
尖孢镰刀菌古巴专化型4号生理小种fpd1基因敲除与表型分析
2015-05-30 15:41王飞燕郭立佳杨腊英汪军王国芬黄俊生
热带作物学报 2015年8期
王飞燕 郭立佳 杨腊英 汪军 王国芬 黄俊生


摘 要 为了研究尖孢镰刀菌古巴专化型4号生理小种fpd1基因的功能,构建了该基因敲除突变体并对其表型特征进行了分析。结果表明,与野生型菌株相比,fpd1基因敲除突变体生长减慢,产孢量显著降低,对巴西蕉的致病性明显减弱;fpd1基因可能在尖孢镰刀菌古巴专化型的产孢、生长发育和致病性等方面具有重要作用。本研究结果为进一步研究fpd1基因的功能和尖孢镰刀菌的致病机理奠定了基础。
关键词 尖孢镰刀菌古巴专化型;fpd1基因;敲除;致病性;产孢
中图分类号 S482.2 文献标识码 A
Construction and Phenotype Analysis of the fpd1 Deletion Mutants of the Fungal Pathogen Fusarium oxysporum f. sp. Cubense
WANG Feiyan1,2,GUO Lijia2,YANG Laying2,WANG Jun2,
WANG Guofen2,HUANG Junsheng1*
1 Hainan University,Haikou,Hainan 570228,China
2 Environment and Plant Protection Institute,Chinese Academy of Tropical Agricultural Sciences/
Key Laboratory of Monitoring and Control of Tropical Agricultural and Forest Invasive
Alien Pests,Ministry of Agriculture,Haikou,Hainan 571101,China
Abstract To investigate the function of fpd1 in Fusarium oxysporum f. sp. Cubense race 4(Foc4),the fpd1 deletion mutants were constructed and its phenotypic characteristics were analyzed. The results showed that the mutant had a slower growth rate on potato dextrose agar (PDA) plates, and reduced conidiation compared to the wild-type strain. Additionally,the mutants showed decreased pathogenicity to bananas(Musa spp. cv. Brazil). Therefore the fpd1 gene in Foc4 plays an important role on conidiation,growth, pathogenicity. The data lay the foundation for further study on the function of fpd1 gene and the pathogenic mechanism in Foc4.
Key words Fusarium oxysporum f. sp. Cubense;fpd1 gene;Deletion;Pathogenicity;Conidiation
doi 10.3969/j.issn.1000-2561.2015.08.017
