
不同坡位野生三裂绣线菊和黄刺玫成年植株的叶特性
2015-05-05 00:59王芳刘宁
中国水土保持科学 2015年3期
王芳,刘宁
(山西农业大学林学院,030801,山西太谷)
不同坡位野生三裂绣线菊和黄刺玫成年植株的叶特性
王芳,刘宁†
(山西农业大学林学院,030801,山西太谷)
以同一坡面不同坡位上生长的野生三裂绣线菊和黄刺玫成年植株为试验材料,于2013和2014年连续2个生长季内,通过监测上、中、下3个坡位上土壤水分含量的变化趋势,测定土壤养分质量分数以及2种灌木的叶形态及叶营养等指标,研究3个坡位上生长的2种灌木成年植株的叶片是否存在响应和适应相应坡位土壤水分和营养特性的长期机制。结果表明:1)土壤水分含量随坡位上升而显著下降,存在明显的梯度,且3个坡位的土壤养分均较为缺乏,但上坡土壤氮和有机质质量分数显著低于中坡和下坡;2)不同坡位生长的2种灌木的叶特性显著不同,随坡位上升,三裂绣线菊和黄刺玫叶片均面积缩小、质量减轻,比叶面积却未改变;3)2种灌木上坡植株叶片的可溶性糖和非结构性碳水化合物质量分数以及全氮质量分数和氮磷比均显著小于下坡植株,仅碳氮比值显著高于下坡植株,但有机碳、全磷和可溶性蛋白质量分数无差异;4)野生三裂绣线菊和黄刺玫成年植株的叶形态和营养特性虽然差异较大,但随坡位变化的响应趋势相同、程度相近且与土壤水分含量变化耦合度更高;5)2种灌木的叶片对于坡位带来的天然土壤水分梯度具有相似的长期响应机制。
三裂绣线菊; 黄刺玫; 坡位; 水分胁迫; 叶特性
在中国北方的干旱、半干旱区域,灌木是石质山体、贫瘠土壤上植物群落的主要先锋植物。灌木群落能够涵养土壤水分,改善土壤营养和减少水土流失,具有重要的生态功能,是目前干旱区造林的主要类型[1]。干旱区域内常常出现且持续的土壤水分胁迫可显著影响植物的生理过程、形态结构和生长生存,因此,旱生植物的生理生态响应和适应机制一直是干旱区研究的重点[2]。过去,关于旱生植物适应干旱的生理生态机制的研究通常都在人工控制的实验室和大田环境中完成,研究的持续时间较短,一般在1个生长季之内,同时水分胁迫处理的强度较大,可导致植株严重受损或死亡[3-5]。在天然旱生环境中,土壤水分条件受降水影响而变异较大,土壤水分的严重匮乏持续时间也较短;所以,植物所经受土壤水分胁迫的程度可能达不到实验处理的强度。同时,野生旱生植物不仅要响应干旱带来的土壤水分胁迫,而且要应对高温和高光等环境因子的胁迫,而在实验条件下,除土壤水分外的其他环境因子都得到了很好的控制;因此,野外生长的天然旱生植物不仅具有适应极端土壤水分条件的短期机制,也有可能在长期生存中因适应多个环境因子的共同胁迫而产生不同的长期适应机制[6]。目前关于野外条件下旱生植物的生理生态响应和适应研究仍很少见到,而了解不同野外土壤水分条件下野生耐旱灌木在生理生态特征及干旱适应机制上的异同,可进一步补充控制实验的研究结果,有利于完善对于旱生植物干旱适应机制的认识。
三裂绣线菊(Spiraeatrilobata)和黄刺玫(Rosaxanthina)是黄土高原地区常见的2种灌木,生长在海拔600~1 500 m的山地上,不仅是贫瘠土壤上旱生灌木群落的优势种,而且在乔木林下也可见到[7]。虽然这2种灌木都具有抗旱能力强、生态适应性广的特点且分布范围相对重叠,但之前的研究发现,黄刺玫的耐阴能力高于三裂绣线菊,表明这2种灌木对环境胁迫具有一定的生理生态响应和适应差异[8]。近年来,对于三裂绣线菊和黄刺玫的研究多集中生物学特性、根系分布和多样性等方面[9-10],相关水分胁迫研究多为人工控制条件下的短期研究[11-12],对于野外成年植株的研究尚未见报道。笔者利用山地环境中坡位造成的自然土壤水分梯度代替水分胁迫处理,以三裂绣线菊和黄刺玫野生成年植株为实验材料,通过调查不同坡位上生长的2种灌木的叶形态和营养特性,研究因坡位不同而产生的土壤含水量差异是否对2种灌木成年植株的叶特性有影响,其影响是否与短期控制研究一致,同时,探讨天然土壤水分梯度下2种灌木是否存在不同的长期响应和适应机制。
