
四川牡丹的花部特征和繁育系统研究
2015-06-24 14:32:14骆劲涛张必芳宋会兴刘光立曾秀丽
植物资源与环境学报 2015年4期
杨 勇, 骆劲涛, 张必芳, 宋会兴, 刘光立,①, 曾秀丽
(1. 四川农业大学: a. 风景园林学院, b. 玉米研究所 农业部西南玉米生物学与遗传育种重点实验室, 四川 成都 611130;2. 西藏自治区农牧科学院蔬菜研究所, 西藏 拉萨 850032)
四川牡丹的花部特征和繁育系统研究
杨 勇1a,1b, 骆劲涛1a, 张必芳1a, 宋会兴1a, 刘光立1a,①, 曾秀丽2,①
(1. 四川农业大学: a. 风景园林学院, b. 玉米研究所 农业部西南玉米生物学与遗传育种重点实验室, 四川 成都 611130;2. 西藏自治区农牧科学院蔬菜研究所, 西藏 拉萨 850032)
对分布于四川阿坝的四川牡丹(PaeoniadecompositaHand.-Mazz.)的花部特征和开花动态进行了观察,并通过分析花粉活力、估算杂交指数(OCI)、控制授粉方式和观察访花昆虫对其繁育系统进行了研究。四川牡丹花部主要特征为:花色粉红色至红紫色,随开放时间延长渐变为粉白色;辐射状花冠,花瓣8~12枚;多体雄蕊128~203枚,花丝白色,花药黄色,有雄蕊瓣化现象;柱头位于心皮顶部,黄色;边缘胎座,心皮2~7枚,单个心皮含胚珠8~33枚;开花过程中均散发花香,但随花期延长花香略有变化。该种的群体花期相对集中,为4月中旬至5月中旬,但不同年份间略有差异;单花花期多数为6~8 d,整个开花过程中花药与柱头基本处于同一平面,雄蕊先熟。从开花第1天至第6天,该种的花粉活力先升高后降低,其中开花第3天至第5天花粉活力较高(约80%);从开花第2天起柱头普遍具有可授性,长达9 d,开花第5天至第8天柱头可授性最强;花粉具活力和柱头具可授性的重叠时间为5 d;该种的OCI值为4,花粉/胚珠比(P/O)为207 220。四川牡丹花期不分泌花蜜,花粉是传粉昆虫的惟一报酬;主要访花昆虫为蜜蜂(Apidea)、食蚜蝇(Syrphidae)和花金龟(Cetoniinae),其中前2类昆虫可能为该种的有效传粉者,而花金龟的传粉效率较低。授粉实验结果显示:2012年和2014年自然授粉植株的平均结籽率最高,为23.76%;而异株异花人工授粉和同株异花人工授粉植株的平均结籽率均较低,分别为18.33%和11.04%。研究结果表明:四川牡丹的繁育系统属于异交,部分自交亲和,不存在无融合生殖,需要传粉者授粉。
四川牡丹; 花部特征; 开花进程; 访花昆虫; 授粉方式; 繁育系统
被子植物的繁育系统是指包括花部特征、花器官寿命、花朵开放式样和授粉亲和性等直接影响后代遗传组成的所有特征;它是物种性别表达对下一代个体相对遗传贡献的首要影响因子,能有效调控种群遗传结构[1]。植物的花部特征与繁育系统对植物的生殖成功与否至关重要[2],影响植物的遗传多样性及分布格局[3],因此,深入了解植物的花部特征与繁育系统对濒危植物的保护及开发利用具有指导作用[4-5]。
四川牡丹(PaeoniadecompositaHand.-Mazz.)隶属于芍药科(Paeoniaceae)芍药属(PaeoniaLinn.)牡丹组(Sect.MoutanDC.),为珍稀濒危植物[6-7]。该种仅分布于四川西北部的大渡河及岷江上游的河谷地带[8],分布地域较偏僻且狭窄,对其相关研究较少,且主要集中在分类学[8-9]、生态学[10]、种子休眠机制[11-15]和分子生物学[16-18]等方面。2011年,国家卫生部批准牡丹籽油为新型木本食用油,因此,培育优质高产油用牡丹新品种成为新的研究热点之一。四川牡丹专性种子繁殖[19],结实量大,是培育油用牡丹新品种的潜在资源。为深入了解四川牡丹的繁育系统特征,作者对四川牡丹的花部特征、开花进程以及传粉生物学特征进行了观察和研究,以期为该种类的有效保护、人工扩繁和杂交育种提供基础研究数据。
1 研究地概况和研究方法
1.1 研究地概况
研究地点位于四川省阿坝藏族羌族自治州马尔康县脚木足乡,位于东经102°02′20″、北纬31°59′20″。在四川牡丹集中分布的区域内设置1块面积30 m×30 m的永久样地。样地海拔2 500~2 550 m;年平均气温8.3 ℃、 极端最高气温31.0 ℃、 极端最低气温-16.6 ℃;年降水量786 mm,90%降水集中在5月至9月;土壤为山地黄壤(约pH 7.5),母岩为花岗岩,土壤有机质含量5.65 g·kg-1。
