
二型花柱植物红蓼的繁殖生物学研究
2015-03-27 06:53:16文慧慧陈明林张自生
草业学报 2015年11期
文慧慧,陈明林,张自生
(安徽师范大学生命科学学院,安徽省重要生物资源保护与利用研究重点实验室,安徽 芜湖241000)
异型花柱(heterostyly)是由基因调控的花的多态现象,包括二型花柱(distyly)和三型花柱(tristyly),柱头与花药的位置互补,高度不同,即交互式雌雄异位(reciprocal herkogamy)[1]。这种交互式雌雄异位的功能使得传粉者的身体将花的花药授粉与同一水平高度的柱头上,这种功能和进化的意义有利于花粉准确传递,促进异交,避免雌雄器官的性别干扰以及减少自交传粉对植物的不利影响[2]。二型花柱植物常常具有自交不亲和系统,但在不同的植物中又表现出不同程度的可育性;三型花柱常常具有严格的自交不亲和系统。异型花柱的研究是生态学最活跃的领域之一,异型花柱植物是进化生物学的理想材料。Ganders[3]于1979年对异型花柱做了较为详尽的描述,除此之外大都是一些专属专种的研究[4-5]。在我国异型花柱植物广泛存在,该类型植物在特有科属中相继被报道[6]。蓼属(Polygonum)有300多种植物,分布较为广泛,其二型花柱现象最早在荞麦属(Fagopyrum)的荞麦(Fagopyrum esculentum)中被发现[7],蓼属的愉悦蓼(Polygonum jucundum),蚕茧草(P.japonicum)的二型花柱现象已被详细报道[8-10]。
红蓼(Polygonum orientale)为一年生草本。茎直立,粗壮,高1~2m;叶宽卵形或卵状披针形;托叶鞘筒状,被长柔毛,沿顶端具绿色的翅;总状花序顶生或腋生;瘦果近圆形,双凹,黑褐色,有光泽,包于宿存花被内。花期6-9月,果期8-10月。目前对红蓼的研究是对其化学成分、质量控制、药理价值以及园林应用等方面,尚没有对红蓼的二型花柱现象报道。本实验主要通过对红蓼的花部特征和繁育系统的初步探究,以确定其是否为二型花柱植物,是否存在雌雄异位现象,观察其受精方式,并探讨其是否具有异型自交不亲和系统。
1 材料与方法
1.1 材料
实验时间为2013-2014年。实验材料来源于芜湖市天门山、芜湖市鸠江区以及芜湖市镜湖区3个居群,于8-9月间采集的新鲜样品,标本均存放于安徽师范大学标本馆。
1.2 方法
1.2.1 花部生物学特征测量 在上述3个居群中各取30朵2种不同类型的红蓼,用测微尺分别测得它们的雌、雄蕊高度,雌雄蕊高度差,花瓣长度。柱头的宽度在显微镜(Olympus BH2)下测量,数据精确到0.01mm。
