
海岛棉黄烷酮3—羟化酶(F3H)基因序列分析和表达
2015-04-29 00:44:03刘超张俊姚正培倪志勇陈全家
棉花科学 2015年3期
刘超 张俊 姚正培 倪志勇 陈全家


摘要:根据棉纤维发育相关基因转录组学、表达谱分析结果,从海岛棉(新海21)中筛选到一个与棉纤维发育相关的黄烷酮3-羟化酶基因(GbF3H)。生物信息学分析表明,该基因编码区长1107bp,编码368个氨基酸,多序列比对发现GbF3H序列保守性较高,具有典型的2-酮戊二酸双加氧酶结构域。实时定量PCR分析表明,GbF3H基因在纤维发育的不同时期中均有表达,在开花后Sd的胚珠中表达量最高,推测GbF3H基因可能对棉纤维发育具有重要作用。
关键词:海岛棉;黄烷酮3-羟化酶基因;序列分析;基因表达
中图分类号:S562.01
文献标志码:A
文章编号:2095-3143(2015)03-0007-06
DOI:10.3969/j.issn.2095-3143.2015.03.002
O 引言
类黄酮(flavonoids)是一类广泛分布在植物体内的次生代谢物,是植物器官中色素的主要成分。它保护植物减弱UV-B和外界环境带来的氧化损伤,并且在花粉管萌发、种子休眠和生长素运输中起到重要作用。类黄酮的合成涉及酶的种类繁多,但主要分为3种:酮戊二酸-依赖的双加氧酶类、细胞色素P450羟化酶类和NADPH依赖的还原酶类。其中酮戊二酸一依赖的双加氧酶类黄烷酮3-羟化酶(flavanone 3-hydroxylase,F3H)作为类黄酮合成途径中的核心酶,最早在矮牵牛中被证实,其功能是将黄烷酮羟基化,形成二氢黄酮醇,参与花青素的形成,最终可决定花色。该基因的缺陷性突变体是从金鱼草中获得,突变后可使花色变淡。
棉花是中国重要的经济作物,其体内富含的大量次生代谢物在棉纤维发育及抗逆抗病的过程中发挥着重要作用。早在构建的第一个棉纤维cDNA文库中,就发现参与类黄酮代谢的基因在纤维的伸长期和次生壁合成期有显著上调表达,并且在棕色棉中类黄酮代谢的相关基因也有显著上调。通过抑制类黄酮合成途径的其中一个F3H基因,在花瓣色素生物合成和纤维的发育过程中,发现F3H不仅决定着纤维的质量(如纤维长度和马克隆值),也影响着纤维颜色(棕色纤维)的发育。
为探明黄烷酮3-羟化酶基因在类黄酮合成途径中对棉纤维发育的影响,本研究根据海岛棉纤维具有长、细、强的特点与陆地棉形成明显对比,并针对陆地棉和海岛棉的转录组高通量测序(Solexa)结果,对陆地棉和海岛棉转录组文库进行数据差异比较分析,筛选得到一个海岛棉胚珠棉纤维发育表达明显的基因GbF3H,利用生物信息学方法对GbF3H基因进行分析,并通过实时荧光定量PCR进一步分析其开花后在陆地棉与海岛棉中的表达情况,为进一步阐明黄烷酮3-羟化酶在棉花纤维发育中的作用奠定了基础。
1 材料与方法
1.1 材料
海岛棉(新海21号)和陆地棉(新陆中36号)播种于试验田,棉花盛花期分别取开花当天(0 DPA)的胚珠和开花后5、10、15和25不同天数DPA的纤维组织,液氮速冻,-80℃冰箱保存。
1.2 方法
1.2.1 RNA的提取与cDNA第一链的合成 按TRIzol试剂(TIANGEN)说明,提取棉纤维发育不同时期的组织总RNA。使用DNase I(TIANGEN)去除潜在的DNA污染。cDNA第一链的合成按照M-MLV反转录酶(TaKaRa)说明书完成。
