29.92和3.80倍;甲氰菊酯抗性品系(FeR)对哒螨灵、三氯杀螨醇和三唑锡产生了明显的交互抗性,阿维菌素抗性品系(AbR)对甲维盐产生了明显的交互抗性。试验结果可为柑橘全爪螨抗性治理提供参考。
柑橘全爪螨; 甲氰菊酯; 阿维菌素; 交互抗性
柑橘全爪螨[Panonychus citri(McGregor)],又称柑橘红蜘蛛,是一种世界性的柑橘害螨,除为害柑橘外,还为害梨、苹果、花椒、苦楝、桂花等多种经济作物和观赏植物[1]。由于螨体小、世代多、繁殖速度快、发育历期短的特点,极易对药剂产生抗性。已有的研究结果表明柑橘全爪螨对许多杀虫(螨)剂均产生了一定的抗性[25]。
甲氰菊酯(fenpropathrin)和阿维菌素(abamectin)虽然曾有一些抗药性产生的报道[6],但本实验室通过在农药市场和田间调查发现:甲氰菊酯是一种虫螨兼治、触杀性强、速效性好、杀虫谱广的拟除虫菊酯类杀虫、杀螨剂,而且对寄主植物一般不造成药害[7];阿维菌素则是具有高选择性高效生物源杀虫、杀螨、杀线虫剂[8],两种药剂现在仍然在柑橘园内广泛使用[9-10]。为了明确在新的环境下,柑橘全爪螨对这两种杀螨剂的抗性发展变化,以及与其他常用农药是否具有交互抗性,本研究通过甲氰菊酯和阿维菌素对柑橘全爪螨进行室内抗性选育,分析了抗性发生发展趋势,并就11种杀螨剂的交互抗性进行了研究。旨在为合理使用甲氰菊酯和阿维菌素、延长其使用寿命和开展其预防性抗性治理提供依据。
1 材料与方法
1.1 供试药剂
甲氰菊酯(fenpropathrin,92%原药),阿维菌素(abamectin,93%原药),哒螨灵(pyridaben,95.2%原药),辛硫磷(phoxim,90%原药),马拉硫磷(malathion,89.1%原药),甲维盐(全称甲氨基阿维菌素苯甲酸盐,emamectin benzoate,49.5%乳油),炔螨特(propargite,40%乳油),三氯杀螨醇(dicofol, 20%乳油),双甲脒(amitraz,20%乳油),三唑锡(azocyclotin,25%可湿性粉剂),毒死蜱(chlorpyrifos,99.9%原药)。上述药剂除毒死蜱为Sigma公司购得外,其余均由四川省药检所提供。
1.2 抗性筛选和生物测定
1.2.1 试验种群
柑橘全爪螨采自重庆北碚歇马中国柑橘研究所柑橘苗圃园枳壳苗上,该园从未用药。将采集的柑橘全爪螨移接到盆栽枳壳苗上,在温度为(26±1)℃, RH为70%~75%的人工气候室内饲养。将这一室内种群分为3个部分,其中一部分不喷药,作为敏感品系SS;另2部分分别用甲氰菊酯、阿维菌素单一处理。甲氰菊酯抗性品系(FeR)19代中不连续汰选16次获得(第1代、5代、10代未作处理),阿维菌素抗性品系(AbR)19代中不连续汰选11次获得(第1代、5代、9代、11代、13代、14代、16代、17代未作处理)。
1.2.2 抗性培育和生物测定
从敏感品系开始培育柑橘全爪螨的抗药性品系。用小型喷雾器喷施甲氰菊酯和阿维菌素药液,分别处理淘汰掉50%~70%的螨量。存活个体继续饲养,当种群数量达到足够用药时再次进行处理,每隔几代测定1次LC50值。