该群落是以四川牡丹、野花椒(ZanthoxylumsimulansHance)、小叶蔷薇(RosawillmottiaeHemsl.)和豪猪刺(BerberisjulianaeC. K. Schneid.)为优势种的林缘灌丛,主要伴生植物有金花小檗(BerberiswilsonaeHemsl.)、四川丁香(SyringasweginzowiiKoehne et Lingelsh.)、鸢尾(IristectorumMaxim.)、野棉花(AnemonevitifoliaBuch.-Ham. ex DC.)、川滇铁线莲(Clematisclarkeanavar.stenophyllaHand.-Mazz.)和瞿麦(DianthussuperbusLinn.)等。该样地中四川牡丹居群包含96株成年开花植株,是目前发现的成年个体最多的四川牡丹居群。
1.2 研究方法
1.2.1 花部特征和开花动态观察 分别于2012年和2014年的4月份至6月份,对样地内四川牡丹群体花期、单花花期、花部特征及变化进行观测。统计样地内正常发育的花蕾数,并以开花花朵数约占全部花蕾数的5%为初花期,开花花朵数大于全部花蕾数的50%为盛花期,凋谢花朵数大于全部花蕾数的95%为末花期。选取花蕾数量相近的个体15株,每株标记正常发育的花蕾1朵,观察、测量和统计花瓣长、花瓣宽、花瓣数量、苞片形态、花托形态、雄蕊数量和雌蕊数量等花部特征并记录其变化,并统计单花花期;用直尺(精度1 mm)分别测量花瓣最长处和最宽处的长度,即为花瓣长和花瓣宽;单花花期为从花瓣完全展开露出雄蕊至第1片花瓣凋落期间的天数。
1.2.2 花粉活力和柱头可授性检测 采用TTC法[20]检测花粉活力。于2012年4月下旬选取来自不同植株且发育正常的花蕾6朵,在开花后于每天11:00分别取花粉进行花粉活力检测,直花瓣脱落止。
采用联苯胺-过氧化氢法[21]检测柱头可授性。于2012年4月下旬,在盛花期选择株龄和花蕾数目相近的个体,每天上午选取并标记来自不同个体且当日开放的花5朵,连续标记,直至第1天标记的5朵花的柱头完全变黑;收集所有标记的花,取各花的柱头,置于盛有联苯胺-H2O2溶液〔V(质量分数1%联苯胺)∶V(体积分数3%H2O2)∶V(蒸馏水)=4∶11∶22〕的凹形载玻片内,观察其变化;若柱头具有可授性,则柱头周围溶液变蓝且产生大量气泡。
1.2.3 杂交指数估算 参照文献[20]的方法估算杂交指数(outcrossing index,OCI)。于2012年4月下旬随机选取四川牡丹不同个体上发育正常、开放程度一致的花20朵,按照以下赋值标准并根据花朵直径及开花行为估算OCI值。1)花朵直径(d):d≤1 mm记为0;1
依据OCI值对繁育系统进行判断,评判标准为:OCI值为0,繁育系统为闭花受精(cleistogamy);OCI值为1,繁育系统为专性自交(obligate autogamy);OCI值为2,繁育系统为兼性自交(facultative autogamy),有一定的异交可能;OCI值为3,繁育系统为兼性异交(facultative xenogamy),自交亲和,有时需要传粉者;OCI值为4,繁育系统为异交(xenogamy),部分自交亲和,需要传粉者。
1.2.4 花粉/胚珠比分析 于2012年4月下旬,在四川牡丹不同个体上随机选取发育正常且即将开放的花蕾20朵,用FAA固定液〔V(体积分数70%乙醇)∶V(冰乙酸)∶V(体积分数38%甲醛)=18∶1∶1〕固定。参考张文标等[22]的方法进行处理,用OLYMPUS CX31体视显微镜(日本奥林巴斯株式会社)观察并统计每朵花的胚珠数,计算花粉/胚珠比(pollen-ovule ratio,P/O),并按Cruden[23]的标准初步判定四川牡丹的繁育系统类型。
1.2.5 授粉实验 分别于2012年和2014年的4月进行人工授粉实验。具体授粉方法为:1)直接套袋(不去雄套袋),选取即将开放的发育正常的花朵,套硫酸纸袋并挂牌标记。2)去雄套袋,选取即将开放的发育正常的花朵,去掉所有雄蕊,套硫酸纸袋并挂牌标记。3)同株异花授粉(人工自交),选取即将开放的发育正常的花朵,去掉所有雄蕊,套硫酸纸袋并挂牌标记;通过观察未套袋花朵确定花期,从开花第3天开始,连续3 d进行人工授粉,所用新鲜花粉采自同株其他的散粉花朵,授粉完成后重新套袋。4)异株异花授粉(人工异交),选取即将开放的发育正常的花朵,去掉所有雄蕊,套硫酸纸袋并挂牌标记;通过观察未套袋花朵确定花期,从开花第3天开始,连续3 d进行人工授粉,所有新鲜花粉采自距其5 m外的其他植株,授粉完成后重新套袋。5)自然授粉(对照),选择即将开放的正常发育花朵,挂牌标记但不套袋,进行自然授粉。