1 研究区概况
研究地点选在山西省太原市杏花岭区小返乡窑头村(E112°39′,N37°56′)。研究区域属于黄土高原半干旱丘陵区,具有明显的温带大陆性季风气候特点,年均温在9 ℃左右,极端最高温度40 ℃,极端最低温度-25 ℃,年日照时间2 400~2 600 h,无霜期180~215 d,年降水量450~500 mm,年蒸发量1 700~1 850 mm。研究地海拔1 000~1 250 m,土壤母质为风积黄土,土层厚度10~40 cm,坡度5°~40°,其中上坡坡度20°~40°,中坡5°~15°,下坡0°~5°。植被分布以灌丛为主,主要组成物种包括黄刺玫、三裂绣线菊、虎榛子(Ostryopsisdavidiana)、荆条(Vitexnegundovar.heterophylla)、野豌豆(Viciasepium)、白羊草(Bothriochloaischaemum)等,其中黄刺玫盖度为30%左右,三裂绣线菊盖度为20%左右。经年轮调查,样地中黄刺玫灌丛的年龄为(20±3)年,三裂绣线菊灌丛(10±2)年。研究地点人为经营、放牧和干扰活动少,天然灌木保存良好。
2 研究方法
2.1 样地设置
试验采用裂区设计,在同一南坡的坡面上选择上坡、中坡和下坡3个坡位设置调查样地。3个坡位间的直线距离>200 m,海拔范围为1 150~1 250 m,土壤为黄土土质。在3个选择的坡位上分别选择相距10 m以上的黄刺玫和三裂绣线菊灌丛各5丛。每灌丛内选择5株处于优势高度且长势基本一致的单株植株作为测定样株,2种灌木共选择150株样株。
2.2 参数测定
1)土壤水分含量的测定。2013年6月初到9月初、2014年5月初到8月中旬连续2个生长季内,使用HH2(UK)便携式土壤水分计与20 cmWET土壤水分探头,每间隔1周于正午监测灌丛旁0~20 cm土层的土壤水分含量1次,雨天时推迟1 d测量。
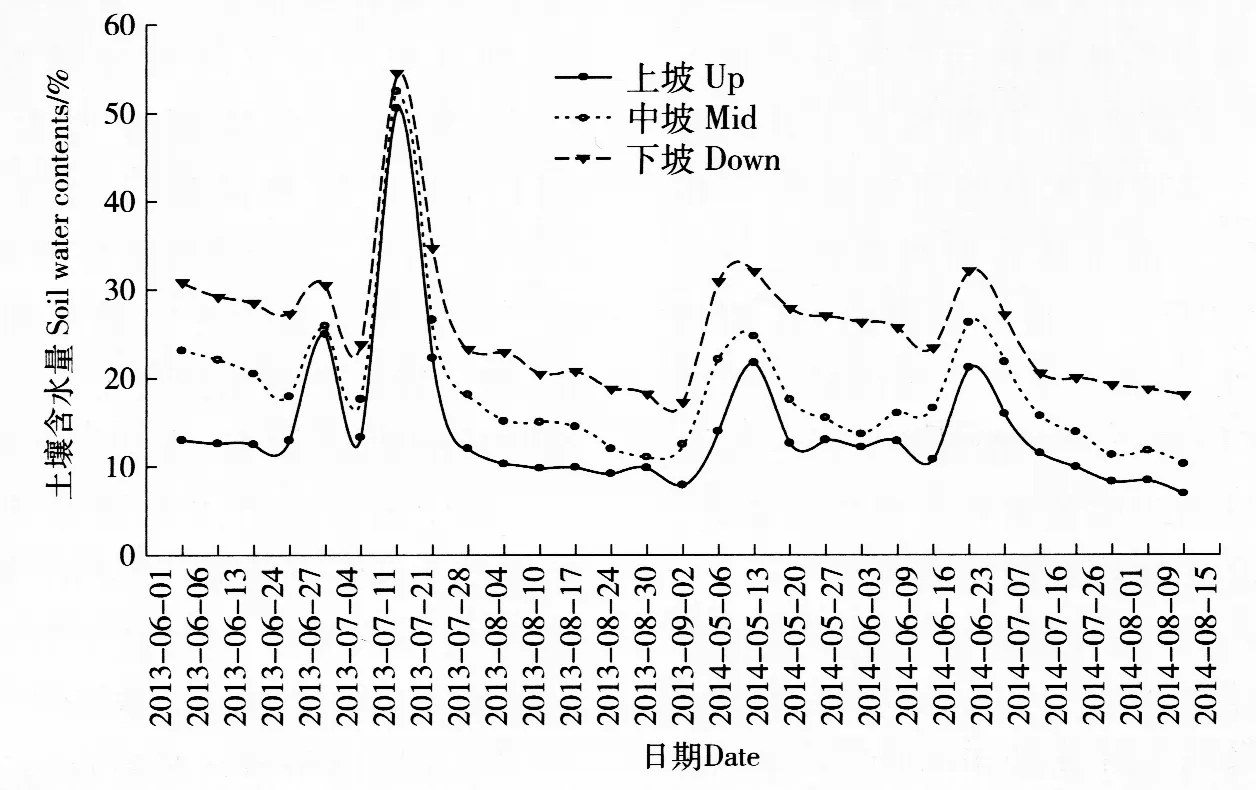
图1 不同坡位0~20 cm土层土壤含水量监测图Fig.1 Soil water contents of 0-20 cm layer at different slope positions
2)土壤养分质量分数的测定。2014年8月末,在待测三裂绣线菊和黄刺玫灌丛旁,用直径5 cm的土钻采集0~20 cm和20~40 cm层土壤带回实验室,风干后,采用电位法测定土壤pH值,K2Cr2O7氧化—外加热法测定土壤有机质质量分数,半微量开氏定氮法测定全氮质量分数以及HCIO4—H2SO4法测定全磷质量分数,速效磷质量分数采用钼锑抗比色法测定,速效钾质量分数采用NH2OAC浸提—火焰光度法测定,速效氮质量分数采用碱解扩散法测定[13]。
3)叶形态指标的测定。采集2种灌木测定样株叶片30~50片带回实验室,首先用高精度游标卡尺测定叶片厚度,然后用扫描仪扫描叶片到计算机中,利用Adobe Photoshop软件测量所有叶片的长、宽和面积,并计算平均单叶叶长、单叶叶宽和单叶叶面积。测量后,在65 ℃下将所有叶片烘干至恒质量,用电子天平测定干质量,计算单叶质量以及比叶面积(单叶面积和单叶质量之比)。
4)叶营养指标的测定。将用于叶形态测量的烘干叶片,加入液氮研磨粉碎,均匀混合为粉末后,分别采用蒽酮比色法测定可溶性糖质量分数,考马斯亮蓝法测定可溶性蛋白质质量分数,重铬酸钾外加热法测定有机碳质量分数,半微量开氏定氮法测定全氮质量分数以及HCIO4- H2SO4法测定全磷质量分数[13]。所有测量均重复3次。叶碳氮比为叶有机碳与全氮质量分数之比,叶氮磷比为叶全氮与全磷质量分数之比。
2.3 数据分析
