
侧金盏花的花部特征与繁育系统观察
2015-04-08 06:50王阿香陈士惠马翠青
草业科学 2015年3期
孙 颖,王阿香,陈士惠,马翠青,何 淼
(东北林业大学园林学院,黑龙江 哈尔滨150040)
侧金盏花属(Adonis)是毛茛科一年生或多年生草本植物,是一个广布欧亚温带的小属,有26 ~30种。侧金盏花属植物大多属于早春短命或类短命植物,大部分种类都有鲜艳的金黄色花朵,小部分种类具有赤红色花朵,是极具开发潜力的野生草本种质资源。我国共有侧金盏花属植物10 种,1 变种和2变型。其中东北地区主要分布有两种:侧金盏花(A. amurensis)、辽吉侧金盏(A. ramosa)[1]。侧金盏花花色明亮艳丽、花形美观可爱,株型紧凑矮小,符合人们的审美要求。侧金盏花植株开花时间很早,可以填补2 -4 月的园林空白,而且极度抗寒,这意味着高纬度地区也可以露地种植观赏。目前,我国的侧金盏花基本处于野生状态[2],引种驯化后可应用于草坪、花坛、护坡及林间空地,从而减少养护成本,形成色彩丰富、层次多样的草坪景观;而且其夏眠的独特生理特性,不会影响后续的景观设计。因此,侧金盏花作为高寒地区的植物引种栽培用于城市绿化,有着很好的应用前景。
繁育系统是指直接影响后代遗传组成的所有有性生殖特征的总和。包括花部各器官的寿命、花部的综合特征、传粉者的种类和行为特征、花的开放式样、自交亲和程度以及交配系统等[3-4]。繁育系统是植物的遗传机制和环境相互作用的表现,对植物的表征变异和进化有重要影响[5],是繁育生物学和生物多样性保护管理的重要组成部分[6],是认识植物生活史的前提,也是开展生物学相关研究的必备基础[4,7]。花部综合特征及交配系统的定量、定性测定是繁育系统研究中相对独立的内容,其方法和实践已经有较多的相关报道[8-11],而侧金盏属植物的相关研究则几无开展。因此,认识其开花过程的自然规律,掌握繁育系统类型,可以更有效地进行种子生产和良种选育,提高其制种效率,从而为侧金盏花在北方高寒地区的合理应用提供技术支持。
1 材料与方法
1.1 试验材料
试材来源为哈尔滨市帽儿山老山人工林试验站(45°20' N,127°34' E)。试验地平均海拔300 m,年平均气温2.8 ℃,相对湿度70%,降水量723.8 mm,土壤类型为棕褐色森林土。选择有代表性的落叶松林样地,取10 个2 m×2 m 样方,样方内植物密度为6.93 株·m-2。以样方内正常生长的侧金盏花作为研究对象。于2011 年及2012 年4 -6 月进行各项试验。
1.2 试验方法
1.2.1 花部综合特征及开花动态观察 于花蕾期随机标记30 株侧金盏花,每天观察1 次直至单花开放。开花当天及开花第2 天07:00 -17:00 不间断观察,之后每天观测一次,直到花部萎蔫。记录花部各部分发生变化的时间、特点、过程,对所标记的花朵进行拍照[12-13]。盛花期,随机选取正在开放的单花30 朵,测量花朵的直径以及雄蕊、雌蕊各个部分的长度。采集新鲜花药置于FAA 固定液中,经过一系列脱水、包埋,采用苏木精染色法[14]染色,对花药横切,切片厚度为8 mm,在显微镜下进行观察、拍照。
1.2.2 繁育系统检测 采用Cruden[15]方法测定单花花粉量和胚珠数,估算花粉胚珠比P/O(Pollen-ovule Ratio)。根据Dafni[16]的标准对侧金盏花的直径、开花行为进行测定,计算杂交指数(Outcrossing Index,OCI)值。