1.2.2 扫描电镜与柱头的石蜡切片观察 取新鲜标本,将柱头、花粉粒和用丙酮酸清洗过的饱满种子,用导电胶带粘于进样台上,在日立牌扫描电子显微镜(HitachiS4800)下观察。
取盛开的2种不同类型的红蓼各5朵,将长短柱头分别固定于FAA中48h以上,采用石蜡切片技术[11]对柱头进行观察。
1.2.3 花粉管萌发观察 取发育良好且柱头未经授粉的红蓼长短柱花隔离昆虫进行人工授粉,人工授粉时将整个柱头涂抹一层花粉。设置以下几个实验组,型内异交:红蓼长柱花♀×红蓼长柱花♂,红蓼短柱花♀×红蓼短柱花♂;型间异交:红蓼长柱花♀×红蓼短柱花♂,红蓼短柱花♀×红蓼长柱花♂;自交:红蓼长柱花自花授粉,红蓼短柱花自花授粉。每个组各取6朵花,将授粉后的花朵按0.5,1,2,3h不同时间段处理后置于FAA固定24h以上,用脱色苯胺蓝荧光染色法制片[12],在荧光显微镜(Olympus BH2)下,观察拍照。
1.2.4 野外调查座果率与人工辅助授粉 在野外调查时,红蓼居群大小用其开花数量衡量。在上述3个居群中每个居群随机选取4个样方,样方大小为5m×5m,每个样方随机选取5个花序统计座果率,分别取平均值。样方内全为同型花则是单型居群,两种花型都有则为双型居群。
人工辅助授粉,设计处理:长柱花自花授粉,套袋;短柱花自花授粉,套袋;长柱花授同型花粉,套袋;短柱花授同型花粉,套袋;长柱花去雄授短柱花花粉,套袋;短柱花去雄,授长柱花花粉。选取5个花序,每个花序10~20个花。
1.2.5 访花者的观察 2014年8月20日和2014年9月6日分别对芜湖市安徽师范大学花津校区和镜湖区的红蓼居群进行观察,上午8时开始观察到18时,每隔1h记录一次访花者。
1.3 统计分析
采用统计软件SPSS 16.0和Excel进行数据处理和显著性检验分析。
2 结果与分析
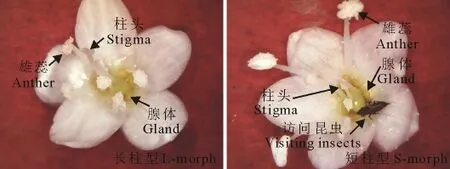
图1 红蓼的长、短柱型花Fig.1 The long and short morphs of P.orientale
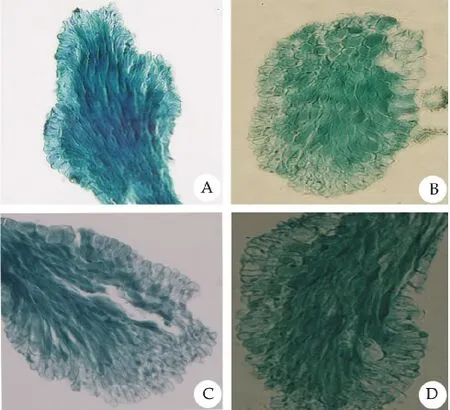
图2 红蓼的石蜡切片图Fig.2 The paraffin section of P.orientale
2.1 红蓼二型花柱的特征
