
东北豆科山羊豆族4属植物叶形态特征及数量分类学研究
2015-04-15 08:04:20史传奇刘玫王臣
草业学报 2015年5期
史传奇,刘玫,王臣
(哈尔滨师范大学生命科学与技术学院,黑龙江省分子细胞遗传与遗传育种重点实验室,黑龙江 哈尔滨 150025)
东北豆科山羊豆族4属植物叶形态特征及数量分类学研究
史传奇,刘玫*,王臣
(哈尔滨师范大学生命科学与技术学院,黑龙江省分子细胞遗传与遗传育种重点实验室,黑龙江 哈尔滨 150025)
本研究采用GMA切片、扫描电镜及叶片透明法,深入研究了东北豆科山羊豆族4属25种植物叶的形态结构。选取与叶相关的23个形态性状,参考8个非叶的特征,进行了聚类分析与主成分分析。结果表明,相似系数在0.58处时,4属植物被分为3组,黄耆属与米口袋属为一组,棘豆属和甘草属各自为一组。小叶是否具腺体、叶柄横切面形状及叶柄细胞是否具单宁等特征具有较高的绝对权重值,可以作为区分刺果甘草与甘草和其他山羊豆族植物的依据。托叶质地、托叶与叶柄联合程度及小叶叶缘是否反卷等特征可作为棘豆属的识别特征。托叶形状在属下种间具一定的分类学价值。本研究为山羊豆族植物分类学及豆科植物系统学研究提供了叶形态学依据。
山羊豆族;叶;形态;结构;分类学
叶的形态结构(如气孔类型、表皮细胞垂周壁式样、叶柄横切面形状等)对分类学具有重要的价值[1-8]。Mehrabian等[9]认为伊朗黄耆属(Astragalus)Incani组叶柄维管束数目、髓部薄壁细胞层数等特征可以作为种的分类依据,但对组间分类的意义不大。康云和李明理[10]指出蔟毛黄耆亚属(subgenusPogonophace)叶表皮细胞形状与垂周壁式样可作为属下组间的分类依据,角质层纹饰对种的鉴定具有一定的意义。甘草属(Glycyrrhiza)植物叶表面气孔有不等型和无规则型,可作为研究甘草属植物分类和演化的依据[11]。陆嘉惠等[12]研究甘草属叶表皮特征,提出叶表皮细胞垂周壁式样,气孔类型及毛状体类型可作为划分甘草属内种以上等级的依据。刘丽和赵一之[13]根据托叶与叶柄分离或合生、小叶着生方式及小叶数目等形态特征,利用分支分类的方法,探讨了内蒙古棘豆属(Oxytropis)27个物种的系统发育关系。
形态学及分子系统学研究揭示山羊豆族(tribe Galegeae)是一个复系类群,分类学一直存在诸多争议[14],本研究的目的是通过深入研究东北山羊豆族4属植物叶的形态结构,选取23个质量性状,并结合前人研究工作,通过聚类分析与主成分分析,为该族植物的分类及系统学研究提供形态学依据。
1 材料与方法
1.1 材料
研究材料为山羊豆族黄耆属13种,甘草属2种,米口袋属2种及棘豆属8种。新鲜材料于2014年5-9月采自于中国东北部,经哈尔滨师范大学王臣教授鉴定。凭证标本存放于哈尔滨师范大学植物标本馆(Harbin Normal University, HANU),干材料于2014年9月取自哈尔滨师范大学植物标本馆和东北生物标本馆(Institute of Forestry and Pedology, IFP)。详细信息见表1。
1.2 方法
叶片透明:按照任海燕等[15]的方法,每一物种取两个植株的成熟叶片,将其置于5%NaOH水溶液中,24 h后取出,50%甘油水溶液封片,Olympus BX53显微镜观察叶表皮细胞特征,重复观察3次,用Olympus DP26数码相机照相。参照Dilcher[16]系统鉴定气孔类型。
叶片扫描:根据陈曦等[17]的方法,切取正方形(4 mm2)成熟叶片,用双面胶将样品粘在样品台上,Eiko IB-3离子溅射仪导电处理,溅射镀膜厚度为7~8 nm,在Hitachi S-4800型扫描电镜下(加速电压为5 kV)观察毛状体形态并照相。