不同坡位0~20 cm土层的土壤水分含量采用单因素方差分析法分析。因土层及坡位与土层间交互作用不显著(数据未显示),土壤养分质量分数数据也采用单因素方差分析法分析。2种灌木的叶特性数据则以不同坡位上2种灌木每灌丛的平均值作为样本数据,进行坡位和树种双因素方差分析。差异显著(P<0.05)的指标采用LSD检验进行多重比较。数据分析使用SAS/STAT®8.0完成。
3 结果与分析
3.1 不同坡位土壤水分含量分析
连续2个生长季对不同坡位的土壤水分含量进行监测,结果见图1。可以看出,生长季中3个坡位土壤平均绝对含水量分别为上坡(14.13±8.32)%,中坡(18.66±8.13)%,下坡(25.88±7.47)%,变异较大,但3个坡位间的差异均显著(P<0.01)。
在自然环境中,地形因子是直接决定植物生长生存所需的光照、温度以及土壤水分和营养等环境条件的主要因素之一,可间接影响植物物种的分布及植被类型[6,14]。在较小尺度上,地形因子的变化同样能够导致环境条件的改变,如本研究中的坡位。尽管本研究涉及的上、中、下3个坡位位于同一坡面上,不同坡位间土壤平均水分含量的差异仍然显著。除了2013年7月中降水明显偏多的期间外,整个生长季内下坡的土壤含水量均在20%~30%之间(图1),与黄土高原的土壤田间持水量(23.4%)相当[15]。与之相比,上坡和中坡的土壤相对含水量仅为下坡土壤的50%和70%左右,对其上生长的植物构成了一定的水分胁迫,但其程度仍低于控制实验的极端低值[12,15]。同时,大量降水可在短期内导致上坡和中坡土壤水分饱和,降水的效果在最长一周的时间内即可消除(图1);因此,整个坡面上的灌木种群在生长季内虽然经历土壤水分条件的快速、极端变化,但仍是生活在一个天然的土壤水分梯度下。
3.2 不同坡位土壤养分质量分数分析
不同坡位0~40 cm土层的土壤养分质量分数测定结果见表1。可以看出,与土壤水分含量相比,3个坡位0~40 cm土层的pH值、速效磷质量分数、速效钾质量分数和全磷质量分数无显著差异,而上坡土壤中速效氮与全氮质量分数显著低于中坡和下坡,有机质质量分数则随坡位上升而显著减小。可知,天然环境中不同坡位上的土壤含水量和有机质质量分数存在较大的差异。
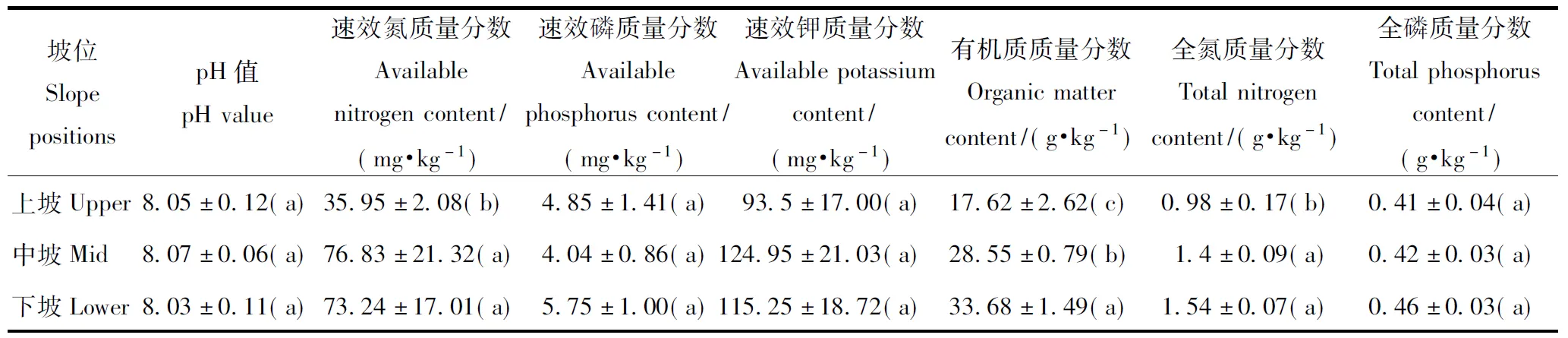
表1 不同坡位0~40 cm土层的土壤养分质量分数
注:不同小写字母表示坡位间差异显著(P<0.05)。Note: Different lowercase letters indicate significant differences among slope positions.
黄土高原地区属于半干旱区,地形破碎,坡度大,土壤侵蚀强烈,在土壤水分含量较低的同时,土壤养分质量分数也较低[15-17]。本研究中,研究坡面上3个坡位的土壤氮、磷、钾质量分数按国家标准处于较为缺乏状态,土壤可供植物利用的养分严重不足。土壤有机质质量分数较高,且随坡位上升显著减少,与其他关于黄土高原灌木林的研究结果[15]相似,可能是由不同坡位上灌木林长期凋落物返还土壤数量不同造成的;然而,3个坡位土壤的pH值均在8.0以上,碱性较强,不利土壤有机质的矿化和植物吸收利用,因此,不同坡位间存在着明显的土壤水分含量差异,同时伴随着土壤氮素和有机质的缺乏。
3.3 不同坡位2种成年植株叶形态特性
对2种野生灌木成年植株的叶形态指标进行分析,结果见图2。可以看出,除比叶面积未受坡位影响外,从下坡到中坡到上坡,其叶片的长、宽、厚、质量及面积均呈显著减小趋势,且不同坡位间三裂绣线菊和黄刺玫叶形态变化幅度不同。随着坡位的上升,三裂绣线菊和黄刺玫单叶的长度和厚度均减小且降幅相近,其中叶长降幅均为46%左右(图2(a)),叶厚降幅均为33%左右(图2(b))。在叶片宽度、质量和面积上,三裂绣线菊的降幅大于黄刺玫,即其单叶叶宽降幅分别为49%和29%(图2(c)),单叶面积降幅分别为72%和60%(图2(d)),单叶质量降幅分别为69%和60%(图2(e))。另外,三裂绣线菊与黄刺玫的叶形态显著不同。三裂绣线菊叶片的长度和厚度相对黄刺玫较小,但二者的差异则随坡位上升而减小;而三裂绣线菊叶片的宽度、质量、面积和比叶面积则始终较大。