红蓼的花部生物学特征显示,红蓼为两型花,有长柱型花和短柱型花两种类型,花瓣5片,为长椭圆形,花冠辐射对称,雄蕊7枚,其中5枚与花瓣互生,着生于花瓣基部,子房基部生有腺体(图1)。长柱型柱头高度高于雄蕊(表1,t=37.624,P<0.001),短柱型柱头高度低于雄蕊(表1,t=57.210,P<0.001);长柱型柱头宽度小于短柱型(表 1,t=14.895,P<0.001);长柱型花粉数目多于短柱型(表1,t=7.531,P<0.05),但花粉大小小于短柱型(表1,t=4.847,P<0.001);长柱型与短柱型柱头乳突细胞的宽度差异不显著(表1,图2,t=0.416,P>0.05),长柱型具有长而稀的柱头乳突细胞,短柱型柱头乳突细胞短且排列紧密(表1,图2A~D,图3C,F;t=13.038,P<0.001)。长柱型和短柱型花粉粒均为球型,具有散孔,孔近似圆形,边缘形状不规则(图3G~J)。
2.2 座果率与人工辅助授粉
从野外采集到的种子来看,红蓼种子黑色,有光泽,扁平卵圆形,双凹,顶端渐狭,在扫描电镜下观察到表面凹凸不平(图3K~N)。
在野外条件下随机统计其座果率,观察到单双居群间座果率有差异,双居群座果率高于单居群,无论单居群还是双居群,短柱型座果率均高于长柱型(表2)。
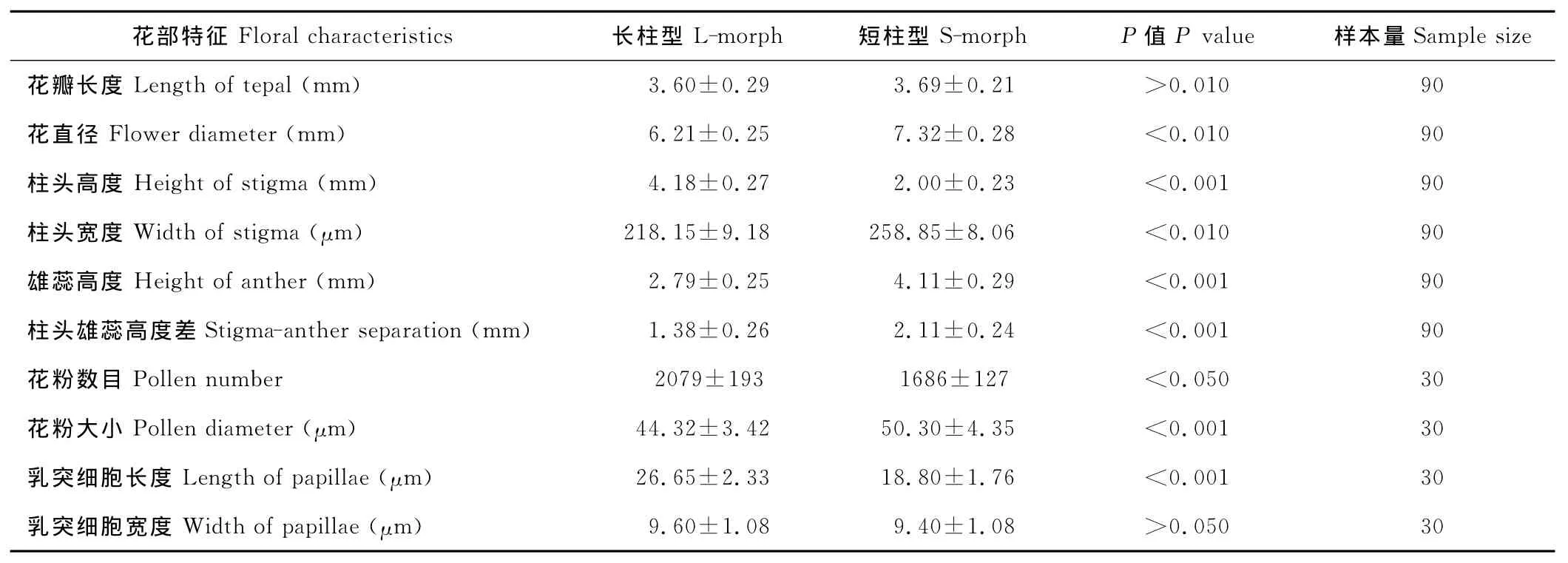
表1 红蓼长、短柱花的花部特征 (平均值±标准差)Table 1 Measurement of the dimensions of flowers and pollen of long and short-morph flowers of P.orientale(mean±standard deviation)
自花授粉和型内异花授粉都有少数种子,表明红蓼自交和型内异花授粉具有一定的亲和性。型间异花授粉座果率略低于自然状态下的座果率(表3)。
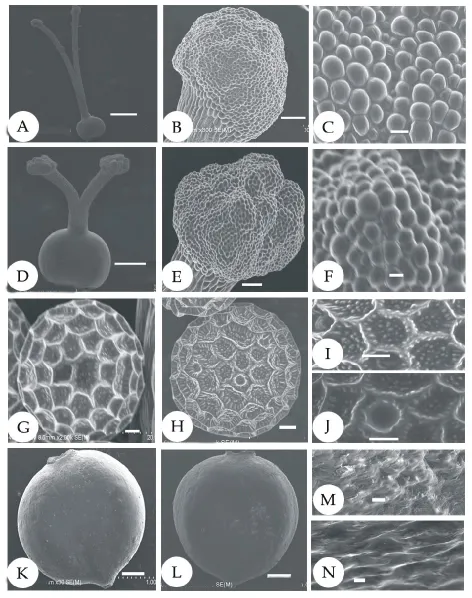
图3 红蓼的微形态特征(SEM)Fig.3 Micromorphological characteristics of P.orientale scanning electron microscopy
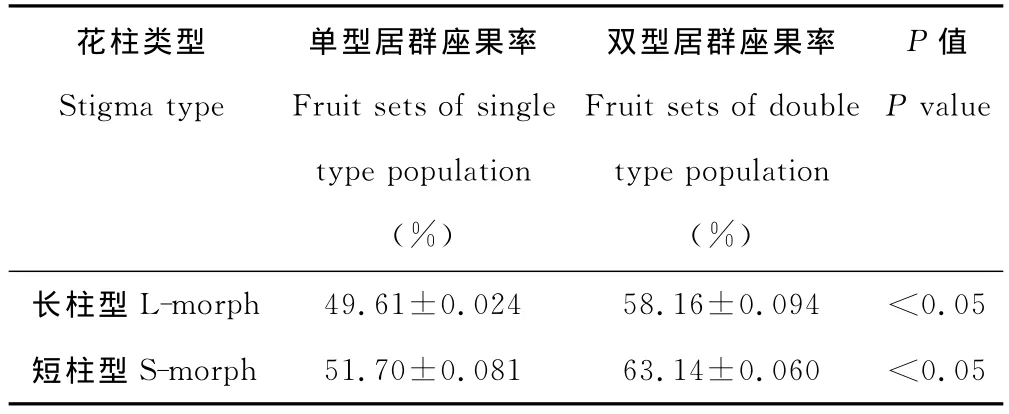
表2 红蓼自然条件下双居群、单居群的座果率 (平均值±标准差)Table 2 Fruit sets of P.orientalein different population in natural condition(mean±standard deviation)
2.3 访花昆虫行为
红蓼的花期为6-9月,果期为8-10月,盛花期为8月,颜色鲜艳,通过野外观察,并没有发现鸟雀和蝙蝠的访问,但发现至少12种访花昆虫,其访花频率见表4,部分访花者见图4,访花者中蓟马、食蚜蝇以及蚂蚁最为频繁。
2.4 花粉管萌发的研究
