
细粒棘球蚴原头蚴包囊和原头蚴成虫抑制差减杂交文库中差异表达基因的筛选及分析
2015-03-27 10:34:39石保新张壮志马正海金映红张文宝
畜牧兽医学报 2015年7期
赵 莉,石保新,李 军,张壮志,马正海,张 旭,薛 晶,金映红,张文宝*
(1.新疆畜牧科学院兽医研究所,乌鲁木齐 830000;2.新疆大学生命科学与技术学院重点实验室,乌鲁木齐 830046;3.新疆医科大学第一附属医院医学研究中心,新疆包虫病基础医学重点实验室,乌鲁木齐 830054)
细粒棘球蚴原头蚴包囊和原头蚴成虫抑制差减杂交文库中差异表达基因的筛选及分析
赵 莉1#,石保新1#,李 军3,张壮志1,马正海2,张 旭1,薛 晶1,金映红1,张文宝3*
(1.新疆畜牧科学院兽医研究所,乌鲁木齐 830000;2.新疆大学生命科学与技术学院重点实验室,乌鲁木齐 830046;3.新疆医科大学第一附属医院医学研究中心,新疆包虫病基础医学重点实验室,乌鲁木齐 830054)
细粒棘球绦虫原头蚴(PSC)具有双向发育的特点,本文旨在利用SSH和生物信息学方法筛选原头蚴包囊(CW)和成虫(AW)特异性的基因。将PSC-CW和PSC-AW双相发育差减cDNA文库的差减PCR产物克隆入pGEM-T载体并测序,利用CAP3 sequence assembly在线软件、Blast2GO软件与GenBank数据库对测序结果进行分析,筛选原头蚴成囊和成虫特异性的基因,并探讨这些基因的功能。结果显示:对PSC-CW和PSC-AW文库进行扩增,分别得到280和200个阳性克隆,菌落PCR鉴定结果表明,这些克隆均插入200~1 000 bp片段。测序结果显示从PSC-CW和PSC-AW SSH文库中分别得到16和10个特异性基因。其中PSC-CW SSH文库中得到4个出现频率较高的基因,其余12个基因仅出现1次,其中5个为未知基因,已知基因分别编码细胞色素C氧化酶I、热休克蛋白70和铁蛋白等功能蛋白质。PSC-AW SSH文库中得到10个基因,其中2个出现频率较高基因,8个基因仅出现1次,4个为未知基因,6个已知基因分别编码基质蛋白1、延伸因子1α和脂肪酸结合蛋白等功能蛋白质。本研究筛选获得原头蚴成囊和成虫发育时差异表达的基因,对这些基因的功能进行分析表明,PSC-CW SSH文库中多是与营养和能量转运功能的相关基因,而PSC-AW SSH文库中多为发育与分化相关的基因,这些基因的差异表达可能与原头蚴处于不同的生长环境和发育方向有关,同时也为棘球蚴病免疫诊断、药物筛选和疫苗研制提供候选基因。
细粒棘球绦虫;抑制差减杂交;特异性基因
细粒棘球蚴病,又称为包虫病,是由细粒棘球绦虫引起的一种严重的人畜共患病[1]。细粒棘球绦虫原头蚴具有独特的双向性发育的能力,可以在中间宿主体内发育为包囊,进而完成生活史的无性繁殖(继发性感染),若中间宿主带包囊的脏器被终末宿主(犬等)采食,原头蚴在犬小肠中发育为细粒棘球绦虫成虫,这又构成了原头蚴有性繁殖(图1),若能寻找到相关的特异性基因,就可以切断其循环史。随着分子生物学技术的发展和对寄生虫虫体发育学的深入认识,人们可以筛选出在虫体不同发育阶段特异性表达的基因,以研究这些基因在细胞分化过程中的作用及相互关系,为寄生虫病的免疫诊断、药物筛选和疫苗研制提供候选基因。抑制差减杂交(suppression subtractive hybridization,SSH)是一种高效鉴定和克隆差异表达基因的新技术[2]。SSH在许多方面具有其独特的优越性:该方法操作简便、周期短,且阳性率高,达94%以上[3],有利于快速建立基因表达图谱,从整体上了解生物(细胞)在特定时期或特定环境下的基因表达动态;该方法灵敏度高,可使低丰度的mRNA得以高于1 000倍的富集,较其他方法更易于在转录水平上研究特定细胞或组织的基因表达差异[4]。本研究利用细粒棘球绦虫生长发育的特殊生物学特点,以幼虫(原头蚴)双相发育为切入点。利用SSH分离原头蚴特异的成囊及成虫基因,并结合生物信息学(bioinformatics)技术对获得基因进行分析,以筛查细粒棘球蚴双相发育相关基因,为研究细粒棘球蚴双相发育现象奠定基础。