GMA(glycol methacrylate,乙二醇甲基丙烯酸酯)切片[18]:新鲜材料用FAA(Formalin-Acetic acid-70% Alcohol mixture,福尔马林∶冰醋酸∶70%酒精=5∶5∶90)固定,时间不少于24 h。干材料放入热水中约30 min,材料充分吸水膨胀后,用FAA固定24 h以上。每个物种各取3份叶柄的中部,经50%酒精、100%酒精、异丙醇、正丁醇脱水处理各2次,每次6 h。进入GMA渗透3次,前2次各为1 d,第3次5~7 d,之后将材料及GMA装入胶囊,置于60℃温箱聚合24 h。用Leica Ultralcut R切片,厚度2.0~2.5 μm。将切片置于DNPH(2, 4-dinitrophenylhydrazine,2, 4-二硝基苯肼)冰醋酸水溶液中30 min,流水冲洗10 min。置于高碘酸水溶液中10 min,流水冲洗5 min,将切片烘干后,置于Schiff试剂30 min,流水冲洗1~2 min,切片烘干。最后置于甲苯胺蓝溶液中染色1~2 min,流水冲洗10 min,烘干后用加拿大树胶封片。切片同样经Olympus BX53显微镜观察,Olympus DP26数码相机拍照。
根据《中国植物志》[19-20]记载的8个非叶形态特征及本研究所观察的23个叶的形态特征,参照徐克学[21]的方法,对31个质量性状编码:01生长习性:多年生草本0/一或二年生草本1;02花序:叶丛基部叶腋处着生0/总状1/伞形2;03花萼形状:管状钟形0/钟形1/管状2/筒状3;04花药药室顶部:不联合0/联合1;05花柱柱头蔟毛:无0/有1;06龙骨瓣具喙:否0/是1;07果皮质地:革质0/膜质1;08果皮附属物:无0/有1;09叶着生方式:基生或近基生0/茎生1;10叶型:轮生羽状复叶0/奇数羽状复叶1;11托叶质地:草质0/膜质1;12托叶形状:近卵形0/三角形1/披针形2;13托叶先端形态:渐尖0/锐尖1;14托叶与叶柄联合情况:离生或仅基部联合0/中部附近联合1/大部分联合2;15小叶基部:近圆形0/圆形或楔形1/楔形2;16小叶先端:多钝0/钝或尖1/多尖2;17小叶叶缘:全缘0/微齿1;18小叶叶缘反卷:否0/是1;19小叶多细胞腺体:无0/有1;20小叶表皮毛状体分布:无0/下表面1/两面2;21小叶边缘毛状体:无0/有1;22小叶非腺毛类型:单毛0/丁状毛1;23小叶气孔分布:下表面0/两面1;24小叶气孔类型:只具无规则型0/兼具不等型及无规则型1;25小叶上表皮垂周壁式样:弓形0/弓形-波形1/波形2;26小叶下表皮垂周壁式样:弓形0/弓形-波形1/波形2;27叶柄横切面形状:近圆形0/心形1/戟形2;28叶柄近轴面凹槽:无0/有1;29叶柄维管束排列:不连续0/近连续1;30叶柄横切面皮层细胞形状:近圆形0/柱形1;31叶柄细胞内含单宁:否0/是1。
参考Zhou等[22]的方法,应用NTSYS pc 2.10e软件包,采用UPGMA法(unweighted pair group method using arithmetic average,非加权配对算术平均法)构建聚类树,Mantel’s test[23]检验聚类树与相似系数矩阵的一致性。参照李鸿雁等[24]的方法,应用SPSS 17.0做主成分分析,求出各主成分的特征值、贡献率、累计贡献率及前3个主成分中各性状特征的权重值,并基于前2个主成分绘制二维散点图。

表1 东北山羊豆族4属植物(用于叶特征研究)基本信息[19-20]
2 结果与分析
2.1 叶片形态及叶柄结构