自交和异交,授粉后,花粉在0.5h内萌发,型内异交、自交以及型间异交的花粉管萌发并无明显差别,0.5h已能够到达子房(图5F,J,L,Q,R)。花粉管萌发时首先多数花粉管伸入中央花柱道(图5A,D,G,K,O),只有少数花粉管沿着花柱壁,到达花柱基部(图5C,I,M,O,P,R),沿子房壁到达胚珠基部的合点而达胚囊,受精方式为合点受精(图5F,J,N,Q,R)。
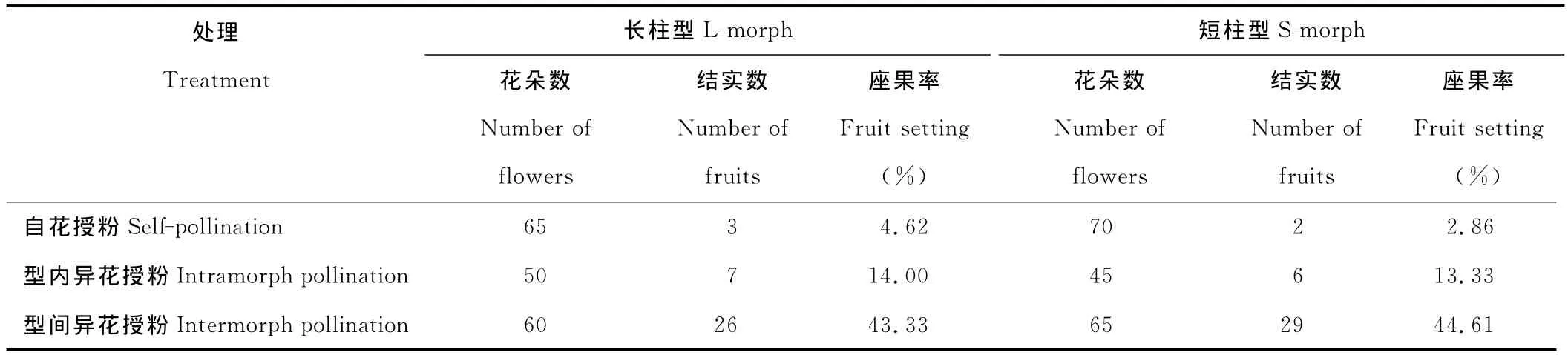
表3 红蓼人工辅助授粉的座果率Table 3 Fruit sets of P.orientalein different population under artificial pollination

图4 红蓼的花序和访花者Fig.4 The florescence of P.orientale and visiting insects on the flowers
荧光实验中观察到花粉管在萌发过程中的行为:1)并不是所有花粉管都能萌发至子房,只有少数花粉管能够到达子房。2)某些花粉管在伸长的过程中边伸长边交汇,最终交汇成一条或几条花粉管沿子房壁到达胚珠基部合点。3)在荧光显微镜下观察到花粉管为密集波浪线形。
3 讨论
异型花柱是由基因控制的花部多态现象,有4种类型,包括二型花柱、三型花柱、柱头高度两型性(stigma-height dimorphism)和花柱对生(enantiostyly,有异常雄蕊存在),它们通过提高植物个体间花粉散布的精确性来增加雄性的交配效率[13-14],其中二型花柱有两种类型的花,即长柱花和短柱花,三型花柱则有3种类型的花,即长柱花,中柱花和短柱花[15]。已被发现蓼属植物有300多种,其花部具有多态现象,已被报道的蓼属植物,如金荞麦(Fagopyrum dibotrys)[16],愉悦蓼[10],蚕茧草[9]等都是典型的二型花柱,不同的蓼属植物它们的花柱数目不同,在蚕茧草中不但有两个柱头的花,同时存在3个柱头的花。红蓼是典型的二型花柱植物,通过野外调查只发现了两个柱头的花,在多地的采样中并未发现红蓼的三柱头的花。
大多数二型花柱植物的长柱型花要小于短柱型花,普遍认为不同花型的花部特征在一定程度上互补能够起到调和雌雄功能的作用[14],短花柱型的花柱头位置比长花柱型的花低,比长柱花较难接受到花粉,因此通过增大花型来增加对传粉昆虫的吸引[15],从而增加传粉效率。已研究的蓼科植物中金荞麦[16]的短柱花比长柱花大,对红蓼两种花型花直径的测量符合上述观点。红蓼的柱头、花药高度、柱头形态以及花粉粒大小均呈现二态性(表1)。长柱型柱头宽度窄于短柱型,表皮细胞乳突较长且排列疏松,显然能够接受大的短柱型花粉。长柱型的花粉小而多也是对位置较低的短柱型柱头较难接受到花粉的适应。花柱的多态性能够提高其繁殖适合度。短柱型柱头表皮细胞乳突相对较多从而提高花粉黏附效率[17],较大的花粉为萌发时通过长柱型花柱提供更多的能量[18-19]。这种典型的二型花柱特征能够促进同配效率,符合资源分配原则。