图1 细粒棘球绦虫原头蚴的双相发育Fig.1 Bidirectional development of the protoscolex of E.granulosus
1 材料与方法
试验于2009—2010年在新疆畜牧科学院兽医研究所完成。
1.1 SSH文库
1.1.1 PSC-CW SSH文库 以细粒棘球绦虫原头蚴为驱动相,包囊为实验相,构建的原头蚴包囊抑制差减杂交文库,由新疆畜牧科学院兽医研究所寄生虫研究室提供。
1.1.2 PSC-AW SSH文库 以细粒棘球绦虫原头蚴为驱动相,成虫为实验相,构建的原头蚴成虫抑制差减杂交文库,由新疆畜牧科学院兽医研究所寄生虫研究室提供。
1.2 菌种和质粒
pGEM-T easy vector购自Promega公司;pMAL-p2x载体由本室保存,大肠杆菌DH5α和BL21由本室保存。其他试剂均为国产分析纯产品。
1.3 生化及分子生物学试剂
PCR 片段回收试剂盒,胶回收试剂盒均购自北京天根生化科技有限公司;ExTaqDNA聚合酶、DNA marker、T4DNA 连接酶、限制性内切酶购自TaKaRa公司;硝酸纤维素膜及3 mm滤纸均为德国 Whatman 公司产品;其余常规试剂为国产分析纯试剂。
1.4 引物设计与合成
根据pGEM T easy vecter载体序列合成以下引物,T7 primer:5′-TAATACGACTCACTATAGGG-3′;M13 reverse primer:5′-AGCGGATAACAATTTCACACAGGA-3′,引物由上海生工公司合成。
1.5 目的片段与载体的连接反应
将原头蚴包囊和原头蚴成虫的SSH文库的PCR产物与pGEM T载体于T4DNA连接酶作用下,4 ℃连接过夜,将连接产物转化大肠杆菌DH5α感受态细胞,涂在含Amp+(75 μg·mL-1)琼脂平板上,37 ℃,正置20 min后,倒置培养过夜。
1.6 SSH文库的鉴定
用牙签挑取单菌落接入少量LB培养基(Amp+)中,摇床振荡培养过夜,取少量细菌为模板进行PCR扩增。PCR扩增参数:94 ℃预变性5 min;94 ℃变性30 s,60 ℃退火45 s,72 ℃延伸1 min,共进行30个循环;72 ℃延伸7 min。在0.7%的琼脂糖凝胶电泳分析扩增产物后,用DNA凝胶回收试剂盒对PCR产物进行切胶纯化。
1.7 SSH文库测序与序列分析
将菌落PCR扩增片段在200~1 000 bp的克隆分别挑选100个克隆送上海博尚生物有限公司进行测序。PSC-CW和PSC-AW SSH文库测序成功的序列分别为86和80个序列。将测序结果去除载体和引物序列以后,首先利用CAP3 sequence assembly在线软件对序列的正反向进行比对,获得高表达基因和单一基因,然后利用BLAST2GO在线软件与GenBank数据库进行同源序列比对分析和已知序列在生物学过程、分子功能和细胞组分方面所具有的功能。
2 结 果
2.1 消减文库插入片段的PCR鉴定
将PSC-CW和PSC-AW文库的PCR产物分别插入pGEM-T easy vector,转化DH5α感受态细胞,分别得到280和200个阳性克隆,以pGEM- T easy vecter通用引物进行菌落PCR扩增,均能扩增出大小不等的插入片段,电泳结果显示插入片段大小主要分布在 200~1 000 bp,这些条带可能代表差异表达的基因片段 (图2、3)。

M.DL2000 DNA相对分子质量标准;1~46.cDNA插入片段的PCR产物M.DNA marker DL2000;1-46.The PCR products of cDNA inserts图2 PSC-CW SSH文库的部分PCR扩增结果Fig.2 PCR analysis of the PSC-CW SSH library