叶外部形态:山羊豆族叶着生方式分为基生或近基生和茎生,黄耆属多为茎生,如斜茎黄耆,而乳白黄耆和新巴黄耆的叶为基生或近基生。叶型多为奇数羽状复叶,也有轮生羽状复叶,如棘豆属的海拉尔棘豆。托叶草质或膜质,棘豆属多为膜质,多枝棘豆的为草质,其他3属多为草质,而乳白黄耆为膜质。托叶形状划分为近卵形、三角形和披针形,在种间有差异,如米口袋属的狭叶米口袋的为三角形,少花米口袋的为披针形。托叶先端渐尖或锐尖,黄耆属的达乌里黄耆、小米黄耆和小果黄耆的为锐尖,其余物种的皆为渐尖。托叶和叶柄分离或仅在基部联合,大部分物种属于此类,黄耆属的糙叶黄耆、棘豆属的硬毛棘豆等联合至托叶中部,棘豆属的山泡泡和砂珍棘豆的几乎全部联合。小叶基部近圆形或楔形,也有同一植株上不同叶片的小叶基部兼具圆形和楔形的,如狭叶米口袋。甘草属和棘豆属小叶先端多尖,米口袋属的小叶先端形态多样,钝或尖,黄耆属的多为钝圆,或有时具凹陷,如黄耆。除甘草属的刺果甘草叶缘具微齿外,其余物种叶缘均为全缘。棘豆属小叶叶缘反卷(硬毛棘豆除外),为本属特有。

图1 山羊豆族植物叶表皮毛状体、垂周壁及气孔形态Fig.1 Trichome, anticlinal wall and stomata morphology of leaf epidermis of tribe GalegeaeA: 斜茎黄耆叶上表皮 Upper epidermis of A. adsurgens leaf; B1: 甘草叶上表皮腺体 Gland on the upper epidermis of G. uralensis leaf; B2: 甘草叶上表皮Upper epidermis of G. uralensis leaf; C1: 少花米口袋叶下表皮单毛 Single-celled hair on lower epidermis of G. verna leaf; C2: 少花米口袋叶下表皮 Lower epidermis of G. verna leaf; D: 砂珍棘豆叶上表皮 Upper epidermis of O. racemosa leaf. 标尺 Scale bar: 100 μm in A; 50 μm in B1, C2, D; 25 μm in B2; 200 μm in C1.

图2 山羊豆族植物叶柄横切面结构Fig.2 Petiole structure in transversal section of tribe GalegeaeA: 斜茎黄耆 A. adsurgens; B: 草木樨状黄耆A. melilotoides; C: 刺果甘草G. pallidiflora; D: 少花米口袋G. verna; E: 砂珍棘豆O. racemosa; F: 多叶棘豆O. myriophylla; G: 多枝棘豆O. ramosissima. c: 柱状皮层细胞 cylindrical cortex cell; e: 表皮 epidermis; h: 毛状体 hair; t: 单宁 tannin; v: 维管束 vascular bundle. 标尺 Scale bar: 300 μm in A, C; 200 μm in B, G; 500 μm in D; 450 μm in E, F.
叶表皮特征:叶表面具多细胞腺体、丁状毛和单毛。刺果甘草和甘草小叶上、下表面均具有多细胞的腺体(图1B1)。黄耆属的部分种具丁状毛,如斜茎黄耆(图1A)等,多数物种具单毛,如少花米口袋(图1C1)。毛状体通常分布于小叶上、下表面及叶缘。小叶上、下表面均具有气孔分布,仅黄耆属的黄耆小叶的上表面未见气孔。气孔主要为无规则型和不等型,多数物种兼具两种类型(图1A,C2,D),刺果甘草和甘草(图1B2)的气孔仅具无规则型。叶表皮垂周壁呈弓形、兼具弓形与波形和波形,如少花米口袋(图1C2)下表皮细胞垂周壁为弓形,斜茎黄耆(图1A)的为弓形-波形,甘草(图1B2)的为波形。