在异型花柱的花中,同配(legitimate union)通常是可育的,异配(illegitimate union)往往可育性低或者是不可育的。植物的传粉过程具有高度专一化的特点[20],交互式雌雄异位是二型花柱的典型特征,传粉者身体携带的花粉只能接触同一高度的花器官,从而避免雌雄器官的性别干扰并有利于传粉者实现精确传粉并提高同配效率[2],在自然条件下,我们发现双居群座果率高于单居群,这说明了杂交对于花粉管萌发的优势,也说明了同型的花粉并不是完全可育的,异型花粉的结实率高于同型花粉,这与Darwin[21]的观点一致。
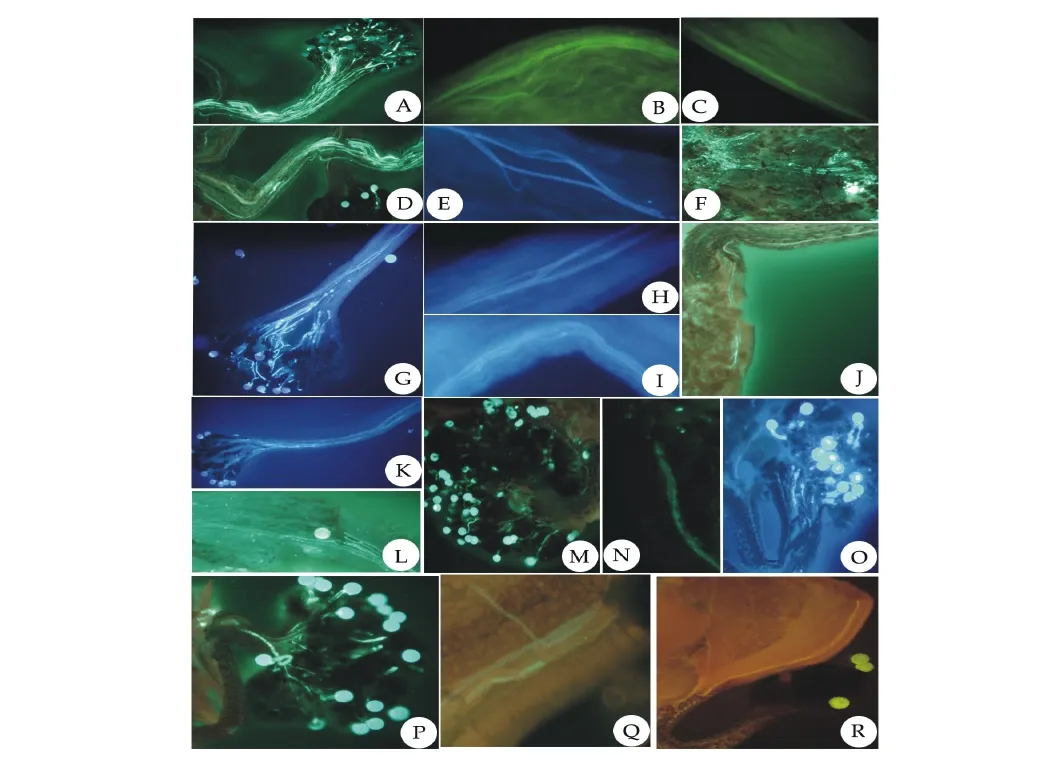
图5 红蓼人工授粉处理后花粉管体内生长图Fig.5 Pollen tube growth in vivo of P.orientale following self-,intramorph and intermorph pollination
异型自交不亲和系统(self-incrompatibility,SI)是异型花柱植物避免自交及型内异交,同时促进型间异交的机制。传粉后的自交不亲和机制与配子体的选择有关[13]。二型花柱常常伴有自交不亲和系统,但是不同植物中又表现出不同程度的可育性。并不是所有的二型花柱在自交表现为完全自交不亲和,不同植物自交亲和性不同,艳丽耳草(Hedyotis pulcherrima)[22]、绣球茜(Dunnia sinensis)[19]自交表现出完全不亲和性,唇形科(Labiatae)的Salvia brandegeei[22]、千蕨菜科(Lythraceae)的Decodon verticillatus[23]表现出自交亲和,在已经研究的蓼科植物中金荞麦[16]自交不亲和但型内异交具有一定的亲和性,本研究中的红蓼自交和型内异交均具有一定的亲和性,但以异交为主。
通过荧光显微镜的观察,发现不同处理授粉后的花粉均可萌发,花粉管中胼胝质分布较为均匀,0.5h内自交、型内异交以及型间异交均有花粉管萌发至子房,这与人工授粉的结果一致,自花授粉以及型内异花授粉结实率低,表明红蓼自交及型内异交具有一定的亲和性。花粉粒富含淀粉,淀粉是花粉管生长的主要能量[24]。在荧光下观察到花粉管生长过程中具有汇合现象,有利于能量的聚集,使花粉管最大限度伸长,只有部分花粉管能够沿子房壁到达胚珠,完成合点受精。