2.2 PSC-CW文库的cDNA测序与序列分析初步结果
2.2.1 序列同源性比较 将文库送至上海博尚生物有限公司,进行全文库单向测序。利用CAP3 sequence assembly在线软件进行正反向序列分析,结果显示,共获得16个基因,其中有4个为出现频率较多基因,12个基因仅出现1次,用Blast2GO在线软件与GenBank数据库进行同源性比较,结果显示11个基因是已知基因,5个未知基因,PSC-CW文库测序结果见表1。

表1 文库中扩增基因的分析

M.DL2000 DNA相对分子质量标准;1~46.cDNA插入片段的PCR产物M.DNA marker DL2000;1-46.The PCR products of cDNA inserts图3 PSC-AW 文库的部分PCR扩增结果Fig.3 PCR analysis of the PSC-AW SSH library
2.2.2 功能分析 对差异表达基因编码的蛋白质按照生物学过程(biological process)、分子功能(molecular function)和细胞组分(cellular component)水平进行功能分类分析。按生物学过程水平分析(图4),其功能主要涉及细胞分化和凋亡(GO:0009987)、细胞内大分子等物质代谢(GO:0008152)、蛋白转运与定位(GO:0051179)、电子传递及细胞内环境调节(GO:0065007)、应激反应(GO:0050896)以及编码细胞构成成分(GO:0016043)等。在分子功能水平分析(图5),它们主要具有核苷酸、阴离子、血红素等的结合能力(GO:0005488)、氧化还原酶类的催化活性(GO:0003824)、核糖体的组成成分(GO:0005198)、基质跨膜转运活性(GO:0005215)和氢、铁等离子载体活性(GO:0009055)等功能。按照细胞组分水平分析(图6),这些基因编码蛋白能够构成细胞膜(GO:0005623)、线粒体、细胞质等器官(GO:0043226)和细胞复杂的大分子物质等(GO:0032991)。
2.3 PSC-AW文库的cDNA测序与序列分析初步结果

图4 基于生物学过程水平对PSC-CW文库序列进行分析Fig.4 The analysis of the clone of the PSC-CW SSH library at biological process level

图5 基于分子功能水平对PSC-CW文库序列的分析Fig.5 The analysis of the clone of the PSC-CW SSH library at molecular function level
2.3.1 序列同源性比较 将文库送至上海博尚生物有限公司,进行全文库单向测序扫描。根据CAP3 sequence assembly在线软件进行正反向序列分析,结果显示,共获得10个基因,其中2个为出现频率较多的基因,8个基因仅出现1次,用Blast2GO在线软件与GenBank数据库进行同源性比较,结果显示6个基因是已知基因,4个未知基因,PSC-AW文库测序结果列于表1。
2.3.2 功能分析 将上述已知序列在生物过程水平上进行分析(图7),其编码的蛋白质涉及细胞大分子物质的合成等生物合成过程(GO:0009987),细胞内大分子、蛋白质等的代谢过程(GO:0008152),细胞营养成分转运等(GO:0051179)。 在分子功能水平上进行分析(图8),结果显示,它们主要具有核酸、脂质体等的结合作用(GO:0005488),水解酶活性(GO:0003824),RNA构成活性(GO:0005198),翻译延伸因子的转运活性(GO:0005215)等功能。按照细胞组分水平分析(图9),这些基因编码蛋白质可构成宿主细胞的细胞核(GO:0005623)、细胞膜(GO:0005576)等成分。