叶柄结构:叶柄横切面为心形、近圆形或戟形,黄耆属和米口袋属的皆为心形,如斜茎黄耆(图2A)、草木樨状黄耆(图2B)和少花米口袋(图2D),刺果甘草的为戟形(图2C),棘豆属具心形或近圆形,前者如砂珍棘豆(图2E),后者如多枝棘豆(图2G)。多数物种叶柄近轴面具凹槽(图2A~E)。叶柄具毛状体,表皮细胞1层,皮层为4~6层薄壁细胞,呈柱状(其细胞长轴同叶柄横切面平行)或圆形,前者多见于草木樨状黄耆(图2B)和多枝棘豆(图2G)。黄耆属、甘草属和米口袋属叶柄具3个明显较大的维管束,分布不连续,近连续分布的只出现在棘豆属部分物种,如多叶棘豆(图2F)。单宁仅存在于刺果甘草和甘草叶柄细胞内(图2C),其余3属均未见。详细特征见表2。

表2 山羊豆族植物31个形态学质量性状编码
2.2 聚类分析
聚类树与相似系数矩阵的一致性较好(r=0.82,t=11.57)。聚类结果如图3,在SM(simple matching,简单匹配)系数为0.56时,棘豆属最先被分出,成为独立的一个类群。在0.58时,甘草属被分出,而黄耆属和米口袋属仍聚在一起。至0.83处,米口袋属才从黄耆属区分出来。聚类结果可以明显看出,本研究选取的31个形态学性状能较好地区分黄耆属、甘草属及棘豆属,但狭叶米口袋与少花米口袋和黄耆属的物种区分不十分明显,并且对属下个别种的区分效果也不十分显著,如黄耆属的华黄耆和丹麦黄耆、棘豆属的海拉尔棘豆和砂珍棘豆,说明所选取的31个质量性状在两者之间有较高的相似性。
2.3 主成分分析
主成分分析结果(表3)可以看出,前3个主成分的累计贡献率为51.36%,至第6个主成分累计贡献率达到70%以上。从31个质量性状中筛选出与叶相关的形态特征,第1主成分中小叶是否具腺体(0.973)、小叶气孔类型(-0.973)及叶柄细胞是否含单宁(0.973)具有相同的绝对权重值,说明对4属植物具有同等的分类价值,小叶叶缘形态的权重值为0.788,这些性状都可作为区分刺果甘草与甘草和其他3属植物的重要依据。第2主成分中托叶质地(0.818)、托叶与叶柄联合程度(0.716)及小叶叶缘是否反卷(0.755)的绝对权重值大于0.7,可以作为棘豆属植物的识别特征。第3主成分中叶柄形状(0.728)、近轴面是否具凹槽(0.759)及维管束分布(0.675)具有较高的绝对权重值,可作为属间及棘豆属属下种间的分类依据。

图3 基于山羊豆族植物31个质量性状的聚类分析树状图(UPGMA)Fig.3 Cluster analysis dendrogram (UPGMA) based on 31 qualitative characters of tribe Galegeae
基于第1主成分和第2主成分绘制二维散点图(图4),与聚类分析结果(图3)十分相近,4属植物能较好地被分为3个组。黄耆属13个物种和米口袋属2个物种分布比较紧密,区分不明显。第1主成分坐标轴上甘草属2个物种和第2主成分坐标轴上棘豆属8个物种的分布比较分散,说明对此2属属下种间具有较高的分类学价值。
3 讨论与结论
本研究选取的23个叶形态结构的质量性状,对多份材料进行分析和归纳,部分叶外观形态特征与前人研究结果[19-20]基本相似。基于31个质量性状进行的聚类分析和主成分分析结果,对研究山羊豆族4个属的属间及种间关系进行了探讨。
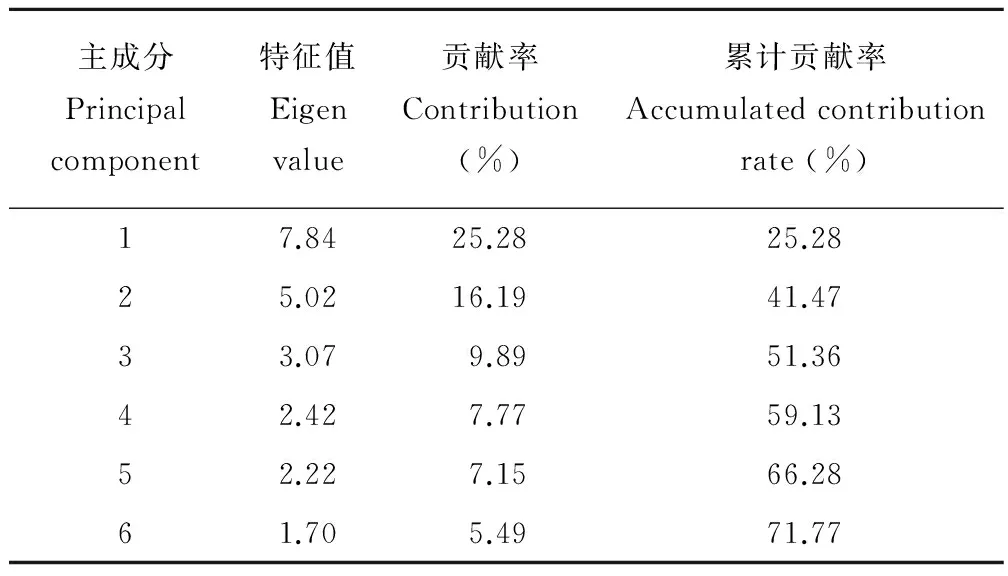
表3 基于山羊豆族植物31个质量性状的主成分分析

图4 基于山羊豆族植物31个质量性状的前2个主成分构建的二维散点图Fig.4 2-dimensional plot based on the first two principal components of 31 qualitative characters of tribe Galegeae