主要访花者为蓟马、蚂蚁和食蚜蝇。一般认为蚂蚁在传粉方面并不起作用,蚂蚁口器用于吸收花蜜,蚂蚁身体表面的抗菌物质减弱花粉粒的附着[25]。蓟马与食蚜蝇是否为有效传粉者需进一步研究。
综上所述,红蓼具有互补式雌雄异位,且花冠形态、柱头大小、花粉数目及大小等方面均表现出二型性的特点,野外观察到红蓼常常以单居群存在,且双居群座果率大于单居群,荧光实验观察到红蓼的自交、型内异交及型间异交花粉管萌发且0.5h内均有少量花粉管萌发至子房,为合点受精方式。人工授粉时自交及型内异交结实率低,是不是花粉管萌发至子房时子房对自交及型内异交具有拒绝作用,这个问题也有待进一步研究。
[1]Barrett S C H.Heterostylous genetic polymorphisms:model systems for evolutionary analysis.Evolution and Function of Heterostyly,1992,15:1-29.
[2]Barrett S C H.The evolution of plant sexual diversity.Nature Reviews Genetics,2002,3:274-284.
[3]Ganders E.The biology of heterostyly.New Zealand Journal of Botany,1979,17:607-635.
[4]Tom Beeckman.Pollen deposition rates and the functioning of distyly in the perennial Pulmonaria officinalis(Boraginaceae).Plant Systematics and Evolution,2008,273:1-12.
[5]Cardoso de Castro C,Cardoso Araujo A.Distyly and sequential pollinators of Psychotria nuda (Rubiaceae),in the Atlantic rain forest,Brazil.Plant Systematics and Evolution,2004,244:131-139.
[6]Chen M L,Liu D Y,Liu X Y.Observation of micromorphological characters of five species in Oxalis.Journal of Plant Resources and Environment,2007,16(3):7-18.