柑橘全爪螨生物测定采用FAO推荐的玻片浸渍法[11];为模仿田间试验,交互抗性测定则采用叶片浸渍法[12]。生测时将原药或药液先用丙酮溶解,再改用蒸馏水稀释成所用药液浓度,最终所用药液中丙酮含量为0.20%。生测中所选个体均为3~5日龄的健康雌成螨,每一浓度设3个重复,另以丙酮水溶液(0.20%)为对照,对照组死亡率在10%以内为有效试验,所得数据在SPSS 12.0 for Windows软件上进行处理,求出毒力回归直线方程和致死中浓度LC50。将每次测得的LC50进行比较,计算得出相对抗性倍数R,R=抗性品系LC50/敏感品系LC50。
2 结果与分析
2.1 柑橘全爪螨对两种杀螨剂的抗性选育
由表1可知,经19代共16次用药筛选,柑橘全爪螨雌成螨对甲氰菊酯的LC50由选育前的4.187 1 mg/L提高至125.265 3 mg/L,抗性增长了29.92倍。从抗性发展过程来看,选育11代以后,逐渐形成柑橘全爪螨对甲氰菊酯的抗性品系。其中选育初期抗性上升较为缓慢,从F0~F4代抗性仅增加了1.79倍;在F4~F8代选育过程中,抗性发展速度比F0~F4代迅速,抗性增长了5.24倍;从F8~F11代,抗性发展速度也较快,但与F4~F8代比较无显著性差异;从F11代开始,抗性发展极为迅速, LC50由F11代的57.491 3 mg/L至F18代的125.265 3 mg/L,抗性增加了16.18倍,差异非常显著。但是,在整个筛选期间柑橘全爪螨对阿维菌素的抗性发展都较为缓慢,经19代共11次筛选,柑橘全爪螨对阿维菌素的敏感度由选育前的0.022 1 mg/L提高至0.083 9 mg/L,抗性仅增长3.80倍(见表1)。由此可见,柑橘全爪螨对甲氰菊酯抗性增长速度远远大于阿维菌素(如在相同筛选次数第9次时,甲氰菊酯抗性品系已经形成,抗性倍数为13.74倍,而阿维菌素筛选品系抗性倍数仅为2.29倍,见表1)。
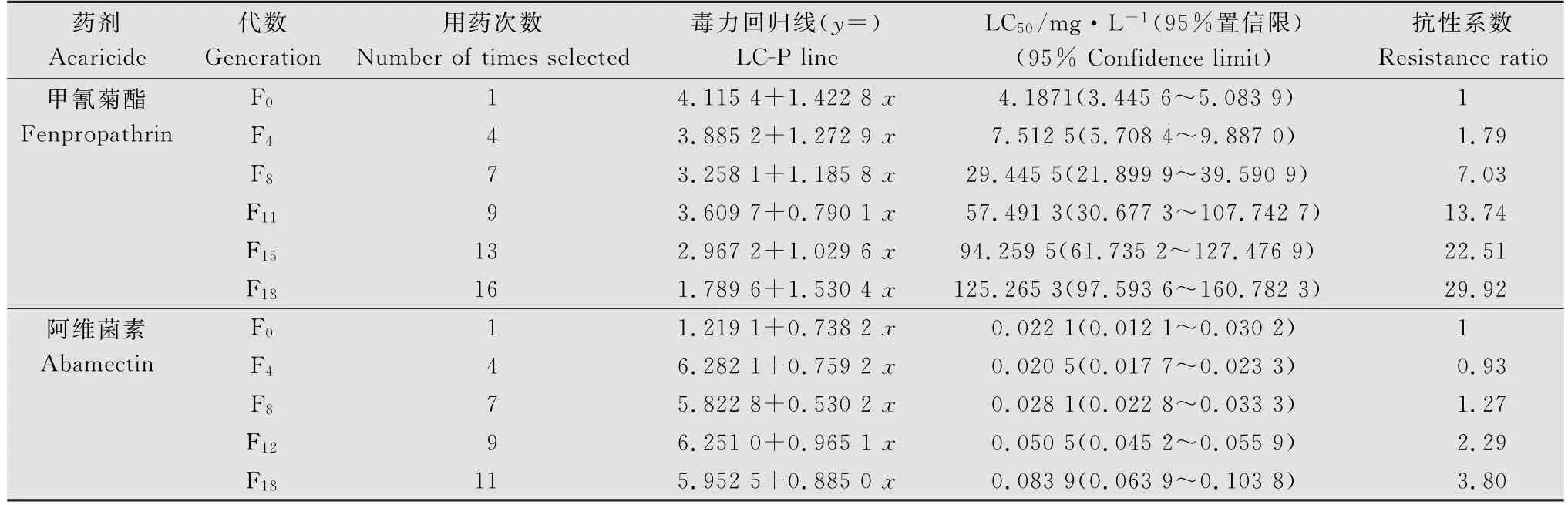
表1 两种杀螨剂对柑橘全爪螨北碚种群的抗性筛选Table 1 Selection of resistance to fenpropathrin and abamectin in Panonychus citri from Beibei,Chongqing
2.2 柑橘全爪螨抗性品系对常用杀螨剂的交互抗性
通过叶片浸渍法测定了柑橘全爪螨敏感品系、甲氰菊酯抗性品系FeR和阿维菌素抗性品系Ab R对11种杀螨剂的毒力(表2)。测得柑橘全爪螨甲氰菊酯抗性品系对甲氰菊酯的抗性为31.14倍,对哒螨灵、三氯杀螨醇和三唑锡分别产生了8.45、6.73和3.37倍的交互抗性,但对阿维菌素、甲维盐和炔螨特无明显交互抗性,而对辛硫磷、马拉硫磷、双甲脒和毒死蜱具有一定的负交互抗性(见表2)。测得阿维菌素抗性品系的抗性为4.37倍,对甲维盐和哒螨灵分别产生了3.32和1.73倍的交互抗性,对甲氰菊酯、辛硫磷、马拉硫磷、三氯杀螨醇、双甲脒、三唑锡和毒死蜱均无明显交互抗性;而对炔螨特有明显的负交互抗性(表2)。