1.2.3 座果率的测定 对侧金盏花进行5 种套袋处理:1)不去雄,不套袋,自由授粉,作为对照,自然状态下检测传粉情况。2)不去雄,套袋,自花授粉,检测传粉是否需要媒介。3)去雄,人工异花授粉,套袋,检测杂交是否亲和。4)去雄、人工异花授粉,不套袋。5)去雄,套袋,不授粉,检测花朵无融合的生殖率。每个处理取50 朵花,两组重复,统计不同处理下的座果率。
2 结果与分析
2.1 侧金盏花花部综合特征及单花开花动态
侧金盏花是多年生草本植物,根状茎短而粗,有多数须根。株高5 ~12 cm,先花后叶。花单生于主茎或少数分枝的顶端,雄蕊多数,心皮多数。在花朵开放7 ~9 d 后长出叶子,叶片为三角状卵形,二至三回细裂。萼片7 ~17 枚,常为淡灰紫色,长圆形或卵状长圆形,长16.51 ~18.93 mm,花瓣12 ~18 枚,黄色,倒卵形或长倒卵形,长14.91 ~19.09 mm(表1)。花冠直径为18. 82 ~45.64 mm。花柱高1.00 ~2.06 mm,柱头成熟时分泌粘液。花丝长2.12 ~4.84 mm,花药长1.68~2.60 mm,初为黄绿色,随后逐渐加深变为黄色,散粉后花药变为橙黄色。侧金盏花单花约84 个花药,花药4 室,整个花药的横切面呈“X”型。花药壁为四层结构,由内到外分别为绒毡层、中层、药室内壁及表皮(图1),花粉大小约为31.57 μm×22.51 μm。侧金盏花子房长1.16 ~1.96 mm,颜色初为淡绿色,逐渐加深变为深绿色。
连续两年对侧金盏花的植株观察发现,侧金盏花花期在3 -4 月,花蕾的颜色从紫褐色变浅直至花萼开始松动持续6 ~7 d。天气晴好时,侧金盏花通常08:00 左右开放,16:30 左右开始闭合。单花开放时,外侧花萼几乎同时松动,也有1 ~2 个花萼先伸展开,内侧的花瓣也随之松动。30 ~60 min内,内部花瓣全部松动。从花萼松动到所有花瓣伸展到最大冠幅,需要2 ~5 h。花朵初开时花瓣为黄绿色,逐渐变为鲜黄色。开花2 ~3 d,外轮花药开始开裂,其余交替开裂。晴天时花药全部开裂所需时间约为1 h,而阴天时花药散粉时间长1 ~2 d,雨天或雪天花朵则一直处于闭合状态。开花5 d 花萼开始皱缩,花药逐渐萎缩。开花6 ~7 d 花瓣开始萎缩,部分花瓣脱落,花萼包围着花瓣呈螺旋状合拢。开花8 d 之后花瓣、花药全部脱落,仅存发育中的子房(图2)。

表1 侧金盏花花部器官数量特征统计(n=30)Table 1 Floral parameters of Adonis amurensis(n=30)
2.2 侧金盏花的单花持续时间
2011 及2012 年,侧金盏花的单花持续时间分别为8.29 和8.26 d,两者差异不显著(P >0.05),表明侧金盏花在两年内的单花寿命比较稳定。单花持续时间频率最高为8 d,分别占每年统计100 朵花的41%和44%,之后是9、7 和10 d 的单花持续时间,持续时间为6 d 的单花所占频率不到3%(图3)。

图1 侧金盏花花部构造及花药横切面结构Fig.1 Floral structure and cross section view of anther of Adonis amurensis

图2 侧金盏花开花动态过程Fig.2 Flowering dynamic of Adonis amurensis
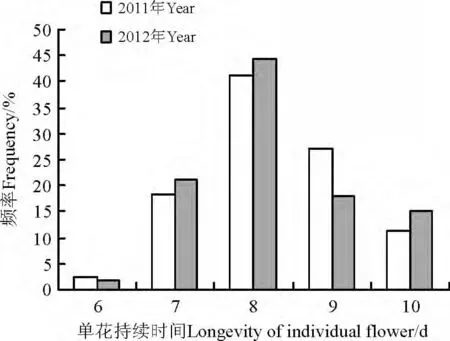
图3 侧金盏花单花持续时间频率分布Fig.3 The frequencies distribution of individual flower longevity of Adonis amurensis