观察自然授粉花朵的变化,待其花瓣和雄蕊全部脱落、柱头黏液变干、柱头变黑时去掉其他4个处理的纸袋,让果荚在相同条件下生长发育。2012年每组处理50朵花;2014年因开花数量较少,每组处理20朵花。分别于2012年和2014年的8月中旬果实成熟但果荚未开裂时收集果实,每个蓇葖果分别采收分装,并统计结籽率。
1.2.6 访花昆虫观察 于2014年4月下旬,在四川牡丹盛花期时选择5个晴朗日,选择6朵花,从9:00至16:00对访花昆虫进行观察,统计访花昆虫的种类和访问频次。
1.3 数据统计与分析
采用EXCEL 2010数据处理软件对实验数据进行统计和计算,采用SPSS 19.0统计分析软件进行差异显著性分析。
2 结果和分析
2.1 四川牡丹花部特征观察和开花动态分析
四川牡丹的花部特征见图1。在四川牡丹每支成熟枝条的枝顶正常着生1朵花,花色粉红色至红紫色(图1-1),随开放时间延长逐渐变为粉白色;辐射状花冠,花被覆瓦状折叠;花瓣数8~12枚,以10枚居多,花瓣长41.0~87.5 mm,平均60.1 mm;苞片3~5枚,披针形、叶形或渐变成枝条,多为绿色,少数出现与花瓣颜色接近的杂色;花托碗型,离生花萼卵圆形或匙形;多体雄蕊128~203枚不等,平均164枚,花丝白色,花药黄色,有雄蕊瓣化现象;边缘胎座,心皮2~7枚,多数5枚,光滑无毛;柱头位于心皮顶部,扇形,黄色,脊部分泌透明黏液;单个心皮含胚珠8~33枚,平均17枚。
四川牡丹群体花期为每年4月中旬至5月中旬,不同年份间略有差异,2012年为4月13日至5月15日,2014年为4月14日至5月12日。始花期和盛花期开放的花朵较大;末花期多出现瘦小花蕾,花朵较小,且存在腋生花。统计结果显示:四川牡丹单花花期为6~13 d,多数为6~8 d。
开花过程中,花瓣长度随花朵开放时间延长逐渐增加,平均值从44.8 mm增至63.9 mm;雄蕊先于雌蕊成熟,且雄蕊内轮花药先开裂(图1-2);开始散粉的雄蕊花药与柱头几乎平齐(图1-3),开放过程中柱头高度变化不明显,而花药则逐渐干瘪扭曲、花丝伸长,一般在散粉的第3天花药高过雌蕊。在开花之后花朵气味随花期延长逐渐变化,初期为稍刺鼻的类似中药丹皮的异香,后期变为类似枇杷果实的果香。从花朵张开至花瓣脱落都未见花蜜分泌。

1. 全花Whole flower; 2. 示花药开裂状态,椭圆形内部的花药开裂,外部花药未开裂Showing anther dehiscencing status, anthers dehiscencing at inside of ellispse, and not dehiscencing at outside; 3. 示雄蕊与柱头基本等高,箭头指示柱头Showing stamens and stigma almost with the same height, arrows indicating stigma.
图1 四川牡丹的花部特征
Fig. 1 Floral characteristics ofPaeoniadecompositaHand.-Mazz.
2.2 四川牡丹花粉活力和柱头可授性分析
四川牡丹花粉活力和柱头可授性随开花时间的变化见表1。由表1可以看出:在开花的第1天至第6天,四川牡丹的花粉活力呈先升高后降低的变化趋势,其中,开花后第3天至第5天花粉活力较高,可达约80%;开花第7天,花药脱离母体。在开花第2天(即花药开裂后第2天),其柱头普遍具有可授性;开花第5天,花粉活力开始降低,柱头可授性达到最强。四川牡丹柱头可授性持续时间长达9 d,其中开花第5天至第8天柱头可授性最强。花粉具活力和柱头具可授性的重叠时间为5 d, 其中花粉活力较强与柱头可授性较强的重叠时间为4 d。

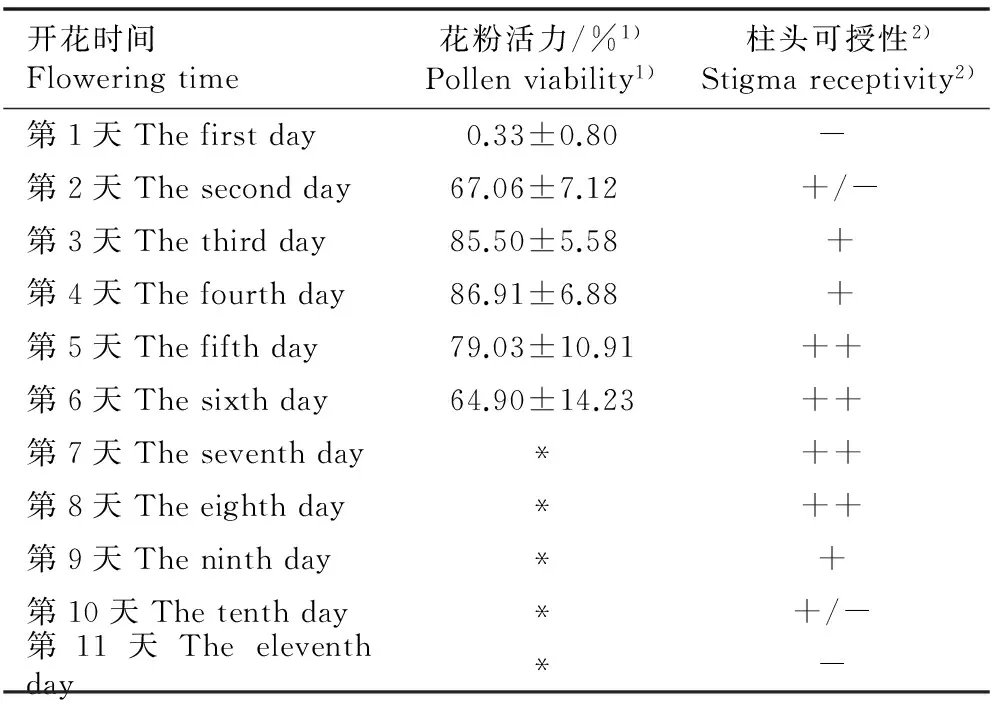