香蕉枯萎病又称香蕉黄叶病、香蕉巴拿马病,由尖孢镰刀菌古巴专化型(Fusarium oxysporum f. sp. Cubense)从香蕉根系侵入并造成维管束萎蔫和植株枯萎,是一种典型的土传维管束病害,病原菌主要有1号和4号生理小种[1-4]。该病菌为制约香蕉产业发展的重要病原菌之一,目前尚无理想的化学防治药剂,生产上主要通过选育抗病品种予以防治[5]。
目前,已有许多学者对重要经济作物枯萎病害病原菌尖镰孢的致病基因、毒力基因、致病相关基因等进行了大量的研究[6-12]。香蕉枯萎病菌致病基因主要包括信号转导相关基因、转录因子基因、克服植物防御系统相关基因以及与镰刀菌自身定殖能力相关的基因,如fmk1、ste12 、fow1、fow2和six1等[10-12]。将T-DNA插入突变体库和基因敲除策略是鉴定尖孢镰刀菌的重要致病基因的主要手段[13-15]。
关于尖孢镰刀菌fpd1基因的研究主要集中于番茄专化型,对尖孢镰刀菌番茄专化型 fpd1基因的研究结果表明,其编码的产物部分与非洲爪蟾(Xenopus laevis)的氯离子电导调节蛋白(chloride conductance regulatory protein, ICln)相似,为一种跨膜蛋白,是一个致病性相关基因[16-18]。而尖孢镰刀菌古巴专化型的fpd1基因具体功能则尚未见报道。
香蕉枯萎病原菌古巴专化型4号生理小种的fpd1基因全长为1 013 bp,含有一个47 bp的内含子,编码序列长度为966 bp。与 1 号生理小种相比,4号生理小种fpd1基因全长多24 bp,两者有31个核苷酸碱基位点存在差异,同源性为 99.3%[19]。
笔者采用DNA重组策略敲除香蕉枯萎病菌4号生理小种致病相关基因fpd1,构建该基因敲除突变体,研究其对菌株产孢量、生长和菌落形态等特征及致病力的影响,以完善该基因的具体功能研究,为尖孢镰刀菌的致病机理研究提供基础理论。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 菌株为尖孢镰刀菌古巴专化型(Fusarium oxysporum f. sp. cubense)4号小种菌株B2。质粒为pCT74质粒,由中国热带农业科学院环境与植物保护研究所微生物资源研究利用课题组保存。
1.1.2 培养基及试剂 PDA培养基(马铃薯200 g/L、葡萄糖20 g/L、琼脂18 g/L,不加琼脂为PDB培养基);再生培养基(马铃薯200 g/L、蔗糖1.2 mol/L、琼脂0.9%);STC溶液[山梨醇1.2 mol/L、氯化钙0.5%、Tris-HCl 100 mmol/L(pH7.0)、EDTA 25 mmol/L];PTC溶液[将40%(W/V)PEG4000溶于STC溶液,用过滤器(孔径0.2 μm)进行过滤];0.8 mol/L氯化钠溶液;Driselase崩溃酶溶液(20 mg/mL,Sigma)和Lysing Enzyme溶壁酶(20 mg/mL,广东省微生物研究所),均用0.8 mol/L氯化钠溶液配制。
1.1.3 植物材料 供试香蕉品种为巴西蕉(Musa spp. cv. Brazil),由中国热带农业科学院组织培养中心提供。
1.2 方法
1.2.1 真菌基因组DNA提取 将B2菌株接种于PDB培养基,于180 r/min、28 ℃摇床震荡培养5 d,过滤收集菌丝体用于基因组DNA提取。DNA提取采用CTAB法[20]。
1.2.2 fpd1基因敲除载体构建 根据基因组测序序列分别设计fpd1基因5′和3′端同源臂特异引物对,分别标记为fpd1-AF和fpd1-AR、fpd1-BF和fpd1-BR(表1)。以野生型B2(WT)基因组DNA为模板,分别用特异引物对进行PCR扩增,获得fpd1基因开放阅读框上游5′端和下游3′端同源臂DNA片段,分别标记为A臂和B臂。PCR反应体系如下:模板50 ng,引物浓度均为10 pmol/L,2×Taq PCR master Mix(TIANGEN)12.5 μL,用ddH2O将总体系补足至25 μL。PCR反应条件为:94 ℃ 5 min;94 ℃ 30 s, 55 ℃ 30 s, 72 ℃ 1 min,30个循环;72 ℃ 10min,4 ℃保温。经1.0%琼脂糖凝胶电泳后回收PCR产物。
