
珠海淇澳岛无瓣海桑人工林更新幼苗种群特征
2015-06-23 16:22缪绅裕陶文琴曾庆昌龙连娣陈健辉易祖盛
广州大学学报(自然科学版) 2015年1期
安 东,缪绅裕,陈 蔚,陶文琴,曾庆昌,龙连娣,陈健辉,易祖盛
(1.广东珠海淇澳-担杆岛省级自然保护区管理处,广东珠海 519000;2.广州大学生命科学学院,广东广州 510006)
珠海淇澳岛无瓣海桑人工林更新幼苗种群特征
安 东1,缪绅裕2*,陈 蔚1,陶文琴2,曾庆昌2,龙连娣2,陈健辉2,易祖盛2
(1.广东珠海淇澳-担杆岛省级自然保护区管理处,广东珠海 519000;2.广州大学生命科学学院,广东广州 510006)
采用样方法,调查研究了珠海淇澳二斜湾5 a的无瓣海桑人工林林缘或林窗的自然繁殖的无瓣海桑幼苗的种群特征,为了解无瓣海桑自然种群的生态适应性或潜在的生态危害提供参考.结果显示,二斜湾的无瓣海桑幼苗种群总个体数为299株,平均密度7.475株·m-2;其中当年生植株比例占72.24%,上年生植株占27.76%;基径和株高结构显示种群呈增长种群模式,基径和株高呈现极显著的线性相关关系,无瓣海桑幼苗种群为集群分布型.由于受人工林遮蔽光线等的限制和滩涂生长地的竞争,其它自然分布的红树植物如秋茄等,甚至外来入侵的互花米草均受其影响,提示无瓣海桑对其他植物的生长存在生态危害,建议对已种植林分进行乡土红树植物的改造,未种植海滩不再种植无瓣海桑.
无瓣海桑;自然更新幼苗;种群;特征;珠海淇澳岛
海桑科乔木无瓣海桑(Sonneratia apetala),天然分布于亚洲沿海和东太平洋群岛,1985年自孟加拉国引入中国海南东寨港种植[1],1993年引至湛江市沿海[2],2000年后作为广东沿海滩涂主要造林树种广为种植[3].对无瓣海桑的研究,前期侧重于引种扩种,因其生长迅速,林冠大遮蔽强,具有较强的化感效应,引起人们对其生态入侵性的重视,研究重点逐渐转移至生态安全评估[4-11]等方面.XIN等研究认为,海南岛无瓣海桑幼苗的传播不会影响其他红树植物幼苗种群建成,无瓣海桑作为入侵种的证据不足[12].LI研究认为无瓣海桑在控制互花米草(Spartina alterniflora)生长过程中,其化感作用的意义重大[13].LU等则认为,在亚热带天然红树林中种植无瓣海桑不是增加碳固存的有效途径,对天然红树植物和底栖动物存在负面影响[14].
综上所述,无瓣海桑是否已成为沿海红树林生态系统的入侵种仍未有定论.国内部分学者认为无瓣海桑不是入侵物种,其主要理由是无瓣海桑种子不会大量萌发,难以形成自然更新的幼苗种群.然而在珠海淇澳岛的二斜湾,无瓣海桑人工种植5 a后的2014年7月,笔者在林缘及林内空隙处发现有一定规模的自然繁殖当年生幼苗和次年生幼苗.为此对该范围内的无瓣海桑幼苗种群特征进行调查研究,为了解这些幼苗作为更新后代的生存状况,以期为深入了解幼苗的生长特征、生态适应性(生态位)、种苗扩散机制和成为沿海红树林生态系统入侵种的可能性提供科学依据.