开花时间Floweringtime花粉活力/%1)Pollenviability1)柱头可授性2)Stigmareceptivity2)第1天Thefirstday0.33±0.80- 第2天Thesecondday67.06±7.12+/-第3天Thethirdday85.50±5.58+第4天Thefourthday86.91±6.88+第5天Thefifthday79.03±10.91++ 第6天Thesixthday64.90±14.23++ 第7天Theseventhday*++ 第8天Theeighthday*++ 第9天Theninthday*+ 第10天Thetenthday*+/- 第11天Theeleventhday*-
1)*: 花药脱落Anther falling off.
2)-: 柱头不具可授性Stigma without receptivity; +/-: 部分柱头具有可授性,部分柱头不具有可授性Some stigmas with receptivity, and some without; +: 柱头具可授性Stigma with receptivity; ++: 柱头可授性较强Stigma with strong receptivity.
2.3 四川牡丹杂交指数(OCI)与花粉胚珠比(P/O)分析
参照文献[20]的方法计算四川牡丹的杂交指数(OCI)。四川牡丹的花冠最小直径67.0 mm,平均直径119.4 mm,赋值为3;花药于开花当天开裂,且花粉具有活性,而从开花第2天开始部分柱头具可授性,故为雄蕊先熟,赋值为1;花药和柱头几乎在同一高度上,赋值为0。据此计算出四川牡丹的OCI值为4,显示四川牡丹繁育系统属于异交,部分自交亲和,需要传粉者。
四川牡丹平均单花花粉量12 225 952粒,平均单花胚珠数59个,P/O值为207 220,按照Cruden[23]的标准,四川牡丹的有性繁殖类型为专性异交。
2.4 采用不同授粉方式后四川牡丹结实状况分析
采用不同的授粉方式后四川牡丹的结实状况见表2。由表2可以看出:采用不去雄套袋方式进行授粉,2012年和2014年四川牡丹的结籽率均较接近,平均值为4.06%,说明该种有一定程度的自交亲和性。采用去雄套袋方式进行授粉,2012年和2014年共收获63个蓇葖果,但果实中未见发育完全的种子,说明四川牡丹不存在无融合生殖。采用同株异花人工授粉方式,2012年和2014年的结籽率分别为7.13%和14.94%,平均结籽率为11.04%,表明四川牡丹部分自交亲和;2年的结籽率差异较大,推测可能与不同年份的天气状况差异相关。采用异株异花人工授粉方式,2012年和2014年的结籽率分别为21.49%和15.17%,平均结籽率为18.33%。采用自然授粉方式,2012年和2014年的结籽率分别为26.43%和21.08%;平均结籽率最高,为23.76%。根据以上结果推断:四川牡丹繁育系统为部分自交亲和,主要为昆虫传粉的异交繁殖类型。

方式1)Measure1)年份Year花朵数Numberofflowers心皮数Numberofcarpels种子数Numberofseeds结籽花朵数Numberofflowerswithseedsetting结籽心皮数Numberofcarpelswithseedsetting胚珠数Numberofovules结籽率/%Seedsettingrate平均结籽率/%2)Averageofseedsettingrate2)1201245225110345932953.344.06±1.02cd2014168060111812564.78220124623000035230.000.00±0.00d2014178500013900.0032012472352484218634787.1311.04±5.52bc201417851741455116514.94420124522971442200332321.4918.33±4.47ab201414701571340103515.17520124823994045215355626.4323.76±3.78a201414702141245101521.08
1)1: 不去雄套袋Baging without emasculation; 2: 去雄套袋Baging after emasculation; 3: 同株异花人工授粉Artificial geitonogamy; 4: 异株异花人工授粉Artificial xenogamy; 5: 自然授粉Natural pollination.
