
遮阴对迷人杜鹃种子萌发和芽苗生长的影响
2015-07-01 08:01:56韦小丽李朝婵训1
贵州农业科学 2015年6期
周 艳,韦小丽,李朝婵,陈 训1,*
(1.贵州大学 林学院,贵州 贵阳550025;2.贵州省植物园,贵州 贵阳550004;3.贵州科学院,贵州 贵阳550001)
光是植物必需的资源之一,对植物的生长发育、生理生化和形态结构等方面有重要影响[1]。不同植物的种子在不同光强条件下的萌发行为和幼苗生长对光强的反应存在较大差异,且幼苗生长过程中表现出多种形态学可塑性,以适应环境光强的变化[2]。光照对植物种子萌发和幼苗的生长已受到学术界广泛关注。迷人杜鹃(RhododendronagastumBalf.)属常绿灌木或小乔木,枝条粗壮,花大,呈粉红色,产于云南、贵州等地,生于海拔1 600~1 900 m 的山坡常绿阔叶林或山地灌丛中,具有重要的观赏价值[3]。尽管光照对迷人杜鹃种子萌发影响方面的研究已有不少报道,但都是采用培养皿法研究迷人杜鹃种子的需光特性,且无一致结论。如黄承玲等[4]认为,迷人杜鹃种子是厌光型种子,在自然光暗交替的光照下发芽率更高,出苗更整齐,而任祝三[5]的研究表明,杜鹃属种子需光性种子,迷人杜鹃在实验室的黑暗条件下发芽率仅为4%。
为更好地模拟自然条件下光照对其种子和芽苗生长的影响,笔者用盆栽方法人工模拟光照条件,设置全光照、轻度遮阴、中度遮阴和重度遮阴4个水平的光照处理,培养2个月。统计计算种子的萌发率、萌发速率系数和萌发指数,测定不同光照条件下芽苗的胚轴和胚根长度,幼叶的长度及宽度,并测定芽苗根系活力和幼叶叶绿素含量,为阐明盆栽条件下迷人杜鹃种子萌发的光适应性,以及芽苗对光环境的适应能力,为迷人杜鹃的抚育管理提供科学依据。
1 材料与方法
1.1 试验区概况与材料
试验在贵州省植物园苗圃进行,地处贵阳市云岩区,东经106°42′,北纬36°24′,年均温15.2℃,年降水量12.00mm,年平均相对湿度80%。
试验用迷人杜鹃种子(R.agastumBalf.)于2013年12月采自贵州省百里杜鹃自然保护区境内约30年株龄的3 棵母树,采后自然风干,保存于5℃冰箱中。
1.2 试验设计
用不同透光率的黑色遮阴网进行遮阴处理,遮阴棚南北向,高1.2m,宽2.0m,东、西方向的遮阴网距地面0cm,南北敞开,便于通风透气。通过增加遮阴网的层数来调整遮阴程度,并用照度计精确测定。设置遮阴梯度分别为30%、50%、80%,以全光照作为对照(CK)。
于2014年4月25日以黄壤和腐殖土1∶2混合作基质,过筛后装入口径为15cm 的营养袋中。将采集的迷人杜鹃种子播种在营养袋中,每袋均匀撒播60粒种子于基质表面,保持土壤湿润,分别置于上述的4个光照条件下,每个处理10盆,3次重复。
1.3 种子萌发参数和芽苗生长指标的测定与计算