以上结果说明,2种灌木成年植株的叶片基本随坡位上升而成比例地减小、减轻,导致比叶面积(图2(f))不改变,且坡位上升对三裂绣线菊叶形态的影响大于对黄刺玫的影响。
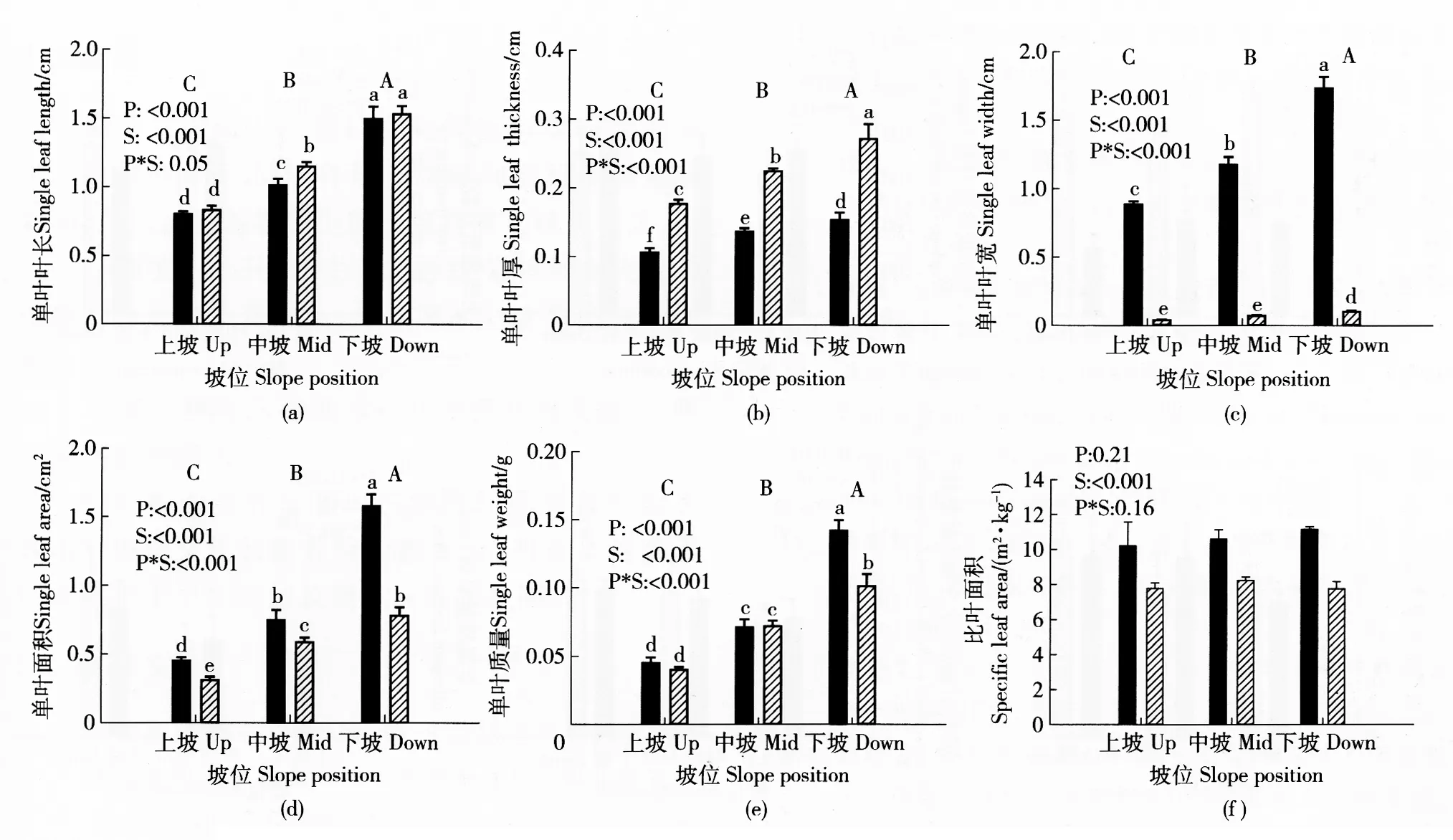
数值为平均值+标准差。不同大写字母表示坡位间的差异显著(P<0.05),不同小写字母表示坡位和树种间交互作用显著(P<0.05),P表示坡位,S表示树种,P*S表示坡位与树种间的交互作用。下同。Note: Data are presented by Means±SE. Different uppercase letters indicate significant differences among slope positions (P<0.05). Different lowercase letters indicate significant interactions between slope position and shrub species (P<0.05). Abbreviations: P, slope position; S, shrub species; P*S, interaction between slope position and shrub species. The same below. 图2 不同坡位三裂绣线菊和黄刺玫的叶形态指标Fig.2 Leaf morphological traits of Spiraea trilobata and Rosa xanthina adult plants grown at different slope positions
干旱生境中生长的植物为了适应有限的土壤水分条件,通常具有较小的叶片以减少水分损失,同时叶肉细胞排列较密、叶片较厚且角质层发达以减轻强光对叶片的损伤[18]。本研究中,随坡位上升,三裂绣线菊和黄刺玫植株叶片缩小、叶质量减轻,与土壤水分变化较为一致,表现出对干旱生境的响应和适应,与短期研究的观察结果相一致;然而,2种灌木的叶片厚度随坡位上升显著减小,比叶面积却未改变,说明这2种灌木在叶形态特征上并不完全具有旱生植物的特点。同时,干旱生境通常伴随着土壤营养物质的贫乏,从而有可能作为混杂因子影响了2种灌木叶形态的变化。此外,三裂绣线菊和黄刺玫具有广泛的生态适应性,如之前的研究就发现,三裂绣线菊和黄刺玫能够在林下分布,具有一定的耐阴性,特别是黄刺玫[11];因此,不同坡位上这2种灌木的叶形态变化是其较强适应能力的表现。
3.4 不同坡位2种成年植株叶营养特性
坡位改变未影响2种灌木成年植株叶的可溶性蛋白、有机碳和全磷质量分数(图3(a)、(b)、(c)),却显著影响了其他叶营养指标。随着坡位的上升,2种灌木叶的可溶性糖和非结构性碳水化合物质量分数显著变小,均减少19%左右(图3(d)、(e))。此外,三裂绣线菊和黄刺玫上坡和中坡植株叶的全氮质量分数显著小于下坡植株且无显著交互作用,降幅分别为27%和42%(图3(f))。与养分质量分数随坡位变化的趋势相反,2种灌木叶片的碳氮比随坡位上升显著增大,其中三裂绣线菊上坡植株高出下坡34%,黄刺玫高出59%(图3(g))。虽然2种灌木上坡和中坡植株叶片的氮磷比值显著低于下坡植株,坡位和树种间的交互作用同样显著(图3(h))。中坡三裂绣线菊植株的氮磷比值显著低于上坡和下坡植株30%左右,黄刺玫则是上坡和中坡植株显著低于下坡植株37%左右(图3(h))。另外,在所有坡位上,三裂绣线菊叶的可溶性蛋白质量分数均低于黄刺玫,碳氮比均高于黄刺玫。