2)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2.5 四川牡丹花期访花昆虫及传粉行为观察
四川牡丹花期的主要访花昆虫见图2。2014年4月下旬样地早晚气温较低,在早晨和傍晚很少能看到昆虫在花朵之间活动。9:00至10:00活动昆虫也较少,仅发现鞘翅目(Coleoptera)昆虫停留于雄蕊群中,且以花金龟(Cetoniinae)最多,该类昆虫在花内几乎不爬动,但停留时间较长(超过5 min),并可观察到这类昆虫在花内有交配行为(图2-1),同时该类昆虫也会取食花粉。10:00之后气温逐渐升高,昆虫开始大量出现,其中以膜翅目(Hymenoptera)的蜜蜂(Apidea)(图2-2)和双翅目(Diptera)的食蚜蝇(Syrphidae)(图2-3)居多。蜜蜂常在花朵间活动,多数情况下停落在花色鲜艳的花朵上,并在雄蕊和雌蕊之间爬动,用前足不停扫动并带走大量花粉,有时可看到蜜蜂的携粉足上黏附着大块的花粉粒;食蚜蝇的活动方式与蜜蜂相似,但在每朵花上停留的时间极短(多数为3~5 s),之后立即飞至其他花上。15:00之后气温逐步降低,活动的昆虫数量逐渐减少,在16:00之后很难看到昆虫在观察区域活动。
在5 d的观察过程中,共记录146次昆虫活动。蜜蜂共记录61次,占昆虫活动总次数的41.8%,其中,当地农民饲养的阿坝蜜蜂〔ApisceranaAbacerana(Aba race) Fabricius〕(图2-2)在四川牡丹的花上活动频繁,获取花粉;也可观察到隧蜂(Halictidaesp.)和切叶蜂(Megachilidaesp.)在花间活动。食蚜蝇(图2-3)共记录46次,占昆虫活动总次数的31.5%。鞘翅目昆虫共记录29次,占昆虫活动总次数的19.9%,主要为花金龟(图2-1),也有少量跳甲(Alticinaesp.)和拟步甲(Tenebrionidaesp.)。观察结果还表明:蜜蜂和食蚜蝇在花朵间活动频繁,且在雄蕊群和雌蕊群之间来回爬动,推测这2类昆虫可能为四川牡丹的有效传粉者;花金龟多数情况下仅停留在雄蕊群中吞食花粉,极少爬动,推测其传粉效率较低。四川牡丹不分泌花蜜,昆虫在花间活动主要是为了取食花粉。

1. 小青花金龟Oxycetonia jucunda Faldermann; 2. 阿坝蜜蜂Apis cerana Aba cerana (Aba race) Fabricius; 3. 食蚜蝇Syrphidae.
3 讨 论
在被子植物中极少有绝对的自交或异交类群,大多数植物都是同时利用2种交配模式来完成授粉[24]。本研究的观察结果显示:四川牡丹的有性繁育系统为异交,部分自交亲和,需要传粉者,且四川牡丹有性繁殖为专性异交。在四川牡丹的单花开花初期,雄蕊群和雌蕊群几乎处于同一高度,之后花丝伸长,高过柱头,花粉活力与柱头可授期在时间上有6 d重叠,因此在单花水平上四川牡丹的花部特征使自花授粉成为可能。授粉实验结果显示:四川牡丹不存在无融合生殖,通过自花授粉可以结籽,但结籽率较低;同株异花人工授粉的结籽率低于异株异花人工授粉,而异株异花人工授粉的结籽率又低于自然授粉,但差异不显著,说明在自然条件下,因开花植株花期集中,在四川牡丹居群内可通过昆虫实现异交,因而,四川牡丹花期是以异花授粉为主,部分自交亲和,需要传粉者。四川牡丹的自然授粉和人工授粉的结籽率均低于25%,且芍药属其他种类的自然授粉结籽率也普遍偏低[25-30],这可能和芍药属植物的遗传特性有关,而较低的自然结籽率也可能是导致芍药属植物濒危的重要原因之一。
四川牡丹花朵较大、花朵坚实、花瓣较为平展,这些特点都有利于甲虫类和蜂类昆虫在花上停留或传粉;四川牡丹在开花过程中都会散发出浓郁的花香,虽然花香能够吸引甲虫类和蜂类昆虫为植物传粉[31],但蜂类昆虫对红色不敏感,对白色和黄色较为敏感[32],而四川牡丹花瓣为红色、雄蕊和柱头均为黄色、花丝为白色,四川牡丹的花色更有利于吸引蜂类昆虫停留在雄蕊或柱头上进行传粉;气味和花色仅作为信号引诱传粉昆虫,而最终吸引昆虫且能补偿访花所消耗能量的是花粉或花蜜[33],四川牡丹没有蜜腺,大量的花粉是其提供给传粉昆虫的惟一报酬,与金红等[34]对白花野牡丹(Melastomacandidumf.albiflorumJ. C. Ou)访花昆虫的观察结果一致。以上这些特征都有利于甲虫类和蜂类昆虫协助四川牡丹完成授粉。通过连续多天的观察发现,在四川牡丹花上活动的昆虫主要是蜜蜂、食蚜蝇和花金龟;其中,花金龟取食花粉,通过在花上的活动起到传粉作用,但其体壁光滑、不易携带花粉且活动迟缓,传粉效率较低;蜜蜂在访花过程中虽然在每朵花上停留的时间较短,但其频繁更换花朵,有利于完成授粉;食蚜蝇的访花行为和蜜蜂类似,且访花频率仅次于蜜蜂,因此推测蜜蜂和食蚜蝇是四川牡丹花期的主要有效传粉昆虫。罗毅波等[28]和李奎等[29]认为蜂类昆虫是矮牡丹(PaeoniajishanensisT. Hong et W. Z. Zhao)和滇牡丹(P.delavayiFranch.)花期的主要传粉昆虫,四川牡丹的传粉昆虫与同属的这2种植物相似,但存在一定差异。