1 研究区概况和研究方法
1.1 研究区概况
广东珠海淇澳-担杆岛省级自然保护区建立于2004年,淇澳岛地处东经113°36′~113°39′,北纬22°23′~22°27′,相当于珠江出海口之咽喉.保护区的淇澳岛部分绿地面积约400 hm2,其中红树林面积约占3/4.研究区二斜湾位于珠海淇澳岛北部,其地理位置见图1.二斜湾呈东西走向(东端位于东经113°38′48.81″,北纬22°26′09.37″;西端位于东经113°38′17.05″,北纬22°26′19.83″),长度约900 m.湾的东面(占总长的2/3)宽度280 m;近西位置(占总长的1/3)滩涂面变窄,宽度55 m.湾内泥滩面积约19 hm2,有林面积为10.5 hm2,淤泥地质,淤泥深度40~50 cm.2008年年初广东境内的冰冻灾害,导致二斜湾原人工种植的海桑(Sonneratia caseolaris)全部冻死,于2009年6月改种无瓣海桑.目前无瓣海桑树高6~12 m,平均胸径约15 cm,每年春、秋2季开花结果.
图1 研究地点示意图[15]Fig.1 Sketch map of research site
1.2 调查方法
二斜湾的无瓣海桑人工林下,由种子自然萌发的无瓣海桑幼苗生长最近的幼苗为离岸边3 m,最远达138 m.调查发现无瓣海桑幼苗的分布呈现东面从吊颈石山边向西到湾的中部580 m,出现很多;湾中间的150 m范围内数量很少;接近西面170 m幼苗数量逐步增多.无瓣海桑幼苗呈片状分布,在互花米草集中的区域,无瓣海桑也有分布. 2014年7月进行野外调查,采用样方法,用1 m× 1 m标准方框(因本研究仅涉及幼苗,相当于森林中的林下灌木层),记录样方内每个植物个体的种类、数量;用数显的游标卡尺测定无瓣海桑幼苗基径,用标杆测定植株高度.用年轮法验证幼苗的年龄为当年生或上年生.为了解本地自然生长的秋茄(Kandelia obovata)和外来入侵的互花米草与无瓣海桑在林缘或林隙的生长竞争状况,调查中也包括了少量没有无瓣海桑幼苗分布的样方,样方总数40个,按照从陆地到海洋方向设4个样带,每个样带各有10个标准样方,各样带间相距约200 m.
1.3 统计分析方法
根据野外调查数据,对幼苗基径按每间隔2 mm为1个单位、植株高度按每10 cm为1个单位,在Excel中绘制无瓣海桑幼苗种群的基径、植株高度结构图,对其实际测定数据进行统计拟合,作曲线图;分析基径与植株高度的相关性.种群分布格局的测定按方差V/均测定值m比值的t检验方法,计算公式[16]:C=V/m.若C>1,为聚集分布;C<1,为随机分布;C=1,则为均匀分布[17].式中:C为偏离指数,V为样本方差,m为样本平均数,可用t检验该值的显著性.
2 结果与讨论
2.1 样方内植物种类及其数量与高度
对二斜湾无瓣海桑人工林林缘及林窗40个样方调查的植物种类结果见表1.表1中可见,无瓣海桑幼苗共299株,出现于其中的30个样方(占全部40个样方的75.0%),平均密度7.475株·m-2;互花米草共2 312株,出现于其中的39个样方(占全部样方的97.5%),平均密度57.8株· m-2;秋茄出现于其中的3个样方中,占全部样方的7.5%,平均密度0.075株·m-2.由于互花米草为丛生性草本,其平均密度远大于木本的无瓣海桑和秋茄.
1999年起,淇澳岛就开始种植大片无瓣海桑,用以恢复海滩植被和控制互花米草的危害.对淇澳岛(非二斜湾范围)的无瓣海桑幼苗的研究发现,在水沟两旁密度为2.5株·m-2,林缘光滩为0.5·m-2,互花米草内为0.22株·m-2,无瓣海桑林内仅0.06株·m-2,且这些扩散苗绝大多数为当年生幼苗,几乎未见多年生小苗,极难更新成林[6].本研究中,二斜湾范围内无瓣海桑幼苗的平均密度7.475株·m-2大于水沟旁的2.5株·m-2,但略小于另一近期研究提及林缘的12.5株·m-2[9].