1.3.1 种子萌发参数 以幼苗第一片真叶出现为萌发标准,每天观察记录种子萌发情况,记录萌发的种子数,直到连续20d没有种子萌发为止。统计计算种子的萌发率(GP)、萌发速率系数(CRG)和萌发指数(GI)。

式中,t为萌发试验开始时的天数,n为t天内萌发的种子数;MDG 为日均种子萌发数,等于萌发试验结束时种子萌发数/萌发试验经历的天数;PV为种子最大萌发数,等于萌发试验期间任意一天中达到的最大萌发数/达到这一最大值所需的天数。
1.3.2 芽苗生长指标 发芽结束后,统计芽苗成活率,在每个处理的3次重复中分别随机抽取芽苗30株,测定其芽苗的胚轴和胚根长度,幼叶的长度及宽度。并计算芽苗活力指数(VI)。
VI=萌发率×(芽苗胚根长度+芽苗胚轴长度)。
在每个处理的3次重复中随机抽取芽苗根系测定其活力,幼苗叶片测定叶绿素。TTC 法测定根系活力,乙醇-丙酮混合提取分光光度法测定叶绿素。
1.4 数据处理
利用SPSS 18.0软件中的单因子方差分析和最小显著差法(LSD)进行各处理间数据的差异性分析。
2 结果与分析
2.1 遮阴处理迷人杜鹃的种子萌发
由表1 看出,迷人杜鹃种子萌发率差异显著(P<0.05),以30%遮阴处理种子萌发率最高(达90%),80%遮阴处理种子萌发率最低(40.00%);与全光照相比,50%和80%遮阴处理的种子萌发率分别降低20.45%和43.30%(P<0.05),30%遮阴处理的种子萌发率提高21.61%。随着光照的减弱,萌发速率系数先增加后减少,弱光照对迷人杜鹃种子的萌发进程有一定的延迟作用,和全光照相比,50%和80%遮阴处理的萌发速率系数分别降低20.15%和54.57%(P<0.05),30%比对照提高10.61%(P>0.05)。不同处理间萌发指数差异显著(P<0.05),其中以30%遮阴处理萌发指数最高(2.46),80%遮阴处理萌发指数最低(0.27)与全光照相比,50%和80%遮阴处理的萌发指数分别降低40.23%和68.97%(P<0.05),30%遮阴处理的萌发指数提高64.92%。
2.2 遮阴处理迷人杜鹃的芽苗生长
由表2所示,不同处理间的芽苗胚根长、胚轴长和芽苗活力指数差异显著(P<0.05),随着光照强度的减弱,均呈先升高后降低的趋势。与全光照相比,30%遮阴处理芽苗胚根长度提高60.71%、胚轴长度提高29.91%、芽苗活力指数增加52.26%(P<0.05);50%和80%遮阴处理的胚根长分别降低27.27%和59.09%,胚轴长分别降低26.83%和54.88%,芽苗活力指数减少53.24%和74.32%(P<0.05)。从幼叶的生长看,各处理间的幼叶长度差异不显著,以30%遮阴处理的叶片长度最长;与全光照相比,各处理的叶片宽度差异性显著(P<0.05),30%遮阴处理叶片宽度提高33.33%,50%和80%遮阴处理的叶片宽度分别降低50.00%和66.67%。以上试验结果表明,30%遮阴处理有利于迷人杜鹃芽苗生长,随着光照强度的减弱芽苗的生长受到抑制。