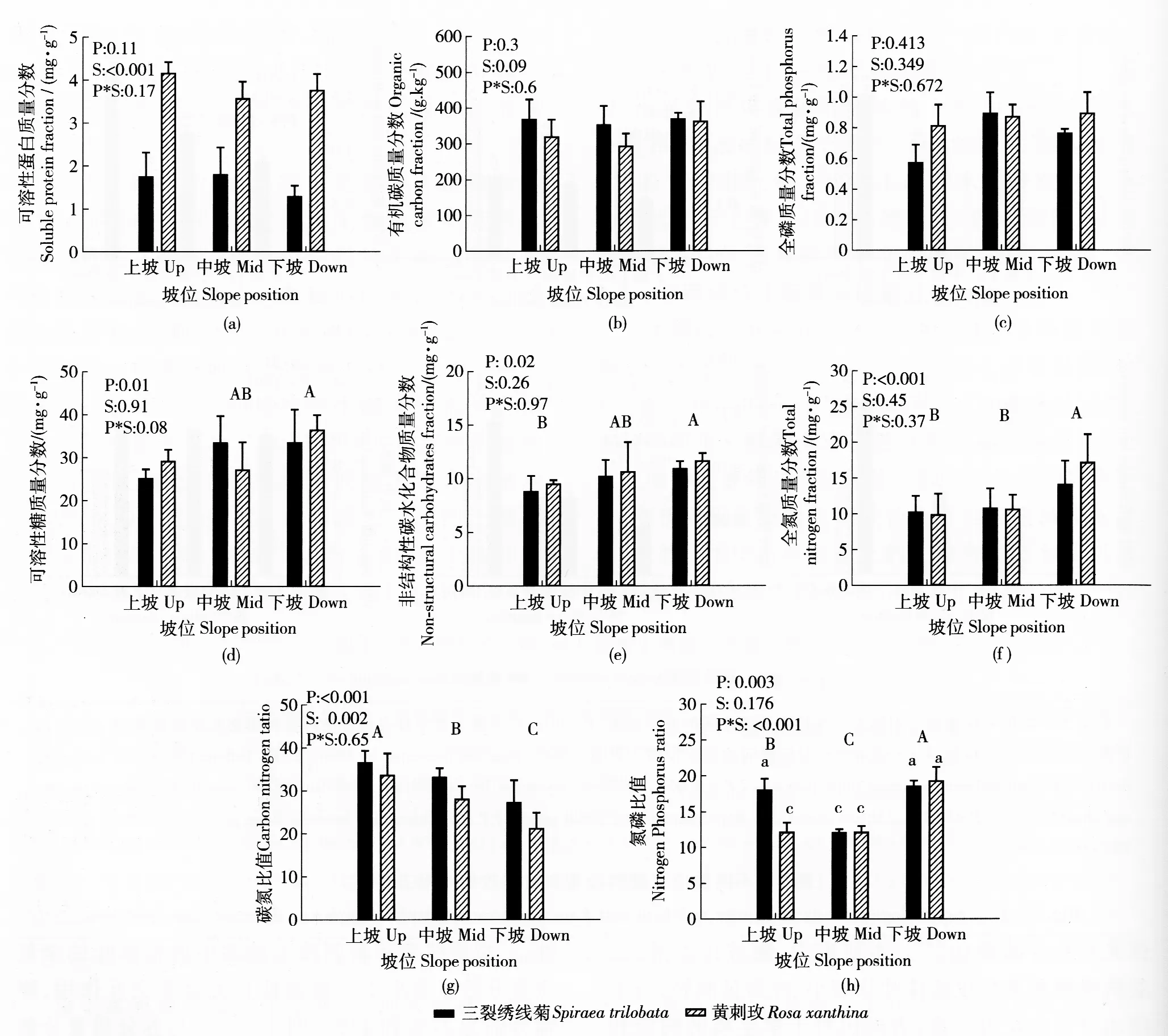
图3 不同坡位三裂绣线菊和黄刺玫的叶营养指标Fig.3 Leaf nutrients of Spiraea trilobata and Rosa xanthina adult plants grown at different slope positions
与下坡植株相比,长期生活在上坡的三裂绣线菊和黄刺玫植株的叶可溶性糖和非结构性碳水化合物质量分数相对较少,可溶性蛋白质量分数无差异,而人工控制的短期水分胁迫常导致可溶性糖和可溶性蛋白质量分数增加[16,19]。可溶性糖和非结构性碳水化合物不仅是叶细胞渗透压的主要调节物质,也是叶光合作用的产物,具有提供细胞能量和营养的作用。叶细胞渗透压的调节有可能是一种短期的响应机制,这种短期机制在长期的土壤水分亏缺下可能无法长时间维持[20]。长期干旱也会导致植物改变能量和营养物质的储存模式,将更多物质输送到根系统中以增强水资源的获取和利用,或储存起来以等待有利的土壤水分条件出现。另外,上坡坡位上土壤中的可利用氮和有机质质量分数较低,有可能导致植株氮营养不足,从而影响碳水化合物的合成,进一步加剧了干旱条件对于植株的不利影响。
干旱不仅会影响植物的生理过程和形态建成,也会改变植物的营养状态。水分胁迫下叶营养元素浓度的增高或保持,有利于叶细胞的渗透调节、生理代谢及增强保水能力[21]。在短期的干旱模拟研究中,土壤含水量下降可造成不同植物叶中的大量元素质量分数增加、保持不变、减少或先增加后减少,反映了植物的耐旱能力存在着较大的种间差异[21-23]。本研究中,三裂绣线菊和黄刺玫叶的碳、氮、磷质量分数与北方荒漠植物相近[23-24],但中上坡植株叶有机碳和全磷质量分数与下坡植株相同,仅全氮质量分数显著较少,同时,随着坡位的上升,2种灌木的碳氮比升高、氮磷比下降。以上结果说明,氮可能是黄土高原干旱生境中这2种灌木的主要限制性营养元素,与干旱减少氮质量分数却增加磷质量分数的“去耦合”趋势相一致[25]。需要注意的是,虽然强碱性的土壤会显著降低土壤有机质的矿化速率,但不同坡位间土壤有机质质量分数的显著差异仍有可能影响植物叶片营养对土壤水分含量变化的响应。
4 结论
1)同一坡面上的坡位变化能够形成明显的天然土壤水分梯度,同时伴随土壤氮和有机质质量分数的差异,虽然各坡位上的土壤营养均较为贫乏。
2)随坡位上升,野生三裂绣线菊和黄刺玫成年植株的叶片均显著变小,与土壤水分含量的变化相耦合。
3)坡位越高,2种灌木叶片的营养物质越少,受氮限制影响越大。
4)叶特性差异显著的三裂绣线菊和黄刺玫表现出对坡位变化的相近响应趋势,说明这2种灌木叶片对于干旱生境的长期适应机制相同。
[1] 张淑勇,周泽福,张光灿,等.水分胁迫下天然次生灌木山桃和山杏光合气体交换特征 [J]. 西北植物学报,2008,28(12): 2492-2499
[2] 王孟本,李洪建.林分立地和林种对土壤水分的影响[J].水土保持学报,2001,15(6): 43-46
[3] 韦莉莉,张小全,侯振宏,等.杉木苗木光合作用及其产物分配对水分胁迫的响应[J].植物生态学报,2005,29(3): 394-402
[4] Dias P C,Araujo W L,Moraes G A B K,et al. Morphological and physiological responses of two coffee progenies to soil water availability [J].Journal of Plant Physiology,2007,164(12): 1639-1647
[5] 蔡海霞,吴福忠,杨万勤.干旱胁迫对高山柳和沙棘幼苗光合生理特征的影响[J].生态学报,2011,31(9): 2430-2436