图6 基于细胞成分水平对PSC-CW文库序列的分析Fig.6 The analysis of the clone of the PSC-CW SSH library at cellular component level 2

图7 基于生物过程水平对PSC-AW文库序列进行分析Fig.7 The analysis of the clone of the PSC-AW SSH library at biological process level

图8 基于分子功能水平对PSC-AW文库序列进行分析Fig.8 The analysis of the clone of the PSC-AW SSH library at molecular function level
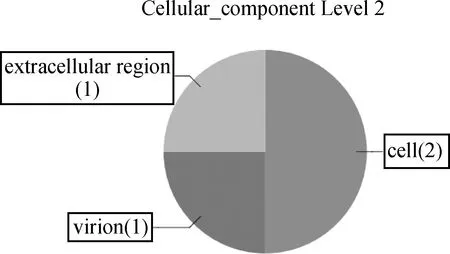
图9 基于细胞成分水平对PSC-AW文库序列的分析Fig.9 The analysis of the clone of the PSC-AW SSH library at cellular component level
3 讨 论
寄生虫的生活史比较复杂,需经过几个不同发育阶段来完成整个生命周期,每个阶段的虫体均有不同的形态结构和生理特点,需要不同的生活条件。因此,每个发育阶段都有其特异表达的基因。目前,已经发现了一些危害较为严重的寄生虫不同发育阶段的差异表达基因。细粒棘球绦虫的生长和发育是一个复杂的过程,存在4个发育阶段,包虫囊可大如西瓜、成虫(绦虫)细小而分节、原头蚴及虫卵期(六钩蚴)仅能借助于显微镜才能看到,它们的生长发育、形态、行为和对环境的需求方面都有很大程度的不同。其原头蚴和包囊是在羊等中间宿主体内生长发育的,而成虫和虫卵是在犬等终末宿主体内生长发育的。在实验室条件和自然宿主体内,诱导细粒棘球蚴原头蚴分别向包囊和成虫方向分化生长,差异不显著。在中间宿主体内,原头蚴是在肝、肺等内脏器官中发育成包囊,而在终末宿主体内,原头蚴寄生在肠道。本研究在实验室条件下,利用培养基中成分的不同,将原头蚴培养成包囊,同时,模拟犬肠道内环境,将原头蚴培养成成虫。但是细粒棘球蚴原头蚴分别向包囊和成虫方向分化生长过程中出现的阶段性、差异性以及各发育阶段产生的特性的内在机制尚不清楚,有待进一步研究。
SSH技术是1996年由L.Diatchenko等[2]在抑制性PCR和DNA差减杂交方法的基础上,建立的一种分离和鉴定不同细胞或不同基因型差异表达基因和低丰度基因的方法。本实验室曾经进行过相关试验,但试验结果不理想,测序结果显示,大部分基因都是终末宿主犬体内的基因。在本研究中,首次以细粒棘球绦虫幼虫原头蚴为驱动相,分别以包囊和成虫为实验相,获得了PSC-CW和PSC-AW两个差减文库,继而将差异PCR产物连接到pGEM-T 载体上,对其进行测序和同源性分析。
对PSC-CW文库的测序结果进行初步分析(表1),结果表明得到了16个基因序列,其中已知基因11个,出现频率较高的有3个基因,分别编码热休克蛋白70(HSP70)、细胞色素C氧化酶I和类质粒复制起始蛋白,其他基因均只出现过一次,分别编码线粒体蛋白、细胞色素C氧化酶Ⅲ、铁蛋白等功能和调控蛋白。其中,EPCf08与编码铁蛋白的基因序列相似性为80.75%,铁蛋白是原核生物和真核生物体内储存铁的一种蛋白质[5]。EPCContig4与编码热休克蛋白70(HSP70)的基因序列相似性为93.1%,HSP70是细胞水平上的一种分子监控蛋白,参与蛋白质合成和细胞内囊泡的转折和装配[6],而且此蛋白在许多原虫感染过程中具有很高的免疫原性[7-9]。鉴于HSP70的功能,我们进行了克隆表达,进一步探讨其在包虫病防治方面可能的应用前景。EPCContig2,与编码细胞色素C氧化酶I(coxI)的基因序列相似性为97.05%,coxI基因已被广泛用来区分扁形动物的种间界限和系统发育的亲和力,它在不同物种的分类和系统发育研究中具有重要意义[9-12]。对于从形态学差异无法区分的物种,可通过coxI基因序列分析加以区分,并可从系统发育学的角度阐述这种物种的形成过程,可作为物种的基因标志[13-15]。EPCc06,与编码线粒体内膜移位酶Tim8的基因序列相似性为61.08%,线粒体内膜移位酶Tim8位于线粒体内膜,在部分营养成分进入线粒体的过程中起一定作用[16-18]。