表2 11种杀螨剂对柑橘全爪螨甲氰菊酯抗性品系、阿维菌素抗性品系的毒力Table 2 The toxicity of 11 acaricides to FeR and AbR
3 讨论
试验结果表明,柑橘全爪螨北碚种群能较快地对甲氰菊酯产生抗性,而对阿维菌素抗性增长较为缓慢,这与孟和生等[1314]以山东种群试验得出的结论基本一致,表明来自不同地理种群的柑橘全爪螨的抗性发展基本相同,且在新环境下的柑橘全爪螨抗性发展也与以往基本相似。在试验中,虽然人为条件控制相同,两种药剂的抗性筛选力度也相同(杀死率60%左右),但是由于甲氰菊酯对柑橘全爪螨的刺激增殖作用比阿维菌素更强(待发表),用甲氰菊酯后种群增长加快,用阿维菌素处理后种群发展较为缓慢,由于种群数量达到一定时才可用药筛选,因此使得在相同时间内甲氰菊酯用药次数多于阿维菌素(19代中用甲氰菊酯汰选了16次,用阿维菌素汰选了11次)。以往有研究表明,昆虫或蜱螨抗药性的产生,除了与体内相关酶系有关外,与药剂对其生殖力的影响密切相关[15],这可能也是柑橘全爪螨对甲氰菊酯比对阿维菌素抗性产生更为迅速的原因之一。柑橘全爪螨对拟除虫菊酯类药剂的抗性发展速度非常快,这与其他叶螨如朱砂叶螨[1617]、二斑叶螨[17]抗性研究结果相同。在其他昆虫的研究上也有类似的结果,如用氯菊酯及苯醚菊酯选择德国蜚蠊对拟除虫菊酯的抗性时,发现抗性产生快,用上述任何一种药剂进行选择,敏感品系在6~7代内就可产生抗性[19]。拟除虫菊酯类杀虫(螨)剂具有高效、广谱的特点,可用于多种害虫(螨)的防治,但害虫(螨)对这类药剂抗性产生快,交互抗性谱广,加之该类药剂对害螨的亚致死效应等问题在很大程度上阻碍了这类药剂的发展。
阿维菌素是生物源农药,抗性发展较为缓慢,但单一长期用药,害虫(螨)仍可对其产生抗性[16]。从柑橘全爪螨对其他农药的抗性研究可知,用哒螨灵室内选育12代后抗性达到23.67倍[20],对噻螨酮室内室外抗性选育,其抗性发展极为迅速,室内选育2年后抗性达到4 450倍[2122],辛硫磷选育15次后抗性为18.6倍[23]。总体来看,柑橘全爪螨对噻螨酮的抗性发展最为迅速,其次是哒螨灵,对甲氰菊酯和辛硫磷的抗性发展也较快,只有阿维菌素作为生物源农药其抗药性的产生最为缓慢,因此,提倡生物源农药的使用非常必要。
本试验交互抗性的测定结果表明,柑橘全爪螨甲氰菊酯抗性品系对哒螨灵、三氯杀螨醇和三唑锡产生了明显的交互抗性,对阿维菌素、甲维盐和炔螨特无明显交互抗性,而对辛硫磷、马拉硫磷、双甲脒和毒死蜱具有一定的负交互抗性。为此,在对柑橘全爪螨甲氰菊酯的抗性治理上,要尽可能避免和哒螨灵、三氯杀螨醇和三唑锡混用或轮用,但可以与双甲脒、毒死蜱、马拉硫磷和辛硫磷等轮用或混用。孟和生[12]认为,甲氰菊酯应避免与拟除虫菊酯混用或轮用,可以与水胺硫磷等混用或轮用,这也是可借鉴的地方。阿维菌素抗性品系虽然只有4.37倍的抗性,但对甲维盐已经产生了明显的交互抗性,对哒螨灵也产生了一定的交互抗性,而对炔螨特有明显的负交互抗性。因此,在大田应用中为防止柑橘全爪螨对阿维菌素产生抗性,除了不能长期单一施用阿维菌素外,还要尽量避免和甲维盐及哒螨灵混用或轮用。