[6] Valladares F, Pearcy R W. Drought can be more critical in the shade than in the sun: a field study of carbon gain and photo-inhibition in a Californian shrub during a dry El Nio year [J].Plant, Cell & Environment, 2002,25: 749-759
[7] 山西省农业区划委员会.山西树木图志[M].北京:科学出版社,1991:243
[8] 王暾,郭晋平,刘宁,等.森林光环境对4种天然灌木的光合作用和形态的影响[J].林业科学,2011,47(6): 56-63
[9] 王芳,高甲荣,朱继鹏,等.晋西黄土高原三种灌木的根构型研究[J].干旱地区农业研究,2006,24(5): 146-150
[10] 于新晓,张晓明,王雄宾.北京山区天然灌丛植被群落特征与演替规律[J].北京林业大学学报,2008,30(2): 107-111
[11] 吕劲松,申静,李小磊,等.不同土壤水分下黄刺玫光合生理生态特性的初步研究[J].水土保持应用技术,2007(5): 1-3
[12] 佟永兴,崔世茂,陈玉珍,等.水分胁迫对黄刺玫抗旱生理特征的影响[J].内蒙古农业大学学报,2011,32(2): 141-144
[13] 张韫.土壤·水·植物理化分析教程[M].北京:中国林业出版社,2011:222-228
[14] Zhao Tingting, Li Junpeng, Zhang Xinjun, et al. Photosynthetic and morphological characters ofCaraganamicrophyllain different slope aspects and positions [J]. Acta Ecologica Sinica,2011,31(3): 163-168
[15] 薛晓辉,卢芳,张兴昌.陕北黄土高原土壤有机质分布研究[J].西北农林科技大学学报:自然科学版,2005,33(6): 69-74
[16] 姜雪昊,穆立蔷,王晓春,等.3种护坡灌木对干旱胁迫的生理响应[J].草业科学,2013,30(5): 678-686
[17] 张晓霞,李占斌,李鹏,等.黄土高原小流域不同地貌下土壤有机碳的分布特征[J].西安理工大学学报,2010,26(3): 309-313
[18]李芳兰,包维楷,刘俊华,等.岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究[J].应用生态学报,2006,17(1): 5-10
[19] 安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干旱胁迫的生长与生理响应[J].生态学报,2011,31(3): 716-725
[20] 张士功,刘国栋,刘更另.植物营养与作物抗旱性[J].植物学通报,2001,18(1): 64-69
[21] 汪贵斌,曹福亮.盐分和水分胁迫对落羽杉幼苗的生长量及营养元素含量的影响[J].林业科学, 2004, 40(6): 56-62
[22] Pezeshki S R,DeLaune R D,Anderson P H.Effect of flooding on elemental uptake and biomass allocation in seedlings of three bottom land tree species [J].Journal of Plant Nutrition,1999,22(9): 1481-1494
[23] 刘国琴,何嵩涛,樊卫国,等.土壤干旱胁迫对刺梨叶片矿质营养元素含量的影晌[J].果树学报,2003,20(2): 96-98