四川牡丹花期的访花昆虫中最重要的一类是阿坝蜜蜂,该类蜜蜂是东方蜜蜂(ApisceranaFabricius)阿坝地理种群Abacerana(Aba race)[35]。该地理种群是阿坝地区特有的野生蜜蜂种群,适应高海拔、低气温的环境条件,自然生活于河谷地带[36],当地农民多在山谷隐蔽处建立简易蜂箱吸引阿坝蜜蜂筑巢以获取蜂蜜和花粉。因为人类干预使得阿坝蜜蜂在该地区繁殖较快,数量较多。在每年的4月份和5月份,四川牡丹原生地气温较低,多数植物才开始萌发,除四川牡丹外极少有植物开花,此时四川牡丹的花粉成为阿坝蜜蜂的重要食物,而阿坝蜜蜂在花朵上频繁采集花粉的活动有利于四川牡丹完成授粉。
面临强大的选择压力,多数濒危植物选择了大量、集中开花的方式以吸引更多的传粉昆虫[37-38],四川牡丹也具有这一特点。四川牡丹群体花期较短(不足1个月),盛花期在2周内,花期高度一致更加有利于吸引传粉昆虫完成授粉。四川牡丹的花粉可在较长时间内保持活力,其中有5 d花粉活力保持在60%以上;而律春燕[39]的研究结果也表明,在室温条件下,黄牡丹(PaeonialuteaDelavay ex Franch.)花粉生活力能保持10 d左右,低温条件下储存1 a后花粉萌发率仍大于48%,因此,可以推测花粉活力保持时间较长是牡丹组植物的共同特性。高海拔地区传粉昆虫较平原地区少[40],延长花期有利于完成授粉;四川牡丹单花花期多为6~8 d,与滇牡丹单花花期[29]接近,较栽培牡丹单花花期(3~6 d)[41]长,这可能也是四川牡丹对高海拔气候条件的一种适应表现。5月份为四川牡丹的盛花期,川西高原开始进入雨季,导致多数昆虫停止活动,不利于四川牡丹完成传粉;而四川牡丹的自交亲和性高于同属植物矮牡丹和滇牡丹[28-29],这也可能是四川牡丹为适应花期因多雨导致传粉昆虫较少而演化出的一种特性;为维持种群数量,四川牡丹可能选择以较高的自交亲和能力来保证繁殖成功。四川牡丹是牡丹组革质花盘亚组(Subsect.Vaginatae)中尚未得到开发利用的珍贵野生资源[18],其花朵硕大、花色艳丽、叶片纤细且小叶众多[16]、结实能力强,分布于干旱河谷地带,对恶劣环境耐受力强,因此是培育新叶形、高抗性及油用牡丹新品种的优异种质资源。对四川牡丹花粉活力和柱头可授期的掌握,以及通过适合的人工授粉方式提高其结籽率,对牡丹新品种的杂交育种均有重要意义。
致谢: 四川农业大学风景园林学院陈其兵教授、刘佳坤和吴云在实验和论文修改过程中提供了无私的帮助,在此表示衷心感谢!
[1] 何亚平, 刘建全. 植物繁育系统研究的最新进展和评述[J]. 植物生态学报, 2003, 27(2): 151-163.
[2] 张洪芳, 李利强, 刘仲键, 等. 菜粉蝶对两种迁地保护的兰科植物传粉和繁殖成功的作用[J]. 生物多样性, 2010, 18(1): 11-18.
[3] 予 茜, 张彦文, 郭友好. 传粉生物学常用术语释译[J]. 植物分类学报, 2008, 46(1): 96-102.
[4] 宋玉霞,郭生虎,牛东玲,等. 濒危植物肉苁蓉(Cistanchedeserticola)繁育系统研究[J]. 植物研究, 2008, 28(3): 278-282.
[5] 刘 芬, 李全健, 王彩霞, 等. 濒危植物扇脉杓兰的花部特征与繁育系统[J]. 林业科学, 2013, 49(1): 53-60.
[6] 李嘉珏. 中国牡丹品种图志: 西北·江南·江南卷[M]. 北京: 中国林业出版社, 2006: 8.
[7] 王莲英, 袁 涛, 王 福, 等. 中国芍药科野生种迁地保护与新品种培育[M]. 北京: 中国林业出版社, 2013: 8-22.
[8] HONG D Y. Note onPaeoniadecompositaHand.-Mazz.[J]. Kew Bulletin, 1997, 52: 957-963.