表1 不同遮阴处理迷人杜鹃的种子萌发Table 1 Seed germination of R.agastumof different shading treatment
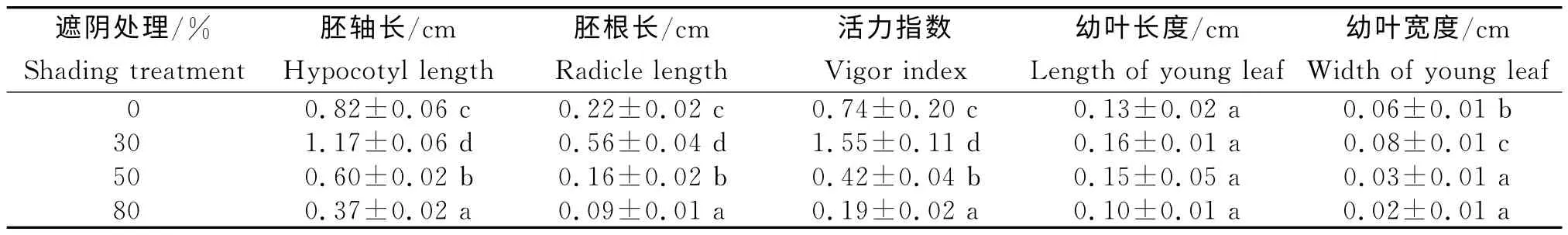
表2 遮阴处理迷人杜鹃的芽苗生长Table 2 Seedling growth of R.agastumof different shading treatment
2.3 光照对迷人杜鹃芽苗生理生化指标的影响
2.3.1 芽苗根系活力 由图示可见,与全光照相比,30%遮阴处理的幼苗根系活力提高38.11%(P<0.05)[3.49mg/(g·h)],促进幼苗根的生长。随着遮阴强度的提高,迷人杜鹃幼苗根系活力降低,50%、80%遮阴处理的根系活力分别降低43.06%、78.24%,说明,在弱光照下芽苗根的生长受到抑制。
2.3.2 丙二醛(MDA)含量 由图示还可见,随着遮阴度的增加,MDA 含量逐渐上升。30%遮阴处理和全光照无显著差异(P>0.05),与50%和80%遮阴处理差异性显著(P<0.05)。其中,以80%遮阴处理芽苗MDA 含量最高,达13.42nmol/g FW,分别比全光照、30%和50%遮阴处理提高61.85%、60.21%和22.35%。
2.3.3 叶绿素含量 从表3 可见,叶绿素a含量以50%遮阴处理的含量最高,为49.07μg/g,与其他处理差异显著(P<0.05),分别比全光照、30%和80%遮阴处理提高16.09%、16.61%和10.68%。
不同处理间叶绿素b 含量差异性显著(P<0.05),以50%遮阴处理的叶绿素b含量最高,为26.24μg/g,分别比全光照、30%和80%遮阴处理提高40.62%、21.38%和2.02%。不同处理间总叶绿素含量差异显著(P<0.05),随着遮阴度的增加,叶绿素含量呈先上升后下降的趋势。以50%遮阴处理叶绿素含量最高,为75.32μg/g,分别比全光照、30%和80%遮阴处理提高24.65%、18.27%和7.67%;以全光照处理的的叶绿素含量最低,为56.75μg/g,分别比30%、50%和80%遮阴处理降低10.19%、18.42%和4.65%。而叶绿素a/b以全光照处理最高,为2.64,与其他处理差异性显著(P<0.05),分别比30%、50%和80%遮阴处理高25.00%、29.17%和35.23%。
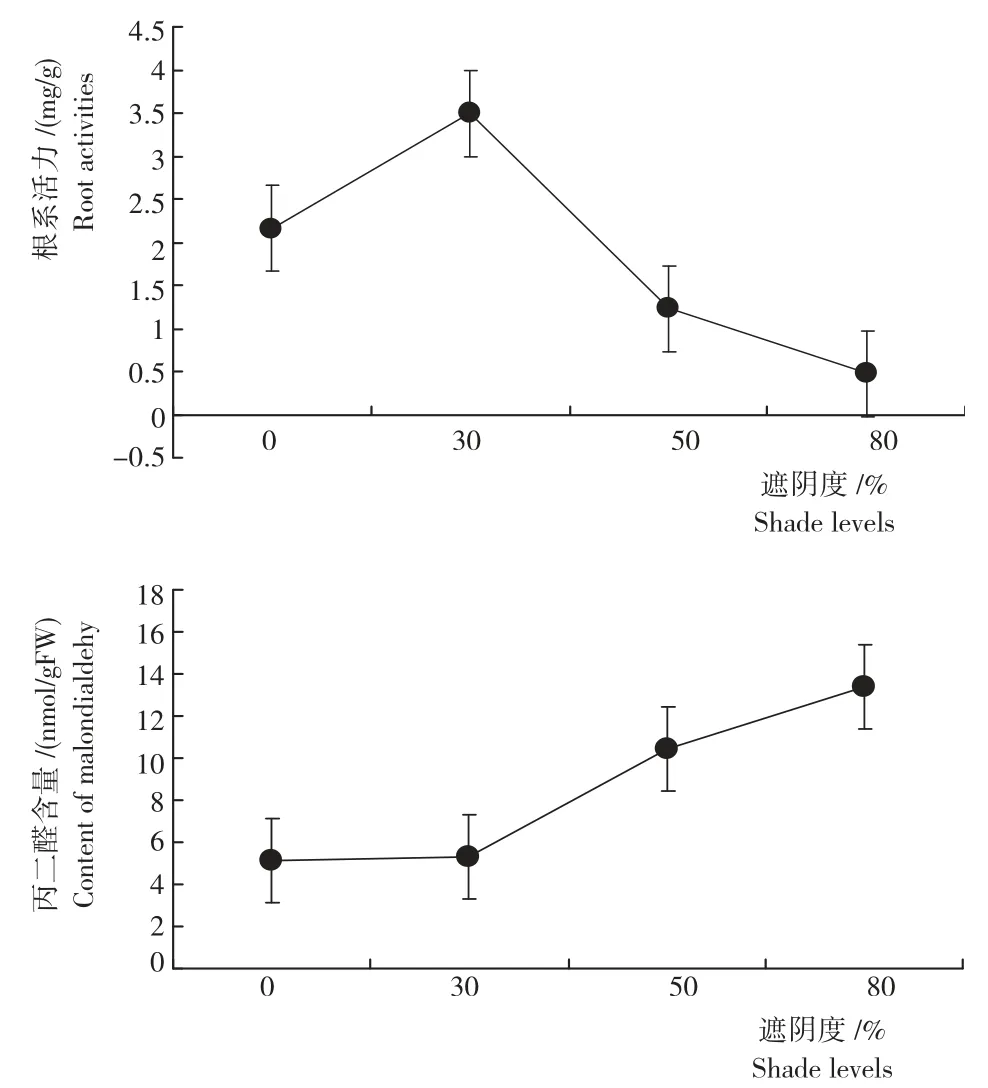
图示 不同遮阴处理迷人杜鹃芽苗的根系活力和丙二醛含量Fig.Root activities and MDA content of R.agastum of different shading treatment