2.2 无瓣海桑幼苗种群植株的基径与高度结构
无瓣海桑幼苗种群的基径结构见图2.图2中可见,基径在0.01~2.00 mm和0.21~0.40 mm的个体数量分布为115株和83株,合计198株,占种群总个体数299株的66.22%,种群的基径结构为种群渐近增长型.种群株高结构见图3,也明显呈种群渐近增长型,其中株高11~20 cm和21~30 cm的个体数分别为98株和86株,合计184株,占整个种群的61.54%.图2和图3在图形外观上很相似,二者结合观察,无瓣海桑幼苗种群整体上呈渐进增长型.一般在生态学上,把幼苗株高小于33 cm定为I级幼苗;因调查时间为7月,种子一般为春季萌发,其生长季不足3个月,从本研究的年轮分析结果看,当年生幼苗的高度均小于40 cm,因而把图3中株高大于40 cm的幼苗定为上年生幼苗,这样当年生幼苗的个体数为216株,上年生幼苗83株,分别占总数的72.24%和27.76%.
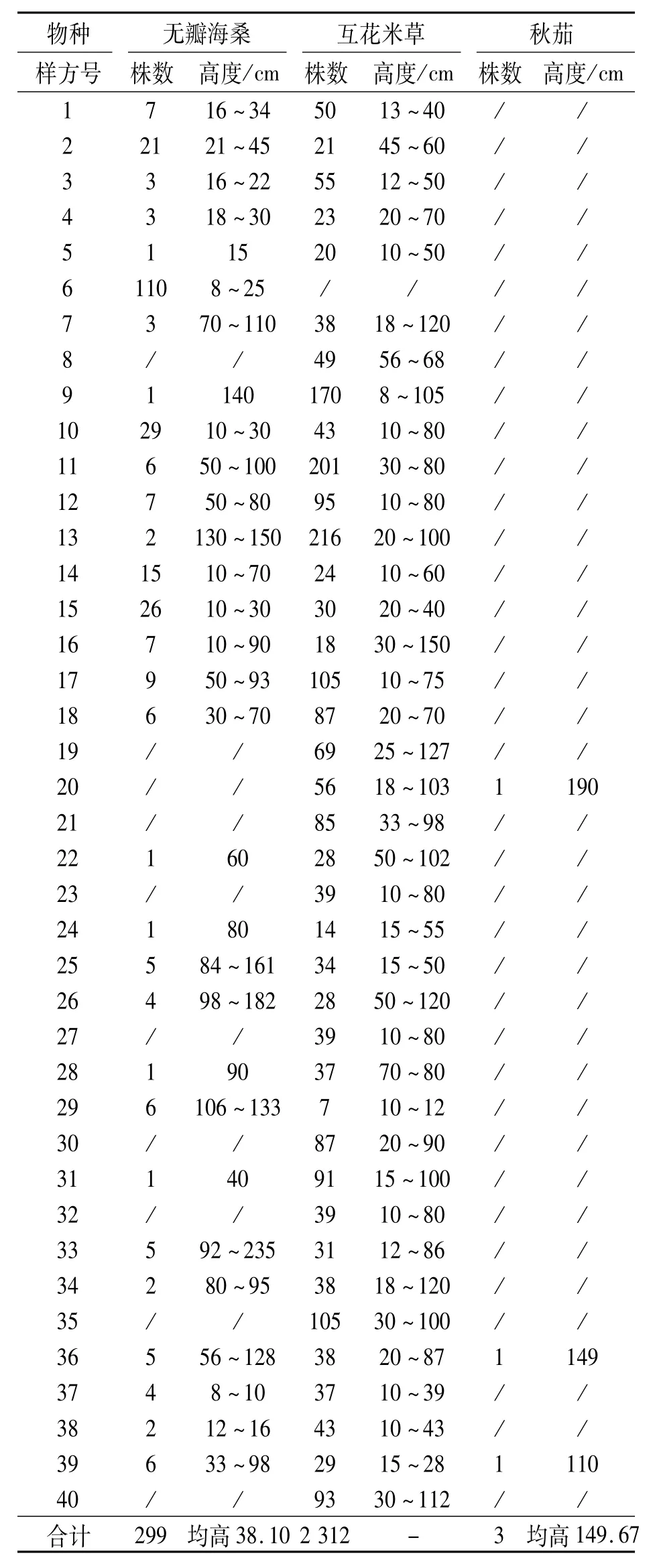
表1 珠海淇澳岛二斜湾无瓣海桑人工林下植物分布状况Table 1 Plant distribution status under the man-made Sonneratia apetala forest at Erxiewan,Qi'ao island,Zhuhai