3.1 黄耆属
傅坤俊等[19]因小叶具丁状毛等特征将新巴黄耆、糙叶黄耆及乳白黄耆等划入裂萼亚属(subgenusCercidothrix),本研究聚类分析结果中,前两者分在一起,关系较近,但他们与乳白黄耆较远。黄耆属是维管植物第一大属,形态学与分子系统学研究都认为它是一个多态性类群,现有的分类系统对该属的属下类群的划分尚有争议[4,14,26-27],而叶的形态特征是否可以作为黄耆属内的分类依据也有诸多争论,如Mehrabian等[9]认为黄耆属Incani组的叶解剖特征与其他组的差异不大,不宜作为组间的分类依据。Haddad和Barnett[25]提出叶柄薄壁细胞和厚角组织细胞数目可以作为黄耆属一些自然类群的划分依据。康云和张明理[10]认为叶表皮特征可作为蔟毛黄耆亚属内组的分类依据。本研究统计分析结果与现有的黄耆属内亚属及组的划分有一些差异,作者认为叶的形态结构可为黄耆属植物区分种及探讨种间亲缘关系提供参考。
3.2 甘草属
本研究揭示刺果甘草和甘草只具无规则型气孔,可作为区分刺果甘草与甘草和其他物种的依据。但陆嘉惠等[12]提出甘草也具有不等型气孔,且冯元忠和闫平[11]发现在甘草属的黄甘草(G.enrycarpa)等物种中也具有不等型气孔,可见无规则型气孔只适于探讨属下种间关系,不宜作为区分甘草属与山羊豆族其他属的依据。Lewis等[14]通过对甘草属形态学与分子系统学的研究,认为甘草属不应归入山羊豆族。孟雷[28]提出甘草属与山羊豆族其他属在系统树上处于不同的分支,同样认为将其放在山羊豆族中不太合适。本研究聚类分析表明SM系数为0.58时,甘草属与其他3属分开。主成分分析中第1主成分可知,小叶具腺体、叶柄形状为戟形及叶柄细胞含单宁等特征将刺果甘草与甘草和其他3属分开,距离较大,这在一定程度上反映甘草属植物的确与其他3属有差别。
3.3 米口袋属
聚类分析可知,米口袋属与黄耆属关系密切。狭叶米口袋和少花米口袋具伞形花序[29-30],与山羊豆族其他物种不同,而其他形态特征和黄耆属的新巴黄耆与糙叶黄耆十分相似,因此叶形态结构不能区分米口袋属与黄耆属。主成分分析表明少花米口袋与狭叶米口袋不易区分,前者叶为长椭圆形或披针形,而后者叶在夏秋季时为线形[20,29-30]。本研究还发现两者的托叶形状、小叶毛状体分布也有所不同,可以作为区分两物种的依据。
3.4 棘豆属
棘豆属与黄耆属关系很近[4,14,20]。高静等[31]和卢萍等[32-33]通过内蒙古棘豆属植物分子系统学研究,认为棘豆属与黄耆属各成一支,前者应为单系类群。本研究聚类分析结果表明棘豆属最先被分出,单独成为一个组。主成分分析结果可以看出,第1主成分坐标轴上棘豆属、黄耆属及米口袋属的关系很近,而第2主成分坐标轴上看,棘豆属与后两者之间又存在一些差异,如托叶膜质、小叶叶缘反卷都可作为识别棘豆属的重要特征,故支持棘豆属为单系类群。硬毛棘豆、多枝棘豆等叶柄近轴面无凹槽、横切面形状及维管束排列成圆形,这些特征相对原始[34],在一定程度上说明棘豆属较其他3属原始。
本研究揭示了东北豆科山羊豆族不同类群叶的形态结构,对探讨属间及种间的分类及亲缘关系具有重要价值,为形态学与分子系统学揭示的山羊豆族植物不是一个自然类群提供了叶形态结构的依据,并为进一步探讨山羊豆族系统学提供了形态学资料。
[1] Hare C L. The anatomy of the petiole and its taxonomic value. Proceedings of the Linnean Society of London, 1943, 155: 223-229.