柑橘全爪螨由于本身的生物学特性,对农药抗性产生较为迅速,为此,在大田生产实践中,如何有效预防其抗性的快速产生尤为重要。室内结果虽然并不能直接用于田间,但仍然可以指导我们在田间防治过程中及时进行抗性监测,以制定相应的预防性抗性治理策略。
[1] 江西大学.中国农业螨类[M].上海:上海科学技术出版社, 1984:121-122.
[2] 冉春,陈洋,袁明龙,等.桔全爪螨田间种群对杀螨剂的敏感性[J].植物保护学报,2008,35(6):537-540.
[3] 陈洋,冉春,熊琳,等.不同柑桔种质资源对桔全爪螨酶活性及其对药剂敏感性的影响[J].西南农业学报,2006,19(3):434-437.
[4] 冉春,陈志永,陈洋,等.桔全爪螨田间种群对双甲脒的敏感性及其两种酶的生化特性[J].中国农学通报,2007,23(11):284-288.
[5] Ran Chun,Chen Yang,Wang Jinjun.Susceptibility and carboxylesterase activity of five field populations of Panonychus citri(McGregor)(Acari:Tetranychidae)to four acaricides [J].International Journal of Acarology,2009,35:115-121.
[6] 孟和生,王开运,姜兴印,等.桔全爪螨对常用杀螨剂的抗药性测定[J].农药,2000,39(2):26-28.
[7] 洪源范,洪青,沈雨佳,等.甲氰菊酯降解菌Sphingomonas sp. JQL4-5对污染土壤的生物修复[J].环境科学,2007,28(5):1121-1125.
[8] Lucija K,Nevenka K Z,Lenny H,et al.Toxicity of abamectin and doramectin to soil invertebrates[J].Environmental Pollution,2008,151:182-189.
[9] 刘永华,蒋红波,袁明龙,等.柑橘全爪螨对4种杀螨剂的抗性监测及增效作用[J].果树学报,2010,27(4):570-574.
[10]袁明龙,冉春,李勇,等.取食不同柑桔种质资源对桔全爪螨药剂敏感性及酯酶同工酶的影响[J].植物保护学报,2008,35 (2):187-188.
[11]FAO.Tentative method for spider mites and their eggs,Tetranychus spp.and Panonychus ulmi(Koch)[J].FAO Plant Protection Bulletin,1974,22:103-107.