将PCR产物分别连接于pMD19T载体,构建pMD19T-A和pMD19T-B克隆载体;将克隆载体分别用限制性内切酶KpnⅠ和XhoⅠ、EcoRⅠ和 XbaⅠ组合进行酶切,获得5′端和3′端同源臂DNA,分别先后连接于经相同限制性内切酶线性化的pCT74载体(其中含潮霉素抗性基因hph和GFP基因表达盒)中,构建fpd1基因敲除载体;再用KpnⅠ和XbaⅠ从敲除载体上切下用于转化的DNA片断,即A+hph+GFP+B,回收纯化备用。
1.2.3 尖孢镰刀菌原生质体制备和DNA转化 从新活化的尖孢镰刀菌平板中获取7 mm菌饼,将其接种于PDB培养基中,于28 ℃摇床培养5 d;用灭菌的2层擦镜纸过滤去除菌丝,获得孢子悬浮液(浓度约为107 cfu/mL);吸取2 mL孢子液接种于PDB培养基中,以28 ℃、120 r/min摇床培养9~10 h;用灭菌的6层擦镜纸收集菌丝,并用0.8 mol/L NaCl冲洗菌丝,充分洗涤;挑取黄豆大小(直径约3 mm)的菌丝,加入崩溃酶和溶壁酶溶液各6 mL,以28 ℃、150 r/min消化酶解2~3 h;用灭菌的2层擦镜纸过滤,收集滤液,于室温中以3 500 r/min 离心10 min,弃上清;加30 mL STC重悬沉淀,于室温中以3 500 r/min 离心10 min,弃上清;加入1~2 mL STC, 将原生质体调整至浓度约为107 cfu/mL。
取5 μg上述酶切回收的DNA片段,即A+hph+GFP+B DNA片段,缓慢加到200 μL 原生质体中,轻轻混匀,于室温中静置20 min;缓慢加1~1.25 mL 40% 的PTC溶液至管中,轻轻混匀,于室温中静置20 min;将上述反应液加入100 mL冷却至50 ℃左右的再生培养基(含有100 μg/mL Amp 和50 μg/mL 潮霉素B)中,混匀后,倒于培养皿上;于28 ℃中恒温培养4~7 d 后获得转化子,并连续培养3代获得稳定转化子。
1.2.4 fpd1基因敲除突变体的鉴定 采用CTAB法[20]提取转化子基因组DNA,经紫外分光光度计检测合格后,将其作为PCR扩增的模板;分别以转化子和野生型菌株B2基因组DNA为模板,根据基因组序列设计特异引物对进行PCR扩增检测。其中,引物F1和F2检测转化子是否在fpd1基因开放阅读框5′端发生同源重组;引物F5和F6检测转化子是否在fpd1基因开放阅读框3′端发生同源重组;引物F3和F4检测转化子是否存在fpd1基因开放阅读框,预计扩增长度分别约1.3、1.4、0.8 kb(表1)。如果引物F1和F2、F5和F6 的PCR扩增结果为阳性而引物F3和F4的 PCR扩增结果为阴性,则此转化子即为fpd1基因敲除突变体。PCR反应体系如下:模板100 ng,引物浓度均为10 pmol/L,2×Taq PCR master Mix(TIANGEN)12.5 μL,用ddH2O将总体系补足至25 μL。PCR反应条件为:94 ℃ 预变性5 min;94 ℃变性 40 s, 55 ℃退火 30 s,72 ℃延伸 2 min,共30个循环;72 ℃ 10 min,4 ℃保温。
1.2.5 突变体表型分析 生长速率测定:将敲除突变体与野生型菌株B2 的菌饼(约7 mm)分别接种于PDA平板上培养,每个菌株接种3个重复,5 d后测量菌落直径。
产孢量分析:分别从长有fpd1敲除突变体和野生型菌株B2(均培养了5 d) 的PDA平板上打7 mm的菌饼5个,放入含有20 mL 0.05% 吐温-20的蒸馏水中,慢速摇5 min,然后采用血球计数板计数。