[24] 张香凝,崔令军,王保平,等.土壤干旱胁迫对Larreatridentata叶片矿质营养元素含量的影响[J].生态环境,2008,17(6): 2387-2390
[25] Bohnerth H J,Jensen R G. Strategies for engineering water-stress tolerance in plants [J].Trends in Biotechnology,1996,14(3): 89-97
(责任编辑:宋如华)
Leaf traits of adult plants of wildSpiraeatrilobataandRosaxanthinagrown at different slope positions
Wang Fang, Liu Ning
(College of Forestry, Shanxi Agricultural University,030801,Taigu,Shanxi,China)
The objective of this study was to explore the responses and acclimations of wildSpiraeatrilobataandRosaxanthinato soil water and nutrient conditions at different slope positions. We selected five wild adult plants for each of the two shrub species grown at three different slope positions (upper slope, mid-slope and lower slope) on a south-facing slope as experiment materials. In growing seasons of 2013 and 2014, we monitored soil water contents at the three slope positions once a week from June to September. Soil nutrient contents were measured in July 2014. In August 2014, we measured leaf morphological traits and nutrient contents of the plants. The results showed that the average soil water contents decreased significantly with ascending slope positions. Although soil pH, contents of total phosphorus and potassium were generally low and similar at all three positions, soil nitrogen and contents of organic matters were significantly lower at upper slope than at mid- and lower slope sites. Meanwhile, leaf traits of the two shrub species grown at different slope positions differed significantly. With slope position rising, both leaf area and mass of the adult plants of the