图2 无瓣海桑幼苗种群的基径结构Fig.2 Base diameter structure of S.apetala seedling population
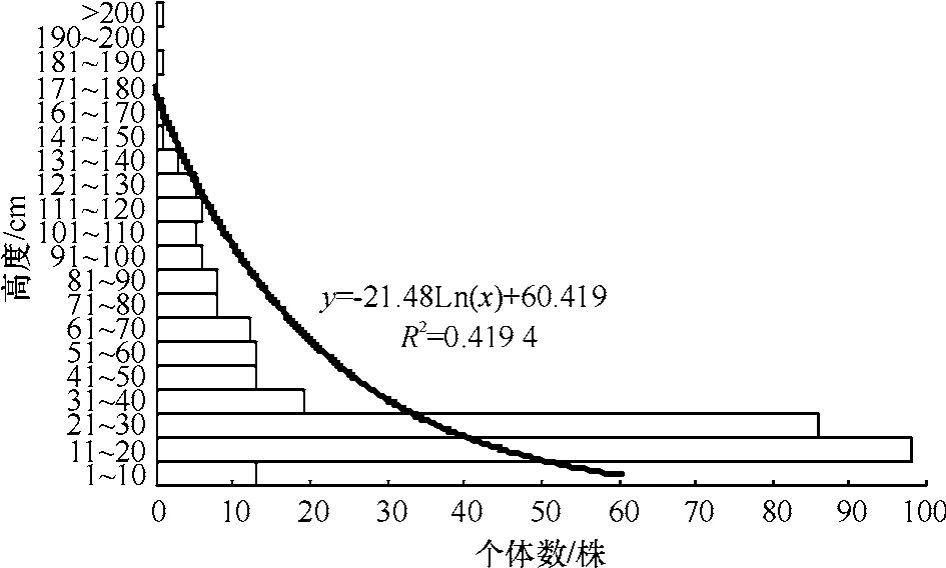
图3 无瓣海桑幼苗种群的株高结构Fig.3 Height structure of S.apetala seedling population
图2中,通过对具体数据的对数拟合曲线,可见相关系数r=0.917 4(自由度df=12),大于置信水平0.01上差异显著的临界值r=0.661 4;图3中,对数拟合曲线的相关系数r=0.647 6(自由度df=18),大于置信水平0.01上差异显著的临界值r=0.561 4.
2.3 无瓣海桑幼苗种群植株的基径与高度的相关性
为进一步探讨无瓣海桑幼苗生长的规律,对无瓣海桑幼苗的基径和株高相关性进行分析,结果见图4.图4显示,基径和株高的线性相关系数r=0.981 4,达到极显著水平(P<0.001),即幼苗生长初期,基径与高度生长极其相关.海南岛东寨港2年生无瓣海桑年生长规律研究结果表明,无瓣海桑树高生长速度与地径生长速度表现并不相一致,树高以4~6月为高峰期(地径生长高峰期则为7~9月)[4],随着无瓣海桑幼苗的生长,这种植株基径与株高的相关性将有可能变得不显著.