上述PSC-CW文库中的基因序列,从生物过程水平分析,基因序列EPCf08编码的蛋白质在细胞内有运输金属离子等功能,如运输铁离子等二价和三价无机阳离子,从而维持细胞内平衡。基因序列EPCe12-2编码的蛋白质在线粒体中具有运输ATP、氧和铁离子等作用,在维持呼吸系统电子传递和磷酸化过程中起重要作用;基因序列EPCContig2编码细胞内氧化呼吸作用及代谢过程中能量产生所需的酶;基因序列EPCc06编码蛋白质导向和胞内蛋白转运相关的蛋白质;基因序列EPCe06 e08编码核糖体蛋白,在翻译及生物大分子的合成中起重要作用;基因序列EPCContig4编码的蛋白质能够调节生长素等激素的合成和代谢及对刺激物的反应。从细胞组分角度上,其大多基因编码的蛋白质参与合成线粒体、核糖体、细胞膜等生物器官。从分子功能上,其大多基因编码的蛋白与离子、氧、ATP转运有关,以维持细胞生长和代谢,其中基因序列EPCContig2和EPCe12-2具有细胞色素C活性,编码运输阳离子的跨膜蛋白;基因序列EPCf08编码的蛋白质能转运铁离子等阳离子和氧;基因序列EPCe06、EPCContig4编码的蛋白质具有RNA结合能力的结构分子。另外还有一部分克隆在GenBank中无法查到相对应的同源性基因,可能是一些新基因。
对PSC-AW文库的测序结果进行初步分析,得到10个基因,其中6个为已知基因,且都是出现频率较低的基因,分别编码脂肪酸结合蛋白、基质蛋白和碱性磷酸酶等。其中,PA5与编码脂肪酸结合蛋白的基因序列相似性为76.8%,脂肪酸结合蛋白接种动物后可诱导产生免疫保护[19]。有报道称其能诱导动物产生抗血吸虫的免疫保护效果,能够作为预防血吸虫病的候选疫苗[20]。在C.T.Fonseca等的研究中,脂肪酸结合蛋白能诱导小鼠产生免疫应答,在小鼠体内的荷虫量减少24%~33%,肝中的虫体数量减少27%~29%[21]。PA10与延伸因子1α(EF-1α)的基因序列相似性为98.85%,其在不同物种间具有高度保守性,可用于物种的种系发生分析,同时它还是一个多功能蛋白质,在翻译过程中起重要作用[22-24]。以往确定细粒棘球绦虫的亚类主要根据线粒体基因[25-30]。最近,研究者开始尝试用核酸延伸因子1α(EF-1α)基因的序列和类型来区分细粒棘球绦虫亚型[31-32]。M.Okamoto 等在细胞凋亡试验中发现EF-1在转录过程中起重要作用,并调节细胞凋亡[32]。PA71与编码基质蛋白1的基因序列相似性为89%,基质蛋白1 是细胞生长和发育过程中不可或缺的成分,它有诱导细胞分化的作用[33]。根据生化过程水平的分析,PA5与代谢产物的转运和细胞识别有关,PA10在基因翻译和表达以及细胞内蛋白质等大分子的合成和代谢等过程中发挥作用。分子功能水平分析表明,PA5、PA10、PA71和PA82能够编码一些调控蛋白,可调控信号转导或基因表达等过程。细胞成分水平分析表明,A10为细胞质的组成成分,PA71是细胞膜的组织成分。同样,PSC-AW文库中也测得一些未知基因。
原头蚴通过无性繁殖发育成包囊,这是一个缓慢的发育过程,3~6月才能发育成0.5 cm的包囊,原头蚴进入中间宿主绵羊等体内,寄生在宿主的肝、肺等脏器,从宿主体内获得营养成分,使自身得到繁殖,故需要营养转运和能量获取相关的蛋白质。本研究中,PSC-CW SSH文库筛选出的基因大多能编码和调控转运营养和能量的功能性蛋白质,如coxI、铁蛋白、coxⅢ和Tim8等,这与成囊期发育过程所需蛋白质相吻合。