[7]Reddy N P,Bahadur B,Kulnar P V.Heterostyly in PolygonumChinese.Journal of Geneties,1977,63(2):79-82.
[8]Chen M L,Zhang X P.Distyly in Polygonum jucundum Meisn.(Polygonaceae).Plant Systematics and Evolution,2010,288:139-148.
[9]Lv F J,Cui M C,Chen M L.Reproductive biology of Polygonum japonicum.Acta Prataculturae Sinica,2013,22(3):196-203.
[10]Cui M C,Lv F J,Chen M L.Reproductive ecology of the distylous species of Polygonum japonicum.Bulletin of Botanical Research,2013,33(4):429-434.
[11]Hu S Y.Method of preparation of slides used to examine the pollen germination on the stigma and pollen tube growth in the style.Chinese Bulletin of Botany,1994,11:58-60.
[12]Li Z L.Plant Production Technology[M].Beijing:Science Press,1978.
[13]He Y P,Liu J Q.A review on recent advances in the studies of plant breeding system.Acta Phytoecologica Sinica,2003,27(2):151-163.
[14]Barret S C H,Jesson L K,Baker A M.The evolution and function of stylar polymorphisms in flowering plants.Annals of Botany,2000,85(Supp.A):253-265.
[15]Chen M L,You Y L,Zhang X P.Advances in the research of heterostyly.Acta Prataculturae Sinica,2010,19(1):226-239.