突变体形态显微镜观察:从PDB培养液中吸取少量菌液到载玻片上,制片后在荧光显微镜(FV1000,Olympus)下观察拍照。
1.2.6 致病性测定及统计 选取长势相近的巴西香蕉苗(株高约10 cm,5~6片叶子)种植于一次性塑料碗中;将培养了5 d的fpd1敲除突变体和野生型菌株经过滤离心回收孢子,并用蒸馏水制孢子悬液(约106 cfu/mL),每株苗浇50 mL孢子液,以无菌水浇苗作为对照,每个菌株致病性测定设置3个重复,每个重复取10株幼苗;处理后,按常规方法栽培管理,并观察植株的发病情况;4周后调查病害发生情况,将球茎从中间纵向切开,观察球茎褐变程度,记录每株幼苗发病级数[20]。
1.3 数据处理
采用SAS 9.2分析软件对产孢量、菌落直径大小和病情指数进行显著性分析。香蕉幼苗发病情况用病情指数表示,病情指数计算采用如下公式:
病情指数=100×∑(植株发病级数×对应株数)/(植株发病最高级数×总株数)[20]。
2 结果与分析
2.1 fpd1基因敲除载体构建
对构建的敲除载体pCT74-fpd1进行酶切验证。结果表明,用限制酶KpnⅠ和XhoⅠ可从敲除载体pCT74-fpd1上切下约1 100 bp的DNA条带,即A臂;用限制酶 EcoRⅠ和XbaⅠ可从载体上切下B臂,约1 004 bp;用KpnⅠ和和XbaⅠ限制酶可从载体上切下同源重组片段A+hph+gfp+B,与预计大小一致(图1-A)。同时对载体的A臂和B臂进行PCR扩增鉴定,结果均可扩增到1条1 kb左右的DNA同源臂(图1-B)。将载体进行测序,测序结果表明所克隆的DNA片段正确。这些结果表明fpd1基因敲除载体已构建成功。
2.2 PCR鉴定fpd1 敲除突变体
经再生培养和潮霉素抗性筛选,共获得10个转化子。分别提取野生型菌株B2基因组DNA和转化子基因组DNA,并以之为模板,分别用引物对F1/F2、F3/F4和F5/F6进行PCR扩增(图2-A)。结果显示(图2-B),以其中2个转化子DNA为模板,用引物对F1/F2和F5/F6进行PCR扩增分别获得约1.3、1.4 kb 的DNA条带(泳道1和2,4和5),与预计大小一致,而野生型菌株模板无此PCR扩增条带(泳道7和8)。用引物对F3/F4进行PCR扩增,结果显示,以2转化子基因组DNA和清水为模板均无扩增条带(泳道3、6和10),而以野生型菌株DNA为模板进行PCR扩增,可扩增到约0.8 kb的DNA条带(泳道9)。这些结果表明这2个转化子的fpd1基因已被敲除,故将其分别命名为Δfpd1-1和Δfpd1-2。其中Δfpd1-1的孢子和菌丝荧光显微镜图分别见图2-C、D。
2.3 fpd1敲除突变体表型分析
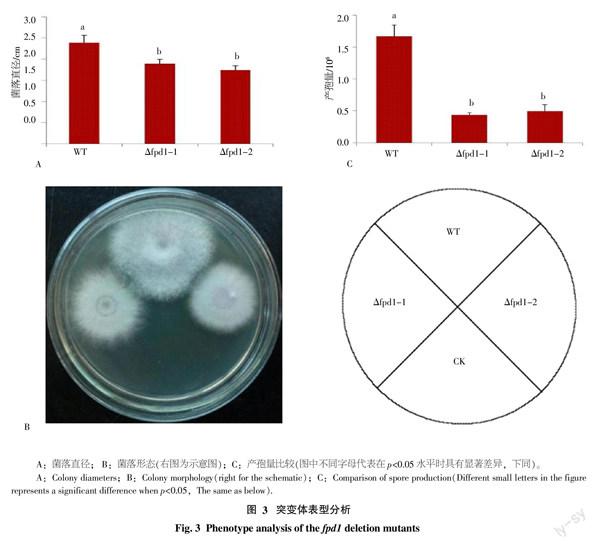
将fpd1基因敲除突变体菌株Δfpd1-1、Δfpd1-2和野生型菌株B2分别接种于PDA平板上并培养5 d,测量其菌落直径。结果表明,野生型B2菌落平均直径(2.39 cm)大于敲除突变体菌株Δfpd1-1和Δfpd1-2的菌落平均直径(1.89 cm和1.74 cm),而且差异极显著(p<0.01)。此外,与野生型相比,突变体的菌落不能形成大量气生菌丝(图3-A、B)。
将Δfpd1-1、Δfpd1-2突变体和野生型菌株B2分别接种于PDA平板上培养5 d,计算单菌落产孢量。结果表明,2个突变体菌株单菌落产孢量(0.44×106和0.5×106)均极显著(p<0.01)低于野生型B2菌株单菌落产孢量(1.67×106,图3-C)。
综合研究表明,fpd1基因敲除突变体菌丝生长减慢,产孢量减少。由此可见,fpd1基因对尖孢镰刀菌4号生理小种的菌丝生长和产孢具有重要的作用。
2.4 fpd1基因敲除突变体对巴西蕉的致病性