原头蚴通过有性繁殖发育成成虫,原头蚴进入终末宿主犬等体内,寄生在小肠内,约45 d后,犬开始排泄虫卵,约10 d左右,大量虫卵都脱落了,故成虫发育较快,需要大量参与代谢的酶以及转运蛋白,同时也需要大量构成细胞的组分。我们的研究中,PSC-AW SSH文库中筛选获得的基因主要参与生物发育与分化,可促进生物体的快速发育,这与成虫阶段的快速发育相一致。
[1] MCMANUS D P,ZHANG W,LI J,et al.Echinococcosis[J].Lancet,2003,362(9392):1295-1304.
[2] DIATCHENKO L,LAU Y F,CAMPBELL A P,et al.Suppression subtractive hybridization:a method for generating differentially regulated or tissue-specific cDNA probes and libraries[J].ProcNatlAcadSciUSA,1996,93(12):6025-6030.
[3] GRUMOLATO L,ELKAHLOUN A G,GHZILI H,et al.Microarray and suppression subtractive hybridization analyses of gene expression in pheochromocytoma cells reveal pleiotropic effects of pituitary adenylate cyclase-activating polypeptide on cell proliferation,survival,and adhesion[J].Endocrinology,2003,144(6):2368-2379.
[4] REBRIKOV D V,BRITANOVA O V,GURSKAYA N G,et al.Mirror orientation selection (MOS):a method for eliminating false positive clones from libraries generated by suppression subtractive hybridization[J].NucleicAcidsRes,2000,28(20):E90.
[5] ERSFELD K,CRAIG P S.Cloning and immunological characterisation ofEchinococcusgranulosusferritin[J].ParasitolRes,1995,81(5):382-387.
[6] SHONHAI A,BOSHOFF A,BLATCH G L.The structural and functional diversity of Hsp70 proteins fromPlasmodiumfalciparum[J].ProteinSci,2007,16(9):1803-1818.
[7] JENSEN A T,CURTIS J,MONTGOMERY J,et al.Molecular and immunological characterisation of the glucose regulated protein 78 ofLeishmaniadonovani(1)[J].BiochimBiophysActa,2001,1549(1):73-87.
[8] JENSEN A T,ISMAIL A,GAAFAR A,et al.Humoral and cellular immune responses to glucose regulated protein 78- -a novelLeishmaniadonovaniantigen[J].TropMedIntHealth,2002,7(5):471-476.