对侧金盏花单花持续时间、花冠直径、花柱高度、花药数量、子房数量的相关分析结果表明(表2),侧金盏花的单花持续时间与花柱高度呈显著正相关(P <0.005),与花冠直径、花药数量、子房数量则无显著相关性(P >0.01)。同时,花柱高度与花冠直径呈显著负相关(P <0.01)。由此可知,花柱高度对单花持续时间有较大影响,花柱越高,单花开放时间越长。
2.3 侧金盏花繁育系统的确定
2.3.1 套袋试验 通过检测不同条件下侧金盏花的座果率发现,去雄、套袋、不授粉,其座果率为0,表明侧金盏花不存在无融合生殖现象。不去雄、不套袋、自由传粉,座果率为78%,不去雄、套袋、自花授粉条件下座果率为58%,去雄、不套袋、自由传粉条件下坐果率为44%,而人工自花授粉、人工异花授粉后授粉率均超过80%。套袋试验结果表明,侧金盏自花可孕,同时需要传粉者进行传粉(表3)。

表2 单花持续时间、花冠直径、花柱高度、花药数量、子房数量的相关分析Table 2 Correlation analysis among flower longevity,diameter of flower,length of style,anther number and ovary number

表3 侧金盏花不同授粉方式的套袋试验结果Table 3 The test results of different pollinating ways of Adonis amurensis
2.3.2 杂交指数(OCI) 按照Dafni 的方法测算,侧金盏花花朵直径约为32.97 mm,大于6 mm,OCI 记为3;侧金盏花的花药在开花第2 天才开始散粉,而柱头在开花前1 天就具有微弱的可授性[17],即为雌蕊先熟,OCI 记为0;花药未成熟时低于柱头,但成熟后二者逐渐向对方靠近,有可能接触进行授粉,OCI 记为0;综合以上结果,侧金盏花的OCI值为3 +0 +0 =3。根据Dafni[16]的繁育系统划分标准:OCI=0,闭花受精;OCI =1,专性自交,OCI=2,兼性自交;OCI=3,自交亲和,有时需要传粉者;OCI=4,部分自交亲和,异交,需要传粉者;侧金盏花的繁育系统为自交亲和,需要传粉者。此结果与套袋试验得出的结论一致。
2.3.3 花粉胚珠比(P/O) 侧金盏花的单花花粉数平均为157 352 粒,单花胚珠数为69 个,P/O 值为2 280。根据Cruden[15]对繁育系统的划分标准:P/O值为2. 7 ~5. 4,闭花授精;P/O 值为18. 1 ~39.0,专性自交;P/O 值为31.9 ~396.0,兼性自交;P/O 值为244. 7 ~2 588. 0,兼性异交;P/O 值为2 108.0 ~195 525.0,专性异交。同杂交指数测算结果及套袋试验结果相结合,综合判定侧金盏花的繁育系统为兼性异交,自交亲和,需要传粉者。
3 讨论
3.1 侧金盏花的花部特征与繁育适应性
侧金盏花属于早春类短命植物,其花期在3 -4月,为在低温气候条件下达到生殖成功的目的,其发展出一系列吸引传粉者的策略:侧金盏花的花径能达到32.97 mm,按照Abe[18]的划分标准属于大型花;花朵为鲜艳的金黄色,黄色对于蜂类是可见花色[19],能比雪地吸收更多热量,达到吸引昆虫的效果;同时,侧金盏花产生大量的花粉和花蜜,是昆虫重要的食物来源,从而增加了植株授粉结实的机会。