[2] Metcalfe C R, Chalk L. Anatomy of the Dicotyledons (Vol.1)[M]. Oxford: Clarendon Press, 1950.
[3] Howard R A. The Vascular Structure of the Petiole as a Taxonomic Character[C]. Proceedings of the 15th International Horticultural Congress, Nice, 1958. Pergamon Press, 1962: 7-13.
[4] Lu Y M. Study on Taxonomy and Characters of Leaf Epidermis ofAstragalusL. andOxytropisDC. in Altay Region of Xinjiang[D]. Urumqi: Xinjiang University, 2010.
[5] Sun X Q, Liu M, Sun T H,etal. Morphological stduy of the leaf structures ofViolain northeastern China and discussions of their taxonomic values (Violaceae). Acta Prataculturae Sinica, 2014, 23(2): 223-234.
[6] Cheng X Y, Liu M, Zhang X X,etal. Vegetative organ structures of Ranunculaceae in northeastern China and notes on systematic implications. Acta Prataculturae Sinica, 2014, 23(3): 62-74.
[7] Sun T H, Liu M, Sun X Q,etal. Morphology study on the leaf structures ofPotentillain Northeastern China and its taxonomic value (Rosaceae). Acta Prataculturae Sinica, 2014, 23(3): 75-84.
[8] Shi C Q, Liu M, Wang C,etal. Taxonomic values for leaf structure in the tribe Vicieae (Leguminosae) in northeastern China. Acta Prataculturae Sinica, 2014, 23(6): 157-166.
[9] Mehrabian A R, Zarre S H, Azizian D,etal. Petiole anatomy inAstragalusSect. Incani DC. (Fabaceae) in Iran (a phylogenetical approach). Iran Journal of Botany, 2007, 13(2): 138-145.
[10] Kang Y, Zhang M L. Leaf epidermal feature ofAstragalusL. SubgenusPogonophaceBunge (Leguminosae) from China and its systematic significance. Bulletin of Botanical Research, 2009, 29(1): 25-32.
[11] Feng Y Z, Yan P. Observation on the leaf epidermis stomatal patterns ofGlycyrrhiza. Journal of Shihezi Agricultural College, 1995, 29(1): 1-4.
[12] Lu J H, Li X Y, Zhou L L,etal. Characters of leaf epidermis and their systematic significance inGlycyrrhiza. Acta Botanica Yunnanica, 2005, 27(5): 525-533.
[13] Liu L, Zhao Y Z. A preliminary study on the species ofOxytropisin NeiMonggol by cladistic taxonomic method. Acta Scientiarum Naturalium Universitatis NeiMonggol, 1996, 27(1): 72-82.
[14] Lewis G M, Schrire B, Mackinder B,etal. Legumes of the World[M]. London: Kew Press, 2005.
[15] Ren H Y, Pang Y L, He X J,etal. Characters of leaf epidermis and systematic research inPleurospermumfrom China. Acta Botanica Boreali-Occidentalia Sinica, 2009, 29(1): 49-60.
[16] Dilcher D L. Approaches to the identification of angiosperm leaf remains. The Botanical Review, 1974, 40(1): 154-157.
[17] Chen X, Xing Y, Wang M,etal. Morphology structure of leaf epidermis of genusRubusin Heilongjiang. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(10): 2035-2039.
[18] Feder N, O’ Brien T P. Plant microtechnique: some principles and new methods. American Journal of Botany, 1968, 55(1): 123-142.
[19] Fu K J, He Y Q, He S B,etal. Flora Tomus 42(1)[M]. Beijing: Science Press, 1993.
[20] Cui H B, Zhang Z W, Xu L R,etal. Flora Tomus 42(2)[M]. Beijing: Science Press, 1998.
[21] Xu K X. Numerical Taxonomy[M]. Beijing: Science Press, 1994.
[22] Zhou H, Liao J, Xia Y P,etal. Morphological characteristics for classifying evergreenAzalea(Ericaceae) cultivars in China using numerical taxonomy. Pakistan Journal of Botany, 2013, 45(2): 593-598.
[23] Mantel N A. The detection of disease clustering and a generalized regression approach. Cancer Research, 1967, 27: 209-220.