1.2.2 GbF3H基因cDNA序列克隆及分析 根据NCBI登录的海岛棉GbF3H(Gene Bank ID:DQ912945)基因的cDNA序列设计引物(表1),以开花后5d棉纤维cDNA为模板,扩增基因序列。反应程序为94℃预变性5min后,94℃变性30s,58℃退火45s,72℃延伸1min,30个循环后,72℃再延伸10min,4℃保存。经1.0%琼脂糖凝胶电泳检测PCR产物,按照凝胶回收试剂盒(TIANGEN)纯化目的片段,将目的片段连接至PMD19-T(TaKaRa)载体,通过转化E.coli DH5a感受态细胞,筛选阳性克隆送上海生工生物公司测序。利用Prot-Param程序(http://web.expasy.org/protparam/)推测氨基酸的基本性质,CDD程序寻找保守结构域(http://ww-w.ncbi.nlm.nih. gov/cdd/),BlastP检索获得同源序列,Clustal W输出同源比对结果,Espript在线分析蛋白二级结构。
1.2.3 GbF3H基因的DNA序列克隆 按照基因组DNA试剂盒(TlANGEN)说明书提取新海21叶片DNA,用引物(表1)进行PCR扩增、克隆并测序(方法同1.2.2)。用GSDS在线软件(http://gsdsl.cbi.pku.edu.cn/in-dex.php)分析确定GbF3H基因中内含子的插入位点和长度。
1.2.4 实时定量PCR分析 利用实时定量PCR方法,以开花当天(0 DPA)的胚珠和开花后不同天数(5、10、15和25DPA)CDNA为模板,检测GbF3H和GhF3H基因在胚珠和不同发育时期棉纤维中的表达差异。根据基因序列设计定量引物(见表1),使用Life公司的7500 Fast Real PCR System实时荧光定量PCR仪,以SYBRGreen(TaKaRa)为荧光染料,进行实时定量分析。反应程序为95℃预变性3min,95℃变性15s,60℃退火15s,72℃延伸20s,共40个循环。以棉花UBQ7为内参基因,采用2-△△Ct法对数据进行分析,实验采用3次生物学重复。
2 结果
2.1 GbF3H基因ORF序列克隆及分析
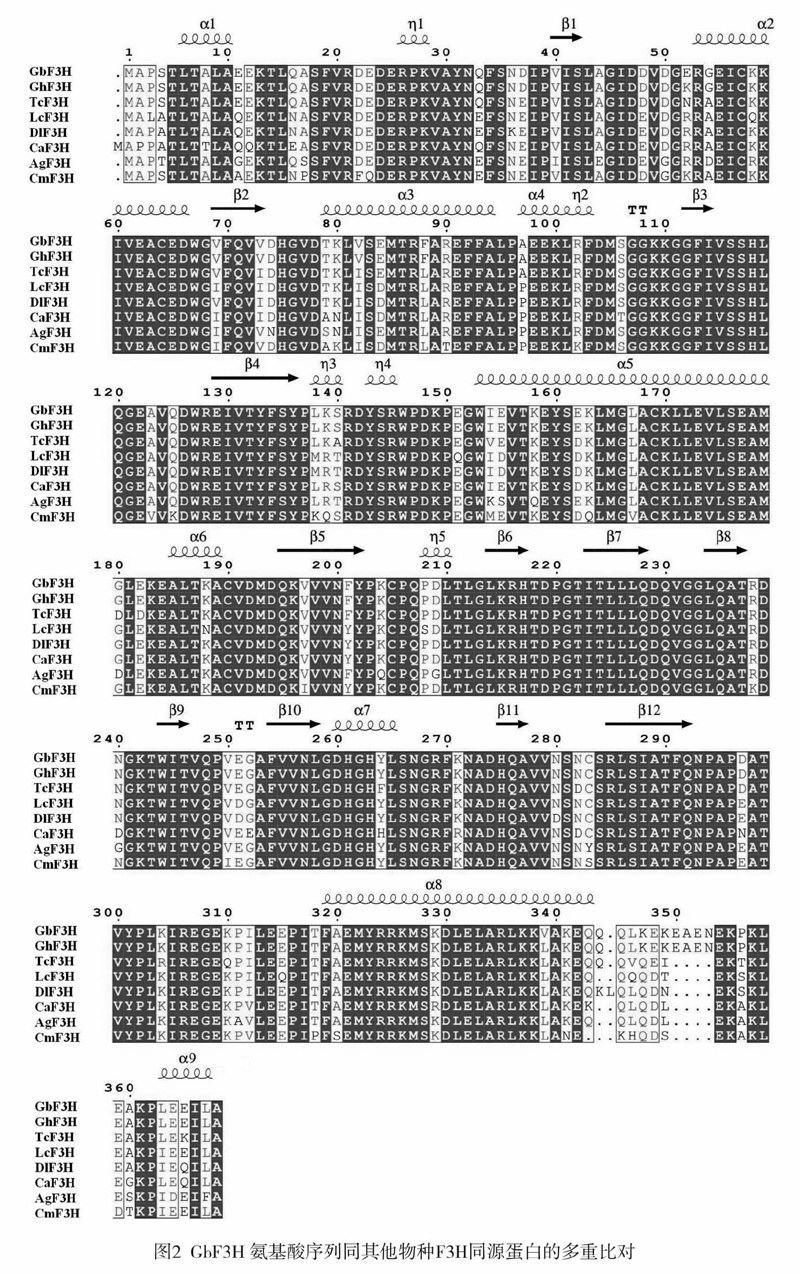
从海岛棉中获得的GbF3H基因序列编码区长1107bp(见图1),编码368个氨基酸,预测分子量约为41.424kDa,等电点为5.28。蛋白二级结构中a螺旋占34.78%,延伸序列占21.2%,自由卷曲占44.02%(见图2)。同源序列比对表明,海岛棉GbF3H与陆地棉GhF3H(ABM64799)的氨基酸序列同源性高达99.46%,仅相差两个氨基酸(52E-K和339V-L),进一步将可可(Theobroma cacao,XP-007046697)、荔枝(Litchichinensis,AD095201)、龙眼(Dimocarpus longan,AB048521)、橄榄(Canarium album,AE036935)、显齿蛇葡萄(Ampelopsin grossedentata,AFN70721)、柚子(Citrus maxima,ADB92595)这些物种的F3H氨基酸序列进行多重比对,发现F3H蛋白高度保守,都具有2一酮戊二酸双加氧酶结构域。
2.2 GbF3H基因DNA序列分析