Primack[20]、Schoen 和Ashman[21]分析了35 个科110 种植物的单花持续时间,认为系统发生限制了单花持续时间的进化,单花持续时间的变异不是出现在属、种的水平,而是出现在科的水平。侧金盏花的单花花期一般为6 ~10 d,按照Abe[18]的分类标准,其单花花期属于很长类型,较长的花期对于保证花粉流的稳定、增加昆虫的访花行为具有积极意义。从个体水平分析,侧金盏花的单花花期除与系统发生有关以外,还与花柱高度、单花始花时间[2]有关。花柱越高,始花时间越早的侧金盏花,单花持续时间越长。花型、花色、花粉、花期等因子的综合作用,使得侧金盏花在环境条件不利的早春也能达到很高的结实率。
3.2 侧金盏花的繁育系统判断
生殖成功是植物适应环境条件的重要组成部分。关于自交和异交的互相演化是系统发育检验的重点[5]。Dafni 根据花部结构和开花行为进行繁育系统的判断。具有相同OCI 值的植物,一般具有相似的形态和行为特征,繁育系统类型也趋于一致。侧金盏花的OCI 值为3,按照Dafni[16]的标准,侧金盏花的繁育系统为自交亲和,需要传粉者。Cruden[15]认为P/O 值是表示性比的有效指标,其能够简洁、快速地预测繁育系统的基本类型,P/O 值越低的植物,传粉效率越高。按照Cruden[15]的评判标准,侧金盏花的繁育系统为兼性异交,自交可孕。Cruden[15]同时认为,在进行繁育系统的判断时,P/O值的计算应在杂交指数之后。
人工授粉及套袋试验发现,人工自花授粉和人工异花授粉后侧金盏花的座果率均超过80%,说明其自交亲和,且亲和性很高,这验证了上述侧金盏花的繁育系统为混合交配系统的判断。自然条件下,植物的自交率是多种原因作用的结果,不但取决于自身的花粉量、而且取决于从其他个体接受外来花粉的时机、数量以及自花花粉和异花花粉在获得受精机会上的竞争能力[22-24]。侧金盏花在不去雄、套袋、自然自花授粉情况下,座果率仅为58%,说明侧金盏花存在一定的自花限制,花粉数量不足和缺乏花粉与柱头的有效触碰是造成这一现象的主要原因。同时,前期研究结果[17]表明,侧金盏花在开花当天部分柱头即具备一定的可授性,但花药在开花后2 ~3 d 才开裂,造成了自花散粉与柱头可授期在时间上具有一定间隔,为异花授粉提供了可能性。去雄后自由传粉条件下,侧金盏花座果率为44%,说明传粉媒介的效率是阻碍自然异花授粉效果的一个原因。在自然条件下侧金盏花的结实率能够达到78%,应该是异交与自交共同作用的结果。相关研究认为,在极端环境下,植物的交配系统有从异交向自交演变的倾向,从而更好地适应恶劣的生存条件[25-26],对于侧金盏花自交率与自然群落之间的相互关系应进行进一步的深入研究。
4 结论
通过对野生环境下侧金盏花的花部形态特征、开花动态、授粉特性的观察研究,侧金盏花花期在3 -4 月,花型大,单花花期持续6 ~10 d,花部具有吸引传粉者的特征。侧金盏花雌蕊先熟,花药与柱头无空间间隔,杂交指数结果为3,花粉胚珠比为2 280。综合各项指标结果认为,侧金盏花的繁育系统为兼性异交,自交亲和,需要传粉者。
[1] 万清林,刘鸣远.侧金盏花成株年生长节律生理特性的研究[J].植物研究,1996,16(3):351-355.
[2] 何淼,陈士惠,马翠青,孙颖.野生及引种侧金盏花的开花物候与传粉特性[J].草业科学,2014,31(3):431-437.
[3] 陈香,胡雪华,肖宜安,谢骞,王春香,李蕴,戴文红.紫茉莉的花部综合特征与繁育系统[J].生态学杂志,2008,27(10):1653-1658.
[4] 陈雄伟,邵玲,梁广坚,梁霞.紫背天葵花部特征与繁育系统的研究[J].园艺学报,2013,40(2):363-372.