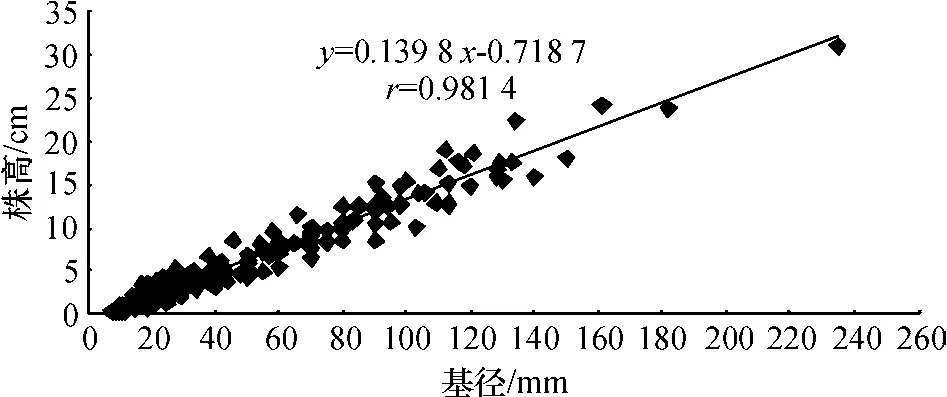
图4 无瓣海桑幼苗基径与株高的相关性Fig.4 Relationship between the base diameter and height of S.apetala seedlings
2.4 无瓣海桑幼苗种群的分布格局
根据C=v/m计算公式,所得出的计算结果见表2.表2中,无瓣海桑和互花米草种群均属于聚集分布,秋茄种群则为随机分布.无瓣海桑种群的聚集分布与大亚湾的桐花树(Aegiceras corniculatum)、木榄(Bruguiera gymnorhiza)、海漆(Excoecaria agallocha)、老鼠簕(Acanthus ilicifolius)的聚集分布一致,主要是红树植物以胎生为其重要的生态特征之一,母树附近的后代个体数较多,从而形成聚集分布型[17].本研究中的秋茄种群为随机分布,与大亚湾秋茄种群的聚集分布有所区别,而与大亚湾在高度和盖度上比其它种更具一定优势的白骨壤(Avicennia marina)种群的随机分布较一致,原因主要是调查不以秋茄为主要目的种,该随机分布格局并不能代表二斜湾范围内秋茄种群的总体分布格局.
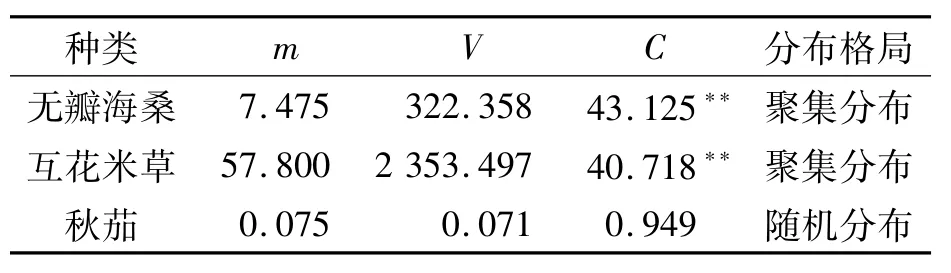
表2 无瓣海桑、互花米草和秋茄的种群分布格局Table 2 Population distributive patterns of Sonneratia apetala,Spartina alterniflora,and Kandelia obovata
3 讨 论
在珠海淇澳二斜湾人工种植的无瓣海桑林附近,目前仅在岸边可见其它少量红树植物,如老鼠簕、秋茄(29丛,每丛由1~5棵组成,高度110~170 cm)、桐花树(5丛,在西面开阔区域,有果实);卤蕨(Acrostichum aureum)仅1处(低矮).以往研究发现,无瓣海桑人工林内自然更新主要以老鼠簕和卤蕨为主.9年龄和12年龄无瓣海桑人工林内出现大量老鼠簕,地势较髙地段有少量卤蕨,但其高度不超过2 m,且未见乔木树种的定居[10].