[9] HONG D Y.Paeoniarotundiloba(D. Y. Hong) D. Y. Hong: a new status in tree peonies (Paeoniaceae)[J]. Journal of Systematics and Evolution, 2011, 49: 464-467.
[10] 马 莘, 夏 颖, 马庆庆, 等. 四川牡丹群落物种生态位及空间关联度分析[J]. 重庆师范大学学报: 自然科学版, 2011, 28(4): 57-61.
[11] 景新明, 郑光华, 裴彦龙, 等. 野生紫斑牡丹和四川牡丹种子萌发特性及与其致濒的关系[J]. 生物多样性, 1995, 3(2): 84-87.
[12] JING X M, ZHENG G H. The characteristics in seed germination and dormancy of four wild species of tree peonies and their bearing on endangerment[J]. Acta Phytophysiologica Sinica, 1999, 25: 214-221.
[13] 宋会兴, 刘光立, 高素萍, 等. 四川牡丹种子浸提液内源抑制物活性初探[J]. 园艺学报, 2012, 39(2): 370-374.
[14] SONG H X, LIU G L, CHEN Q B. Effects of aqueous extracts ofPaeoniadecompositaseeds on germination and some metabolic activities associated with growth of wheat seedlings[J]. Acta Biologica Hungarica, 2012, 63: 362-371.
[15] 杨 勇, 刘光立, 宋会兴, 等. 四川牡丹胚乳浸提液对油菜种子萌发与幼苗生长的影响[J]. 西南农业学报, 2013, 26(1): 89-92.
[16] 刘光立. 四川牡丹(Paeoniadecomposita)自然居群遗传多样性研究[D]. 成都: 四川农业大学风景园林学院, 2013: 38-61.
[17] YUANJH,CORNILLEA,GIRAUDT,et al. Independent domestications of cultivated tree peonies from different wild peony species[J]. Molecular Ecology, 2014, 23: 82-95.
[18] ZHOU S L, ZOU X H, ZHOU Z Q, et al. Multiple species of wild tree peonies gave rise to the ‘king of flowers’,PaeoniasuffruticosaAndrews.[J]. Proceedings of the Royal Society B: Biological Sciences, 2014, 281: 1687.
[19] 成仿云, 李嘉珏, 陈德忠. 中国野生牡丹自然繁殖特性研究[J]. 园艺学报, 1997, 24(2): 180-184.
[20] DAFNI A. Pollination Ecology: A Practical Approach[M]. New York: Oxford University Press, 1992: 21-35
[21] 刘林德, 张洪军, 祝 宁, 等. 刺五加花粉活力和柱头可授性的研究[J]. 植物研究, 2001, 21(3): 375-380.
[22] 张文标, 金则新. 濒危植物夏蜡梅花部综合特征与繁育系统[J]. 浙江大学学报: 理学版, 2009, 36(2): 204-210.
[23] CRUDEN R W. Pollen ovule ratios: a conservative indicator of breedingsystemsinflowering plants[J]. Evolution, 1977, 31: 32-46.
[24] 葛 颂. 植物群体遗传结构研究的回顾和展望[M]∥李承森. 植物科学进展: 第1卷. 北京: 高等教育出版社, 1998: 1-8.
[25] TURPIN R A, SCHLISING R A. A new method for studying pollen dispersal using iodine-131[J]. Radiation Botany, 1971, 11: 75-78.
[26] SCHLISING R A . Reproductive proficiency inPaeoniacalifornica(Paeoniaceae)[J]. American Journal of Botany, 1976, 63: 1095-1103.
[27] 红 雨, 刘 强. 芍药的传粉生物学研究[J]. 广西植物, 2006, 26(2): 120-124.
[28] 罗毅波, 裴颜龙, 潘开玉, 等. 矮牡丹传粉生物学的初步研究[J]. 植物分类学报, 1998, 36(2): 134-144.
[29] 李 奎, 郑宝强, 王 雁, 等. 滇牡丹的开花特征及繁育系统[J]. 东北林业大学学报, 2013, 41(1): 63-67.
[30] BERNHARDT P, MEIER R, VANCE N. Pollination ecology and floral function of Brown’s peony (Paeoniabrownii) in the Blue Mountains of northeastern Oregon[J]. Journal of Pollination Ecology, 2013, 11: 9-20.
[31] WRIGHT G A, SKINNER B D, SMITH B H. Ability of honeybee,Apismellifera, to detect and discriminate odors of varieties of canola (BrassicarapaandBrassicanapus) and snapdragon flowers (Antirrhinummajus)[J]. Journal of Chemical Ecology, 2002, 28: 721-740.
[32] 李绍文. 生态生物化学[M]. 北京: 北京大学出版社, 2001: 2.
[33] 王若涵. 木兰属生殖生物学研究及系统演化表征探析[D]. 北京: 北京林业大学生物科学与技术学院, 2010: 29-36.
[34] 金 红, 焦根林, 陈 刚. 白花野牡丹的开花进程、花部形态特征及访花昆虫观察[J]. 植物资源与环境学报, 2015, 24(2): 73-79.
[35] 曾 荣, 曾爱平, 游兰韶, 等. 中国东方蜜蜂亚种学名订正及生物地理学研究[J]. 中国农学通报, 2012, 28(20): 17-23.