将fpd1基因敲除突变体菌株Δfpd1-1、Δfpd1-2与野生型菌株B2分别接种巴西蕉幼苗。根据病情指数统计,结果表明,接种fpd1基因敲除突变体菌株Δfpd1-1和Δfpd1-2的巴西蕉幼苗平均病情指数(14.4和16.2)均极显著(p<0.01)低于野生型菌株B2(61.3)(图4)。由此可见,fpd1基因敲除导致尖孢镰刀菌对巴西蕉的致病性减弱。
3 讨论与结论
氯离子电导调节蛋白(ICln)是非洲爪蟾(Xenopus laevis)膜通道蛋白离子传导孔的一个亚基,功能上与动物溶胀相关的氯离子通道有关[16-17]。Kawabe等[18]研究发现,与非洲爪蟾的氯离子电导调节蛋白相似的番茄专化型FPD1蛋白是致病因子,该基因缺失会导致致病性减弱;而蛋白氨基酸序列比对分析发现,尖孢镰刀菌古巴专化型的FPD1蛋白与非洲爪蟾的氯离子电导调节蛋白序列相似性仅为15.9%,与番茄专化型FPD1蛋白相似性为99.7%。因此,笔者推测尖孢镰刀菌古巴专化型的FPD1蛋白具有与番茄专化型FPD1蛋白相似功能,可能参与尖孢镰刀菌的致病性。
笔者通过基因敲除的策略获得了尖孢镰刀菌古巴专化型4号生理小种的fpd1基因敲除突变体。通过表型分析发现,与野生型菌株相比,fpd1基因敲除突变体孢子产量降低,生长较缓慢,即其在PDA平板上的菌落直径小于野生型(图3),这些结果与古巴专化型的fgb1、ste12、fomps1等基因敲除突变体的研究结果相似[10-12,21]。同时笔者通过致病性测定,发现fpd1基因敲除突变体的致病力明显低于野生型,表明fpd1基因敲除导致其致病力降低,这与郭立佳等[22]报道的尖孢镰刀菌古巴专化型fgb1基因的研究结果相似。综上分析,笔者认为fpd1基因对尖孢镰刀菌古巴专化型的产孢、菌丝生长情况以及致病性等方面具有调控作用。另外,根据尖孢镰刀菌番茄专化型fpd1基因的相关研究结果,FPD1具有氯离子电导调节蛋白部分相似结构,是一种跨膜转运蛋白,因此,笔者推测尖孢镰刀菌古巴专化型可能通过调控离子跨膜运输影响其产孢、菌丝生长情况和致病力。关于fpd1基因突变体致病性减弱的具体机理有待进一步研究。
参考文献
[1] 黄小光, 徐志宏, 邝哲师, 等. 香蕉枯萎病拮抗菌筛选试验[J]. 广东农业科学, 2006(11): 96.
[2] 胡莉莉, 窦美安, 谢江辉, 等. 香蕉枯萎病抗病性研究进展[J]. 广西热带农业, 2006, 8(1): 57-60.
[3] Waite B H, Stover R H. Studies on fusarium wilt of bananas: VI. Variability and the cultivar concept in fusarium oxysporum f. cubense[J]. Canadian Journal of Botany, 1960, 38(6): 985-994.
[4] Su H, Hwang S C, Ko W H. Fusarial wilt of Cavendish bananas in Taiwan[J]. Plant Disease, 1986, 70(9): 814-818.
[5] Michielse C B, Rep M. Pathogen profile update: Fusarium oxysporum[J]. Molecular Plant Pathology, 2009, 10(3): 311-324.
[6] De Ascensao A R, Dubery I A. Panama disease: cell wall reinforcement in banana roots in response to elicitors from Fusarium oxysporum f. sp. cubense race four[J]. Phytopathology, 2000, 90(10): 1 173-1 180.