无瓣海桑作为木本植物,生长快,为速生树种,群落的生产力水平在我国红树林群落中最高.从幼苗到开花结果仅需3 a时间,每年开花结实2次,即秋果和春果,以秋果为主.深圳湾春果为6~7月成熟,秋果为10~11月成果.随着纬度的增加,无瓣海桑的结果期相应推迟[4].2009年6月在二斜湾种植后,2012年10月挂果(秋果),到2013年出现幼苗也只经历了4 a时间,至2014年7月其幼苗个体已增至一定数量,种群形成一定规模,并在滩涂上和互花米草等混生在一起.由于其生长迅速,很可能在较短时间内形成林冠遮蔽,从而导致其他红树植物或互花米草等因光线不足而难以维持生长.1999年以来在淇澳岛种植了大片无瓣海桑人工林,因种植时间较长,其自然繁殖的幼苗已长成8 m以上的大树,占据了原先秋茄、互花米草的生境,呈现出较明显的生态入侵性.
研究认为无瓣海桑具入侵特征的主要理由有:速生、高生产力、对低温(成熟个体半致死温度-1.945℃[9])和土壤适应性强;具较强的化感效应;人工林在自然条件下已开始扩散;本地红树植物难以进入无瓣海桑成林区,几乎没有本地乡土红树植物幼苗能存活于林下;认为引进乡土红树老鼠簕、桐花树也难以在其林下自然更新.由于无瓣海桑的入侵,3 a内各地乡土红树植物的密度变化如下:深圳湾2 812棵·hm-2降低为37棵· hm-2;湛江2 473棵·hm-2降低为51棵·hm-2;海南琼山2 899棵·hm-2降低为111棵·hm-2[5].低纬度地区的无瓣海桑具有更强的传播能力,即具更强的潜在入侵性[11].目前在广东、海南、广西、福建、香港和澳门均发现无瓣海桑的自然扩散和传播的现象.
认为无瓣海桑不具备入侵特性的理由主要有:自然扩散靠种子繁殖,种子量虽大,但萌发率低(盐度0‰~10‰条件下)、定居特点方面有较大限制,不能进行自我更新,只在局部的适宜生境内扩散,不足以造成大规模爆发.其生态位宽度值仅处于中等水平,比中低潮滩的桐花树、秋茄、白骨壤、木榄低,比中高潮的乡土植物角果木(Ceriops tagal)、红海榄(Rhizophora stylosa)、海莲(Bruguiera sexangula)高,与同属植物海桑的较接近.无瓣海桑与桐花树、秋茄、白骨壤、木榄重叠较大;资源匮乏时,无瓣海桑主要与中低潮植物发生较激烈的竞争作用.而淇澳岛自然分布的种类恰好就是桐花树、秋茄、木榄等,因此无瓣海桑对淇澳岛红树林的发展存在较大风险.
本研究认为,无瓣海桑在淇澳二斜湾已出现一定规模的自然更新幼苗种群,呈现出较明显的入侵性.究其原因,主要是二斜湾的生境条件十分符合无瓣海桑种子的萌发.整个二斜湾淤泥深厚,达40~50 cm、松软且肥沃;位置避风,温度常年在0℃以上;受到西江淡水影响,此地咸淡水盐度较低,多在0‰~10‰.如果进一步观察,整个珠江河口的海岸生境都适宜无瓣海桑自然生长繁殖,然后与其他红树植物竞争,侵占生境.
如何用适用密度的乡土红树植物种植,来控制该生境中互花米草种群的恢复,尚有待于进一步研究.
无瓣海桑人工林内幼苗较少,主要原因是幼苗生长同样需要较强的阳光,结合珠海淇澳的红树林改造,在无瓣海桑人工林下种植木榄、桐花树、秋茄,不能只满足于它们存活(营养生长),更重要的是看它们能否开花、结果、繁殖.二斜湾的秋茄多数现今只存活不开花,而在阳光充足之处可开花结果.所以林分改造,应砍伐一定数量的无瓣海桑,降低郁闭度,阳光充足后,进入的乡土树种才可以正常生长.