[36] 杨冠煌. 中国蜜蜂资源调查[J]. 中国养蜂, 1984(3): 4-7.
[37] 肖宜安, 何 平, 李晓红. 濒危植物长柄双花木开花物候与生殖特性[J]. 生态学报, 2004, 24(1): 14-21.
[38] 柴胜丰, 韦 霄, 蒋运生, 等. 濒危植物金花茶开花物候和生殖构件特征[J]. 热带亚热带植物学报, 2009, 17(1): 5-11.
[39] 律春燕. 黄牡丹野生种与牡丹、芍药栽培品种远缘杂交研究[D]. 北京: 中国林业科学研究院林业研究所, 2010: 21-23.
[40] 伍翔宇, 陈 敏, 王其刚, 等. 中甸刺玫和川滇蔷薇的繁育系统比较研究[J]. 园艺学报, 2014, 41(10): 2075-2084.
[41] 李嘉珏, 张西方, 赵孝庆, 等. 中国牡丹[M]. 北京: 中国大百科全书出版社, 2011: 78.
(责任编辑: 张明霞)
Studies on floral characteristics and breeding system ofPaeoniadecomposita
YANG Yong1a,1b, LUO Jintao1a, ZHANG Bifang1a, SONG Huixing1a, LIU Guangli1a,①, ZENG Xiuli2,①
(1. Sichuan Agricultural University: a. College of Landscape Architecture, b. Maize Research Institute, Key Laboratory of Biology and Genetic Improvement of Maize in Southwest Region, Ministry of Agriculture, Chengdu 611130, China; 2. Institute of Vegetables, Tibet Academy of Agricultural and Animal Husbandry Sciences, Lhasa 850032, China),
J.PlantResour. &Environ., 2015, 24(4): 97-104
Floral characteristics and flowering dynamic ofPaeoniadecompositaHand.-Mazz. distributed in Aba of Sichuan were observed, and its breeding system was researched through pollen viability analysis, outcrossing index (OCI) estimation, pollination measure control and flower visiting insect observation. The main floral characteristics ofP.decompositaare as follows: flower color from pink to reddish purple, and turning to pinkish white with prolonging of opening time; radialized corolla with 8-12 petals; 128-203 polyadelphous stamens with white filament and yellow anther, having petal phenomenon in stamens; yellow stigma on the top of carpel; border placenta, 2-7 carpels, 8-33 ovules per carpel; aroma spreading during flowering process and changing with prolonging of flowering period. The flowering period of populations of this species is concentrated from the middle of April to the middle of May, but varies with years. The flowering period of solitary flower mostly is 6-8 d. Anther and stigma are basically at the same plane during whole flowering process, and stamens are protandrous. From the first day to the sixth day of flowering period, pollen viability ofP.decompositaincreases firstly and then decreases, in which pollen viability (about 80%) is higher from the third day to the fifth day. Stigma has receptivity generally from the second day of flowering period with duration time of 9 d, and stigma receptivity is the strongest from the fifth day to the eighth day of flowering period. And overlap time of pollen with viability and stigma with receptivity is 5 d. OCI value ofP.decompositais 4, and pollen-ovule ratio (P/O) is 207 220.P.decompositahas no nectary, pollen is the only reward for visiting insects. And the main visiting insects include Apidea, Syrphidae and Cetoniinae, in which the former two insects may be the effective pollinators, while Cetoniinae has low pollination efficiency. The result of pollination experiment shows that average of seed setting rate of natural pollination plant in 2012 and 2014 is the highest, with 23.76%, and that of artificial xenogamy and artificial geitonogamy plants is lower, with 18.33% and 11.04%, respectively. It is suggested that breeding system ofP.decompositais xenogamy, part self-compatibility and no apomixes, and needs pollination by pollinators.
PaeoniadecompositaHand.-Mazz.; floral characteristics; flowering process; visiting insect; pollination measure; breeding system
2015-03-16
国家农业科技成果转化资金项目(2011GB2F000016); 四川省教育厅重点课题(201ZA0009); 西藏自治区科学技术厅重点项目〔藏财企指(专)字[2011]48号〕
杨 勇(1990—),男,河南驻马店人,硕士研究生,主要从事芍药属野生资源开发与利用方面的研究。
①通信作者 E-mail: liugl_1@163.com; zeng_xiuli2004@aliyun.com
Q944.43; Q948.12+2.6; S685.11
A
1674-7895(2015)04-0097-08
10.3969/j.issn.1674-7895.2015.04.13
猜你喜欢
发明与创新·中学生(2023年9期)2023-09-21 09:19:02
中国科学探险(2022年8期)2022-12-31 07:27:54
疯狂英语·新悦读(2022年12期)2022-03-08 02:37:12
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25 01:40:26
课外生活(小学1-3年级)(2020年2期)2020-03-09 03:24:26
铜仁学院学报(2018年7期)2018-09-08 08:50:40
现代园艺(2017年22期)2018-01-19 05:06:57
中国稻米(2017年6期)2017-12-28 02:13:01
中国绿色画报(2016年9期)2017-05-31 08:04:53
作品(2015年5期)2015-11-18 03:20:02