[5] 胡春,刘左军,伍国强,赵志刚.钝裂银莲花花部综合特征及其繁育系统[J].草地学报,2013,21(4):783-788.
[6] 刘芬,李全健,王彩霞,连静静,田敏.濒危植物扇脉杓兰的花部特征与繁育系统[J].林业科学,2013,49(1):53-60.
[7] 赵天荣,蔡建岗,施永泰,倪建刚,沈岚.大花萱草‘金娃娃’的花器结构和繁育系统观察[J].草业科学,2013,30(1):52-57.
[8] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
[9] 方炎明.森林植被的显花、传粉与繁育系统[J].南京林业大学学报(自然科学版),2012,36(6):1-7.
[10] 刘宗才,焦铸锦,董旭升,代金星.鸢尾的花部结构及繁育系统特征[J].园艺学报,2011,38(7):1333-1340.
[11] 林辰壹,韩文娟,祁晨霞,叶强.新疆高葶韭雄蕊开花动态特征与花粉育性研究[J].草业科学,2012,29(4):561-566.
[12] 段友爱,李庆军.少花柊叶传粉生物学的研究[J].植物分类学报,2008,46(4):545-553.
[13] 关文灵,李叶芳,陈贤,杨德.蝴蝶花花器结构和开花授粉生物学特性[J].园艺学报,2009,36(10):1485-1490.
[14] 张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社,2005:127-128.
[15] Cruden R W.Pollen-ovule ratios:A conservative indicator of breeding system in flowering plants[J]. Evolution,1977,31:32-46.
[16] Dafni A.Pollination Ecology:A practical approach[M].New York:Oxford University Press,1992:1-57.
[17] 孙颖,王阿香,陈士惠,马翠青,何淼.野生侧金盏花的花粉活力与柱头可授性[J].草业科学,2014,31(6):1045-1051.
[18] Abe T.Threatened pollination systems in native flora of the Ogasawara(Bonin)Islands[J].Annuals of Botany,2006,98(2):317-334.
[19] 施翔,刘会良,张道远,王建成,杨姗霖,董金鑫.沙丘稀有种准噶尔无叶豆花部综合特征与传粉适应性[J].生态学报,2013,33(18):5516-5522.
[20] Primack R B.Longevity of individual flowers[J].Annual Review of Ecology and Systematics,1985,16:15-37.
[21] Schoen D J,Ashman T L.The evolution of flower longevity:Resource allocation to maintenance versus construction of repeated parts in modular organisms[J].Evolution,1995,49:131-139.
[22] Holsinger K E.Pollination biology and the evolution of mating systems in flowering plants[J].Evolutionary Biology,1996,29:107-149.
[23] Buide M L,Díaz-Peromingo J A,Guitián J. Flowering phonology and female reproductive success in Silene acutifolia Link ex Rohrb[J].Plant Ecology,2002,163(1):93-103.
[24] Brys R,Jacquemyn H,Edels P,Rossum F V,Hermy M,Triest L.Reduced reproductive success in small populations of the selfincompatible Primula vulgaris[J].Journal of Ecology,2004,92:5-14.
[25] 翟新莹,马登坦,王成章,严学兵.海拔对高山植物交配系统的影响[J].草业科学,2012,29(4):640-645.
[26] 张丙林,穆春生,王颖,王彦静.五脉山黧豆开花动态及有性繁育系统的研究[J].草业学报,2006,15(2):68-73.
猜你喜欢
中国科学探险(2022年8期)2022-12-31
作文与考试·小学低年级版(2020年19期)2020-11-16
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
蔬菜(2019年5期)2019-05-21
大科技·百科新说(2018年3期)2018-04-11
文苑·感悟(2016年5期)2016-05-16
浙江农业科学(2013年6期)2013-11-08
中国蔬菜(2013年8期)2013-01-28
植物营养与肥料学报(2011年3期)2011-10-24
中学生数理化·七年级数学华师大版(2008年11期)2008-12-23