two shrub species decreased, but the specific leaf area kept unchanged. Furthermore, contents of soluble sugar, non-structural carbohydrate and total nitrogen and total phosphorous in plant leaves of both species at upper slope position were significantly lower than those of plants grown at mid- and lower slopes, while the mass ratio of total carbon to total nitrogen was significantly higher at upper slope position. However, contents of organic carbon, total phosphorous and soluble protein in leaves did not change with slope positions for both species. Although there were significant differences in most leaf traits ofS.trilobataandR.xanthina, their responses to changing slope positions were similar, both in direction and in magnitude, well coupled with the soil water gradient created by the slope. The above results suggest a similar long-term response and acclimation of leaves of wildS.trilobataandR.xanthinaplants to soil water gradient in the natural environment.
Spiraeatrilobata;Rosaxanthina; slope position; water stress; leaf traits
2014-08-15
2015-03-06
王芳(1980—),女,硕士研究生。主要研究方向:园林植物的生理生态适应性。E-mail: wangfangljy@163.com
†通信作者简介: 刘宁(1975—),男,硕士,讲师。主要研究方向:植物生理生态学和森林遗传。E-mail: dningliu@yahoo.com
S718.5
A
1672-3007(2015)03-0117-07
项目名称: 教育部高等学校博士学科点专项科研基金“华北山地混交林树种功能特性和功能型研究”(20121403110001);山西农业大学科技创新基金“水分和光照条件在华北山地灌木群落共存机制中的作用(201310)
猜你喜欢
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
现代农村科技(2021年11期)2021-11-05
诗潮(2019年10期)2019-11-19
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
中国水土保持科学(2017年1期)2017-04-18
诗潮(2017年2期)2017-03-16
绿色科技(2017年3期)2017-03-14
绿色科技(2017年1期)2017-03-01
植物营养与肥料学报(2012年3期)2012-10-26