鉴于目前无瓣海桑在国内的生长态势和扩散趋势,需引起沿海各红树林保护区的高度重视和警惕,建议在未种植区域停止人工种植,在已种植区域加强管理,及时清除自然更新幼苗,用乡土红树植物替代种植;同时加强科学研究,例如,对无瓣海桑幼苗个体的动态、静态生命表等的进一步研究,可深入了解无瓣海桑的扩散机制,否则在沿海看到的将会是单一物种的红树林生态系统.
[1] 廖宝文,张乔民.中国红树林的分布、面积和树种组成[J].湿地科学,2014,12(4):435-440. LIAO B W,ZHANG Q M.Area,distribution and species composition of mangroves in China[J].Wetland Sci,2014,12(4):435-440.
[2] 廖宝文,郑松发,陈玉军,等.海南东寨港几种国外红树植物引种初报[J].中南林学院学报,2006,26(3):63-67. LIAO B W,ZHENG S F,CHEN Y J,et al.Preliminary report on introduction of several alien mangrove plants in Dongzhai Harbour of Hainan Province[J].J Centr South Forest Univ,2006,26(3):63-67.
[3] 缪绅裕,李德宁,邓鸿英,等.广州南沙3种红树植物生理生态特性的比较[J].广州大学学报:自然科学版,2011,10(1):31-36. MIAO S Y,LI D N,DENG H Y,et al.Comparison of some eco-physiological characteristics of three mangrove species in Nansha District,Guangzhou[J].J Guangzhou Univ:Nat Sci Edi,2011,10(1):31-36.
[4] 廖宝文,郑松发,陈玉军,等.外来红树植物无瓣海桑生物学特性与生态环境适应性分析[J].生态学杂志,2004,23(1):10-15. LIAO B W,ZHENG S F,CHEN Y J,et al.Biological characteristics and ecological adaptability for non-indigenous mangrove species Sonneratia apetala[J].Chin J Ecol,2004,23(1):10-15.
[5] REN H,LU H F,SHEN W J,et al.Sonneratia apetala Buch.Ham in the mangrove ecosystems of China:An invasive species or restoration species?[J].Ecol Engin,2009,35:1243-1248.
[6] 廖宝文,田广红,杨雄邦,等.珠海淇澳岛无瓣海桑种苗天然更新与扩散分析[J].生态科学,2006,25(6):485-488. LIAO B W,TIAO G H,YANG B X,et al.The analysis of natural regeneration and diffusion of the seedling of Sonneratia apetala in the Qi'ao Island,Zhuhai[J].Ecol Sci,2006,25(6):485-488.
[7] 田广红,陈蕾伊,彭少麟,等.外来红树植物无瓣海桑的入侵生态特征[J].生态环境学报,2010,19(12):3014-3020. TIAN G H,CHEN L Y,PENG S L,et al.Ecological traits of invasiveness of alien mangrove species Sonneratia apetala[J]. Ecol Environ Sci,2010,19(12):3014-3020.
[8] 彭友贵,徐正春,刘敏超.外来红树植物无瓣海桑引种及其生态影响[J].生态学报,2012,32(7):2259-2270.PENG Y G,XU Z C,LIU M C.Introduction and ecological effects of an exotic mangrove species Sonneratia apetala[J].Acta Ecol Sin,2012,32(7):2259-2270.
[9] 朱宏伟.珠海淇澳岛无瓣海桑入侵风险综合评估[D].北京:中国林业科学研究院,2012. ZHU H W.Sonneratia apetala invasion risk comprehensive assessment in Qi'ao Island,Zhuhai[D].Beijing:Chinese Academy of Forestry,2012.
[10]刘滨尔.淇澳岛无瓣海桑人工林的自然更新特征及林分改造效果[D].北京:中国林业科学研究院,2013. LIU B E.The features of natural regeneration of Sonneratia apetala plantation and results of its stand improvement on Oi'ao Island[D].Beijing:Chinese Academy of Forestry,2013.