以新海21基因组DNA为模板,克隆到GbF3H基因组序列,用GSDS在线软件分析确定GbF3H基因全长168lbp,包含3个外显子和2个内含子(见图3)。3个外显子的长度分别为357bp、432bp和318bp(图中淡色标记部分)。第一个内含子的长度为488bp,第二个内含子长度为86bp,分别插入在L119氨基酸之后和H263氨基酸之后。
2.3 基因的表达特性
为了揭示F3H基因在海岛棉和陆地棉胚珠和不同发育时期棉纤维中的表达情况,以棉花UBQ7为内参基因定量分析表明,F3H基因在胚珠和各纤维发育时期中都有表达,且在SDPA的棉纤维中表达量达到最高,说明F3H基因在棉纤维发育起始阶段发挥着一定的功能。随着纤维的发育进入伸长期,到次生壁开始加厚,GbF3H和GhF3H基因的表达都呈现下降趋势,较陆地棉而言,海岛棉GbF3H下降幅度更为明显(图4)。
3 结论与讨论
类黄酮合成途径中的黄烷酮3-羟化酶(F3H)属于依赖2一酮戊二酸的双加氧酶家族,同时需要Fe2+、分子氧、抗坏血酸盐及2-酮戊二酸作为辅助因子参与催化反应。F3H蛋白氨基酸中的H217、H275和D219是Fe2+的结合位点,R285是2-酮戊二酸的结合位点(图2),这些氨基酸残基在酶发挥生物学功能时起重要作用。近几十年的研究工作中,已确定了许多重要的纤维发育相关因素。然而,相关的机制在很大程度上仍然未知。当前以大规模转录分析和其他组学研究为基础,为深入了解、研究相关机制带来了方便。
关于F3H的深入研究,绝大多数集中在其色素生物合成功能方面,发现棉花的F3H参与花色及纤维颜色的形成,并且在棕色棉纤维中的表达水平明显高于白色棉纤维中。本文针对海岛棉和陆地棉在纤维品质上差异,通过挖掘公开数据和转录组数据得到一个与类黄酮生物合成代谢相关的黄烷酮3-羟化酶基因。通过定量分析表明,黄烷酮3-羟化酶参与纤维发育,特别是纤维发育的初期,F3H基因的表达明显上调,并且GbF3H的表达量高于GhF3H;而在纤维发育的中后期,F3H基因在海岛棉中的表达水平低于陆地棉,可能在纤维的品质方面起到负调控的作用。由图4可知,棉花F3H基因在2个棉种的纤维发育的次生壁加厚期(10-25DPA)表达逐渐下降,在海岛棉中尤为明显。主要的表达差异在纤维发育的初期(5DPA),这与徐州142和其突变体的纤维发育比较中,优先发现类黄酮的基因在胚珠纤维细胞中表达25结果相同,这可能说明黄烷酮3-羟化酶是纤维细胞发育中介导该通路的关键因素。
总而言之,次生代谢物黄烷酮3-羟化酶可能参与棉纤维的发育,具体的影响机制有待进一步研究。今后如若采用有效的方法控制类黄酮合成途径中相关基因的表达,可为改善纤维品质提供新的途径。
猜你喜欢
江苏农业科学(2017年22期)2018-01-06 00:58:58
江苏农业科学(2017年1期)2017-02-27 10:30:11
中国科技博览(2016年24期)2016-12-28 23:53:07
中国实用医药(2016年30期)2016-12-28 16:54:00
山东农业科学(2016年11期)2016-12-17 21:00:42
中国民族民间医药·上半月(2016年10期)2016-11-19 11:08:04
中国科技博览(2016年22期)2016-11-01 13:58:50
江苏农业科学(2016年5期)2016-07-23 18:29:00
中国种业(2016年3期)2016-04-14 09:48:07
江苏农业科学(2015年8期)2015-09-10 21:03:17