表3 遮阴处理迷人杜鹃芽苗的叶绿素含量Table 3 Chlorophyll content of R.agastumseedlings of different shading treatment
3 结论与讨论
试验表明,迷人杜鹃种子萌发率和萌发指数在30%遮阴处理时达到最大,分别为90.00%和2.46,而在80%遮阴处理下最小,为40%和0.27。弱光照对种子萌发进程有一定的延迟作用,萌发活力指数随着光照的减弱而减小。不同光强处理的迷人杜鹃芽苗具有较大的形态学可塑性,30%遮阴处理的芽苗胚根长、胚轴长、芽苗活力指数和幼叶宽均最大,分别比全光照提高60.71%、29.91%、52.26%和33.33%。根系活力以30%遮阴处理值最大,为3.49mg/(g·h);MDA 含量以80%遮阴处理值最大,为13.42nmol/g FW;叶绿素含量以50%遮阴处理值最大,为75.32μg/g。
光照是影响植物种子萌发的重要环境因子。Drime等[6]在光照、部分遮阴和全黑暗3 种条件下研究了271种植物的种子萌发,结果发现,大多数植物种子萌发率在光照和部分遮阴条件下很高,在黑暗中很低,其中104种植物的种子在黑暗中萌发率不超过10%。本研究中,30%遮阴处理使迷人杜鹃种子的萌发率和萌发指数增加,而在80%遮阴处理下最小,说明盆栽条件下迷人杜鹃的种子萌发需要一定的光照条件。任祝三[5]的研究表明,迷人杜鹃在实验室的黑暗条件下发芽率仅为4%,种子萌发需要一定的光照条件,和本文的研究一致。而由于试验方法、时间、地点的不一致,黄承玲等[4]认为迷人杜鹃种子是厌光型种子。
强光下植物根系生长的投资更大,因而发育出更发达的根系[7]。弱光下耐阴植物幼苗比非耐阴植物幼苗的比叶面积更小、根冠比更高[8]。本研究认为,30%遮阴处理对迷人杜鹃芽苗生长更有利,其芽苗胚根长、胚轴长、芽苗活力指数、幼叶宽均最大。
在一定光强范围内,幼苗叶片的叶绿素含量随着光强的增加而降低[9]。本研究中叶绿素b和叶绿素总量随遮阴度的增加(即光强降低)呈现出先增加后降低的趋势。另据柯世省等[10]报道,过于荫蔽的光照环境不利于叶绿体的充分发育。与强光照处理相比,迷人杜鹃幼叶叶绿素含量在光照最弱的80%遮阴处理下有所降低。根系活力是根系代谢强度的综合反应[11],植物对荫蔽环境的适应常以根系生物量积累的减少为代价[12],因此弱光条件下导致迷人杜鹃芽苗根系活力降低。MDA 含量的高低可衡量质膜受伤害或变性程度[13],试验中,迷人杜鹃幼苗MDA 含量随遮阴度的增加而逐渐上升,表明深度遮阴对幼苗的生长不利。
[1]马银山,杜国祯,张世挺.光照强度和肥力变化对垂穗披碱草生长的影响[J].生态 学报,2014,34(14):3908-3916.
[2]闫兴富,曹 敏.不同光照对望天树种子萌发和幼苗早期生长的影响[J].应用生态学报,2007,18(1):23-29.
[3]陈 训,巫华美.中国贵州杜鹃花[M].贵阳:贵州科技出版社,2003.