[24] Li H Y, Li Z Y, Shi W G,etal. A study on leaf anatomic traits and drought resistance ofMedicagorutenicain Inner Mongolia. Acta Prataculturae Sinica, 2012, 21(3): 138-146.
[25] Haddad R S, Barnett J R. Variation in petiole anatomy of the European spiny species ofAstragalusL. (Leguminosae-Papilionoideae-Galegeae). Botanical Journal of the Linnean Society, 1989, 101: 241-247.
[26] Ma J J. The Taxonomical Study ofAstragalusL. in Inner Mongolia[D]. Hohhot: Inner Mongolia Normal University, 2004.
[27] Wojciechowski M F.Astragalus(Fabaceae): A molecular phylogenetic perspective. Brittonia, 2005, 57(4): 382-396.
[28] Meng L. Systematics ofGlycyrrhizaL. (Fabaceae) - With a Special Reference to Its Relationship toGlycyrrhizopisisBoiss. & Bal.[D]. Beijing: Institute of Botany, the Chinese Academy of Sciences, 2005.
[29] Institute of Forestry and Pedology. Flora Plantarum Herbacearum Chinae Boreali-orientalis Tomus 5[M]. Beijing: Science Press, 1976.
[30] Fu P Y. Clavis Plantarum Chinae Boreali-orientalis Editio Secunda[M]. Beijing: Science Press, 1995.
[31] Gao J, Lu P, Wang J N,etal. Molecular phylogeny of several species ofOxytropisDC. based on 5. 8SrDNA/ITS sequence. Acta Agriculturae Boreali-sinica, 2009, 24(6): 168-173.
[32] Lu P, Gao J, Wang J N,etal. Molecular phylogenetic analysis of severalOxytropisDC. species in Inner mongolia based on 5.8S rDNA/ITS sequences. Acta Botanica Boreali-Occidentalia Sinica, 2011, 31(12): 2420-2428.
[33] Lu P, Gao L X, Jin F,etal. Molecular phylogeny of ten species ofOxytropisbased on psbA-trnH sequences. Plant Diversity and Resources, 2014, 36(3): 279-284.
[34] Rodriguez R L. Systematic anatomical studies onMyrrhidendronand other woody umbellates. University of California Publications in Botany, 1957, 29: 145-318.
参考文献:
[4] 逯永满. 新疆阿勒泰地区黄耆属和棘豆属分类及叶表皮特征研究[D]. 乌鲁木齐: 新疆大学, 2010.
[5] 孙雪芹, 刘玫, 孙天航, 等. 东北堇菜属植物叶形态结构的研究及其分类学价值的探讨. 草业学报, 2014, 23(2): 223-234.
[6] 程薪宇, 刘玫, 张欣欣, 等. 东北毛茛科植物营养器官结构及其系统学意义. 草业学报, 2014, 23(3): 62-74.
[7] 孙天航, 刘玫, 孙雪芹, 等. 东北委陵菜属植物叶形态结构的研究及其分类学价值的探讨. 草业学报, 2014, 23(3): 75-84.
[8] 史传奇, 刘玫, 王臣, 等. 东北野豌豆族植物叶形态结构的研究及其分类学意义. 草业学报, 2014, 23(6): 157-166.
[10] 康云, 张明理. 中国黄耆属蔟毛黄耆亚属的叶表皮特征及其系统学意义. 植物研究, 2009, 29(1): 25-32.
[11] 冯元忠, 闫平. 甘草属植物叶表面气孔类型的观察. 石河子农学院学报, 1995, 29(1): 1-4.
[12] 陆嘉惠, 李学禹, 周玲玲, 等. 甘草属植物叶表皮特征及其系统学意义. 云南植物研究, 2005, 27(5): 525-533.
[13] 刘丽, 赵一之. 内蒙古棘豆属植物分支分类的初步探讨. 内蒙古大学学报(自然科学版), 1996, 27(1): 72-82.
[15] 任海燕, 逄云莉, 何兴金, 等. 中国棱子芹属植物叶表皮微形态特征及其系统学研究. 西北植物学报, 2009, 29(1): 49-60.
[17] 陈曦, 邢怡, 王蒙, 等. 黑龙江悬钩子属植物叶表皮形态结构的研究. 西北植物学报, 2012, 32(10): 2035-2039.
[19] 傅坤俊, 何业祺, 何善宝, 等. 中国植物志第42(1)卷[M]. 北京: 科学出版社, 1993.