[7] Visser M, Gordon T R, Wingfield B D, et al. Transformation of Fusarium oxysporum f. sp. Cubense, causal agent of Fusarium wilts of banana, with the green fluorescent protein (GFP)gene[J]. Australasian Plant Pathology, 2004, 56(33): 69-75.
[8]代 易, 纪春艳, 王振中, 等. 香蕉多聚半乳糖醛酸酶抑制蛋白的分离与纯化[J]. 华中农业大学学报, 2011, 30(6): 707-711.
[9] 吴飞宏. 香蕉枯萎病菌4号小种一个致病相关基因克隆及其功能的初步研究[D]. 海口: 海南大学, 2012: 13-26.
[10] Rispail N, Di Pietro A. Fusarium oxysporum Ste12 controls invasive growth and virulence downstream of the Fmk1 MAPK cascade[J]. Molecular Plant-Microbe Interactions, 2009, 22(7): 830-839.
[11] Michielse C B, Van Wijk R, Reijnen L, et al. Insight into the molecular requirements for pathogenicity of Fusarium oxysporum f. sp. lycopersici through large-scale insertional mutagenesis[J]. Genome Biology, 2009, 10(1): R4.
[12] Pietro A D, Madrid M P, Caracuel Z, et al. Fusarium oxysporum: exploring the molecular arsenal of a vascular wilt fungus [J]. Molecular Plant Pathology, 2003, 4(5): 315-325.
[13]蒋艳琴, 曾 涛, 陈汉清, 等. 尖镰孢古巴专化型Focr4-1701突变体的生物学表型研究[J]. 基因组学与应用生物学, 2013, 32(3): 339-346.
[14] Li M H, Xie X L, Lin X F, et al. Functional characterization of the gene FoOCH1 encoding a putative alpha-1,6-mannosyltransferase in Fusarium oxysporum f. sp. cubense[J]. Fungal Genetics and Biology, 2014, 65(1): 1-13.
[15] Qi X Z, Guo L J, Yang L Y, et al. Foatf1, a bZIP transcription factor of Fusarium oxysporum f. sp. cubense, is involved in pathogenesis by regulating the oxidative stress responses of Cavendish banana(Musa spp.)[J]. Physiological and Molecular Plant Pathology, 2013, 84:76-85.
[16] Grigory B K, Michael JA, Eric AG, et al. Molecular characterization of a swelling induced chloride conductance regulatory protein, pICIn[J]. Cell, 1994, 76(3): 439-448
[17] Fürst J, Jakab M, Konig M, et al. Structure and function of the ion channel ICIn[J]. Cell Physiol Biochem, 2000, 10(5-6): 329-334
[18] Kawabe M, Mizutani K, Yoshida T, et al. Cloning of the pathogenicity-related gene FPD1 in Fusarium oxysporum f. sp. Lycopersici[J]. Journal of General Plant Pathology, 2004, 70(1): 16-20.
[19]唐复润. 香蕉枯萎病菌FPD1基因的克隆、表达及其生防菌分离、 筛选[D]. 儋州: 华南热带农业大学, 2007.
[20]郭立佳, 彭 军, 杨腊英, 等. 大蕉枯萎病病原菌的分离、 鉴定和致病性测定[J]. 热带作物学报, 2013, 34(1): 105-110.
[21] 戴青冬. 香蕉枯萎病菌MAPK信号通路中Fomps1基因的研究[D]. 海口: 海南大学, 2013.
[22] 郭立佳, 杨腊英, 王国芬, 等. 尖孢镰刀菌古巴专化型fgb1基因敲除突变体的构建与表型分析[J]. 热带作物学报, 2014, 35(11): 2 205-2 210.
猜你喜欢
山东农业科学(2020年1期)2020-07-04
安徽农业科学(2020年10期)2020-06-08
安徽农业科学(2018年18期)2018-05-14
中成药(2017年12期)2018-01-19
江苏农业科学(2016年4期)2016-06-14
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年12期)2016-05-17
兽医导刊(2015年5期)2016-01-04
华南农业大学学报(2015年5期)2015-12-04
江苏农业科学(2014年9期)2014-11-15