[4]黄承玲,周洪英,黄家涌.迷人杜鹃种子特性与萌发实验研究[J].种子,2009,28(8):96-98.
[5]任祝三.光与激素对于杜鹃花种子发芽的作用[J].云南植物研究,1986,8(1):73-80.
[6]Grime J P,Mason,G,Curtis A V,et al.A comparative study of germination characteristics of a local flora[J].Journal of Ecology,1981,69:1017-1059.
[7]Karel M,Raison R J,Anatolys P.Critical analysis ofroot:Shoot ratios in terrestrial biomes[J].Global Change Biology,2006,12:84-96.
[8]Portsmuth A,Niinemets U.Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shshade tolerance.Functional Ecology,2007,21:61-77.
[9]郭晋平,李海波,刘 宁,等.华北落叶松和白NFDA4幼苗对光照和竞争响应的差异比较[J].林业科学,2009,45(2):53-59.
[10]柯世省,金则新,林恒琴,等.天台山东南石栎光合生理生态特性[J].生态学杂志,2004,23(3):1-5.
[11]孙文越,王 辉,黄久常.外源甜菜碱对干旱胁迫下小麦幼苗膜脂过氧化作用的影响[J].西北植物学报,2001,21(3):487-491.
[12]闫兴富,王建礼,周立彪.光照对辽东栎种子萌发和幼苗生长的影响[J].应用生态学报,2011,22(7):1682-1688
[13]孙文越,王 辉,黄久常.外源甜菜碱对干旱胁迫下小麦幼苗膜脂过氧化作用的影响[J].西北植物学报,2001,21(3):487-491.
猜你喜欢
新作文·小学低年级版(2021年9期)2022-01-01 07:50:34
心声歌刊(2021年3期)2021-08-05 07:43:52
小聪仔(幼儿版)(2020年12期)2020-02-01 07:08:46
民族音乐(2018年5期)2018-11-17 08:20:00
乡村地理(2018年3期)2018-11-06 06:50:30
现代园艺(2017年21期)2018-01-03 06:41:56
青年文学家(2017年28期)2017-11-28 15:41:54
读写算(中)(2015年25期)2015-11-07 07:25:58
福建林业科技(2015年3期)2015-09-16 02:51:57
作文周刊·小学一年级版(2014年26期)2014-04-29 00:44:03