[20] 崔鸿宾, 张振万, 徐朗然, 等. 中国植物志第42(2)卷[M]. 北京: 科学出版社, 1998.
[21] 徐克学. 数量分类学[M]. 北京: 科学出版社, 1994.
[24] 李鸿雁, 李志勇, 师文贵, 等. 内蒙古扁蓿豆叶片解剖性状与抗旱性的研究. 草业学报, 2012, 21(3): 138-146.
[26] 马建军. 内蒙古黄芪属(AstragalusL.)植物的分类学研究[D]. 呼和浩特: 内蒙古师范大学, 2004.
[28] 孟雷. 甘草属(GlycyrrhizaL.)的系统学研究-兼论与假甘草属(GlycyrrhizopsisBoiss. & Bal.)的关系[D]. 北京: 中国科学院植物研究所, 2005.
[29] 辽宁省林业土壤研究所. 东北草本植物志第五卷[M]. 北京: 科学出版社, 1976.
[30] 傅沛云. 东北植物检索表(第二版)[M]. 北京: 科学出版社, 1995.
[31] 高静, 卢萍, 王金妞, 等. 基于5. 8SrDNA/ITS序列的几种内蒙古棘豆属植物分子系统学研究. 华北农学报, 2009, 24(6): 168-173.
[32] 卢萍, 高静, 王金妞, 等. 基于5. 8SrDNA/ITS序列的内蒙古棘豆属植物分子系统学分析. 西北植物学报, 2011, 31(12): 2420-2428.
[33] 卢萍, 高利霞, 金凤, 等. 基于psbA-trnH序列的十种棘豆属植物分子系统学研究. 植物分类与资源学报, 2014, 36(3): 279-284.
Leaf morphology and numerical taxonomy of 4 genera of the tribe Galegeae in northeastern China
SHI Chuan-Qi, LIU Mei*, WANG Chen
KeyLaboratoryofMolecularCytogeneticsandGeneticBreedingofHeilongjiangProvince,CollegeofLifeScienceandTechnology,HarbinNormalUniversity,Harbin150025,China
The leaf morphological structure of 25 species,representing 4 genera from the tribe Galegeae, northeastern China, was determined using GMA, SEM and leaf transparency methods. Twenty-three morphological features related to leaf and 8 non-leaf characters were selected for cluster analysis and principal component analysis. Three groups were identified;AstragalusandGueldenstaedtiawere placed in one group, andGlycyrrhizaandOxytropisin separate groups. Features including the presence of glands, petiole shape and tannin content of petiole cells were able to distinguishGlycyrrhizapallidifloraandG.uralensisfrom other Galegeae species. Stipule texture, connectivity between stipule and petiole and leaflet margin features are all useful identification characteristics ofOxytropis. Stipule shape is valuable for interspecific classification. This study provided the leaf morphological evidences for taxonomy of the Galegeae and Leguminosae systematics.
Galegeae; leaf; morphology; structure; taxonomy
10.11686/cyxb20150522
http://cyxb.lzu.edu.cn
2014-11-25;改回日期:2015-01-07
哈尔滨师范大学博士研究生创新科研项目(HSDBSCX2013-06)和国家自然科学基金(31270235及31070169)资助。
史传奇(1986-),男,黑龙江齐齐哈尔人,在读博士。E-mail:chuanqi0730@163.com *通讯作者Corresponding author. E-mail:hsd_liumei@163.com
史传奇,刘玫,王臣. 东北豆科山羊豆族4属植物叶形态特征及数量分类学研究. 草业学报, 2015, 24(5): 182-189.
Shi C Q, Liu M, Wang C. Leaf morphology and numerical taxonomy of 4 genera of the tribe Galegeae in northeastern China. Acta Prataculturae Sinica, 2015, 24(5): 182-189.
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31 06:38:50
小天使·四年级语数英综合(2022年6期)2022-06-21 12:49:36
High Technology Letters(2021年4期)2022-01-09 02:08:16
建材发展导向(2021年14期)2021-08-23 00:56:24
作文小学中年级(2021年4期)2021-05-07 01:17:50
河北果树(2020年4期)2020-11-26 06:05:00
祝您健康·文摘版(2019年4期)2019-06-11 10:20:34
海峡姐妹(2019年1期)2019-03-23 02:42:40
中成药(2018年9期)2018-10-09 07:18:46
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:54