[16]Zhang W L,Zhou B,Xiao Y A,et al.Reproductive ecology of distylous Fagopyrum dibotrys.Acta Botanica Boreali-Occidentalia Sinica,2013,33(3):483-493.
[17]Ree R H.Pollen flow,fecundity,and the adaptive significance of heterostyly in Palicourea padifolia (Rubiaceae).Biotropica,1997,29(3):298-308.
[18]Gander F R.The biology of heterostyly.New Zealand Journal of Botany,1979,17(4):607-635.
[19]Zhong Z B,Luo S X,Li A M,et al.Distyly and pollination biology of Dunnia sinensis (Rubiaceae).Journal of Tropical and Subtropical Botany,2009,17(3):267-274.
[20]Guo H C,Yan C,Wei Y.Study on the flowering dynamic,pollen viability and stigma receptive of Kochia prostrate.Acta Prataculturae Sinica,2014,23(4):87-89.
[21]Darwin C.The Different Forms of Flowers on Plants of the Same Species[M].London:Applelon & Compan,1877.
[22]Barrett S C H,Wilken D H,Cole W W.Heterostyly in the Laminaceae:the case of Salvia brandegeei.Plant Systematics and Evolution,2000,233:211-219.
[23]Eckert C G,Barrett S C H.Tristyly,self-compatibility and floral variation in Decodon verticillatus (Lythraceae).Biological Journal of the Linnean Society,1994,53(1):1-30.
[24]Liu X X,Wu X Q,Zhang D X.Distyly and heteromorphic self-incompatibility of Hedyotis pulcherrima (Rubiaceae).Biodiversity Science,2012,20(3):337-347.
[25]Beattie A J,Knox R B,Williama E G.Ant inhibition of pollen function:apossible reason why ant pollination is rare.American Journal of Botany,1984,71(3):421-426.
参考文献:
[9]吕奉菊,崔美辰,陈明林.蚕茧草的繁殖生物学研究.草业学报,2013,22(3):196-203.
[10]崔美辰,吕奉菊,陈明林.二型花柱植物愉悦蓼的繁殖生态学研究.植物研究,2013,33(4):429-434.
[11]胡适宜.检查花粉在柱头上萌发和花粉管在花柱中生长的制片法.植物学通报,1994,11:58-60.
[12]李正理.植物制片技术[M].北京:北京科学出版社,1978.
[13]何亚平,刘建全.植物繁育系统研究的最新进展和评述.植物生态学报,2003,27(2):151-163.
[15]陈明林,游亚丽,张小平.花柱异型研究进展.草业学报,2010,19(1):226-239.
[16]张万灵,周兵,肖宜安,等.二型花柱植物金荞麦的繁殖生态学研究.西北植物学报,2013,33(3):483-493.
[19]钟智波,罗世孝,李爱民,等.绣球茜的二型花柱及其传粉生物学初步研究.热带亚热带植物学报,2009,17(3):267-274.
[20]郭红超,严成,魏岩.木地肤的开花动态与花粉活力及柱头可授性研究.草业学报,2014,23(4):87-89.
[24]刘欣欣,吴小琴,张奠湘.艳丽耳草的二型花柱及异型自交不亲和系统.生物多样性,2012,20(3):337-347.
猜你喜欢
西北植物学报(2023年9期)2023-10-17 13:34:00
作物学报(2022年1期)2022-11-05 08:06:16
新传奇(2021年12期)2021-04-25 17:55:42
中国果树(2020年2期)2020-07-25 02:14:16
数学小灵通(1-2年级)(2018年12期)2018-12-24 13:16:12
中国蔬菜(2018年3期)2018-03-08 03:19:00
建筑科学与工程学报(2017年2期)2017-04-07 15:43:36
建筑科学与工程学报(2016年2期)2016-04-20 21:02:21
湖南大学学报·自然科学版(2015年9期)2016-01-06 15:03:14
湖北农业科学(2015年16期)2015-10-28 22:11:58