[11]文玉叶.不同纬度无瓣海桑的繁殖和扩散特性研究[D].厦门:厦门大学,2014. WEN Y Y.Studies on propagation and diffusion properties of Sonneratia apetala in latitude gradient[D].Xiamen:Xiamen University,2014.
[12]XIN K,ZHOU Q,ARNDT S K,et al.Invasive capacity of the mangrove Sonneratia apetala in Hainan Island,China[J].J Tropic Forest Sci,2013,25(1)70-78.
[13]LI J,PENG S L,CHEN L Y,et al.Use of Sonneratia apetala allelopathy to control Spartina alterniflora weed[J].Allelop J,2010,25(1):123-131.
[14]LU W Z,YANG S C,CHEN L Z,et al.Changes in carbon pool and stand structure of a native subtropical mangrove forest after inter-planting with exotic species Sonneratia apetala[J].Plos One,2014,9(3):4.
[15]百度地图Baidu Map.http:map.baidu.com(2014-11-4)
[16]伊力塔,韩海荣,康峰峰.灵空山林区辽东栎种群空间分布格局[J].生态学报,2008,28(7):3254-3261. YI L T,HAN H R,KANG F F.Spatial distribution patterns of Quercus liaotungensis population in Lingkong Mountains[J]. Acta Ecol Sin,2008,28(7):3254-3261.
[17]陈桂珠,缪绅裕.广东澳头红树植物群落的物种多样性与种群分布格局研究[J].生态学杂志,1994,13(2):34-35. CHEN G Z,MIAO S Y.Species diversity and population distribution pattern of mangrove community Aotou of Guangdong[J].Chin J Ecol,1994,13(2):34-35.
Population characteristics of seedlings under the man-made Sonneratia apetala forest at Qi'ao island,Zhuhai
AN Dong1,MIAO Shen-yu2,CHEN Wei1,TAO Wen-qin2,ZENG Qing-chang2,LONG Lian-di2,CHEN Jian-hui2,YI Zu-sheng2
(1.Administration of Qi'ao and Dangan Island Provincial Nature Reserves,Zhuhai 519000,China;2.School of Life Sciences,Guangzhou University,Guangzhou 510006,China)
The population characteristics of naturally regenerated Sonneratia apetala under the edges and gaps of the five-year man-made S.apetala forest at the Erxiewan of Qi'ao,Zhuhai was surveyed by using the quadrat method,so as to provide basic data for investigating the ecological adaptability and potential damage of natural population of S.apetala.The results showed that the seedling population of S.apetala at Er'xiewan has a total of 299 individuals,with average density of 7.475 individuals·m-2.The current-year seedling accounted for 72.24%,while the last-year seedling took up 27.76%of the whole population.The size structure of base diameter and plant height showed an increasing population type,and there was a significant linear relationship between base diameter and plant height.The population has a clump distribution type.The growth of native mangrove species such as Kandelia obovata,even the invasive plant species Spartina alterniflora were affected by this plantation trees through competing light and habitat,suggesting the Sonneratia apetala has an ecological damage to the other mangrove species.The structure of this plantation forest of S.apetala should be changed with native mangrove species and the S.apetala should not be planted anymore for the bare beaches.
Sonneratia apetala;naturally regenerated seedling;population;characteristics;Qi'ao island
Q 948.15
A
【责任编辑:周 全】
1671-4229(2015)01-0050-06
2014-11-08;
2014-12-08
国家自然科学基金资助项目(31270526)
安 东(1960-),男,高级工程师,硕士.E-mail:2740979771@qq.com
*通信作者.E-mail:272118197@qq.com
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
海洋通报(2021年5期)2021-12-21
广东第二课堂·小学(2021年10期)2021-12-03
世界科学技术-中医药现代化(2021年12期)2021-04-19
能源与环境(2020年4期)2020-09-02
四川林业科技(2019年5期)2019-07-07
安徽农学通报(2019年1期)2019-02-14
湖北林业科技(2019年6期)2019-01-06
湖北林业科技(2017年2期)2017-05-25