[12]孟和生.两种生测方法对杀螨剂毒力测定结果的影响比较[J].农药,2002,28(3):49-51.
[13]孟和生,王开运,姜兴印,等.桔全爪螨的抗药性选育及其解毒酶活性变化[J].昆虫学报,2002,45(1):58-62.
[14]孟和生,王开运,姜兴印.桔全爪螨对甲氰菊酯的抗性选育及交互抗性的研究[J].植物保护,2000,26(6):7-9.
[15]Leeuwen T V,Vontas J,Tsagkarakou A,et al.Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari:A review[J].Insect Biochemistry and Molecular Biology,2010,40(8):563-572.
[16]何林,赵志模,邓新平,等.朱砂叶螨对三种杀螨剂的抗性选育与抗性风险评估[J].昆虫学报,2002,45(5):688-692.
[17]何林,赵志模,邓新平,等.朱砂叶螨对两种杀螨剂的抗性遗传力及风险评估[J].植物保护学报,2002,29(4):331-336.
[18]赵卫东,王开运,姜兴印,等.二斑叶螨对阿维菌素,哒螨灵和甲氰菊酯的抗性选育及其解毒酶活力变化[J].昆虫学报, 2003,46(6):788-792.
[19]Cochran D G.Monitoring for insecticide resistance in field-collected strains of the German cockroach(Dictyoptera:Blattellidae) [J].Journal of Economic Entomology,1989,82:336-341.
[20]孟和生,王开运,姜兴印,等.桔全爪螨对哒螨灵抗性的选育及其生化机理[J].农药学学报,2000,2(3):30-34.
[21]Yamamoto A,Yoneda H,Hatano R,et al.Laboratory selections of populations in the citrus red mite,Panonychus citri (McGregor),with hexythiazox and their cross resistance spectrum[J].Pesticide Science,1995,20:493-501.
[22]Yamamoto A,Yoneda H,Hatano R,et al.Field selection experiments with hexythiazox in the citrus red mite,Panonychuscitri(McGregor) [J].Journal of Pesticide Science(Japan),1995,20:307-315.
[23]Chen Z Y,Ran C,Zhang L,et al.Susceptibility and esterase activity in citrus red mite Panonychus citri(McGregor)(Acari:Tetranychidae)after selection with phoxim[J].International Journal of Acarology,2009,35:33-40.
(责任编辑:王 音)
Resistance selection and cross-resistance of Panonychus citri to fenpropathrin and abamectin
He Hengguo1, Wang Jinjun2
(1.College of Life Science,China West Normal University,Nanchong 637002,China; 2.College of Plant Protection,Southwest University,Chongqing 400715,China)
On the basis of resistance breeding and selection in laboratory,the resistance development of Panonychus citri to fenpropathrin and abamectin was investigated and the cross-resistance of Fe R and Ab R to 11 acaricides was also studied.The results showed that,after 11-and 16-times of selection with abamectin and fenpropathrin,the resistance of P.citri to the two acaricides increased by 3.80-and 29.92-fold,respectively.Bioassay revealed that the crossresistance of FeR to pyridaben,dicofol,azocyclotin and the cross-resistance of AbR to emamectin benzoate were very significant.The results provided references for the resistance management of P.citri to acaricides.
Panonychus citri; fenpropathrin; abamectin; cross-resistance
S 482.5
A
10.3969/j.issn.0529-1542.2015.06.036
2014-10-31
2014-11-10
公益性行业(农业)科研专项(201103020);四川省教育厅项目(11ZA293);西华师范大学博士启动基金(10B029)
联系方式 E-mail:hengguohe@163.com
猜你喜欢
化学工程师(2022年8期)2022-09-15
安徽农学通报(2022年6期)2022-04-07
农药科学与管理(2019年6期)2019-11-23
世界农药(2019年3期)2019-09-10
世界农药(2019年2期)2019-07-13
农家之友(2019年5期)2019-01-17
中国果业信息(2019年1期)2019-01-05
农业知识(2018年21期)2018-01-17
中国茶叶加工(2015年3期)2015-02-27
江苏农业科学(2014年10期)2014-11-22