[9] IWAGAMI M,MONROY C,ROSAS M A,et al.A molecular phylogeographic study based on DNA sequences from individual metacercariae ofParagonimusmexicanusfrom Guatemala and Ecuador[J].JHelminthol,2003,77(1):33-38.
[10] MORGAN J A,DEJONG R J,KAZIBWE F,et al.A newly-identified lineage of Schistosoma[J].IntJParasitol,2003,33(9):977-985.
[11] RAZO-MENDIVIL U J,LEN-RGAGNON V,PÉREZ-PONCE DE LEN G.Description of two new species of Glypthelmins Stafford,1905 (Digenea:Macroderoididae) in Rana spp.from Mexico,based on morphology and mtDNA and rDNA sequences[J].SystParasitol,2004,59(3):199-210.
[12] RAZO-MENDIVIL U,ROSAS-VALDEZ R,PÉREZ-PONCE DE LEN G.A new cryptogonimid (Digenea) from the Mayan cichlid,Cichlasoma urophthalmus (Osteichthyes:Cichlidae),in several localities of the Yucatán Peninsula,Mexico [J].JParasitol,2008,94(6):1371-1378.
[14] LANE N.Biodiversity:On the origin of bar codes[J].Nature,2009,462(7271):272-274.
[15] OGEDENGBE J D,HANNER R H,BARTA J R.DNA barcoding identifies Eimeria species and contributes to the phylogenetics of coccidian parasites (Eimeriorina,Apicomplexa,Alveolata)[J].IntJParasitol,2011,41(8):843-850.
[16] KOEHLER C M,LEUENBERGER D,MERCHANT S,et al.Human deafness dystonia syndrome is a mitochondrial disease[J].ProcNatlAcadSciUSA, 1999,96(5):2141-2146.
[17] CURRAN S P,LEUENBERGER D,SCHMIDT E,et al.The role of the Tim8p-Tim13p complex in a conserved import pathway for mitochondrial polytopic inner membrane proteins[J].JCellBiol,2002,158(6):1017-1027.
[18] BLACKSTONE C,ROBERTS R G,SEEBURG D P,et al.Interaction of the deafness-dystonia protein DDP/TIMM8a with the signal transduction adaptor molecule STAM1[J].BiochemBiophysResCommun,2003,305(2):345-352.
[19] BERGQUIST R,AL-SHERBINY M,BARAKAT R,et al.Blueprint for schistosomiasis vaccine development[J].ActaTrop,2002,82(2):183-192.
[20] CHUNCHOB S,GRAMS R,VIYANANT V,et al.Comparative analysis of two fatty acid binding proteins fromFasciolagigantica[J].Parasitology,2010,137(12):1805-1817.
[21] FONSECA C T,PACIFICO L G,BARSANTE M M,et al.Co-administration of plasmid expressing IL-12 with 14-kDa Schistosoma mansoni fatty acid-binding protein cDNA alters immune response profiles and fails to enhance protection induced by Sm14 DNA vaccine alone[J].MicrobesInfect,2006,8(9-10):2509-2516.
[22] FAVRET C,VOEGTLIN D J.Speciation by host-switching in pinyon Cinara (Insecta:Hemiptera:Aphididae)[J].MolPhylogenetEvol,2004,32(1):139-151.
[23] CHAVERRI P,BISCHOFF J F,EVANS H C,et al.Regiocrella,a new entomopathogenic genus with a pycnidial anamorph and its phylogenetic placement in the Clavicipitaceae[J].Mycologia,2005,97(6):1225-1237.
[24] XIAO N,QIU J,NAKAO M,et al.Echinococcusshiquicusn.sp.,a taeniid cestode from Tibetan fox and plateau pika in China[J].IntJParasitol,2005,35(6):693-701.
[25] NAKAO M,MCMANUS D P,SCHANTZ P M,et al.A molecular phylogeny of the genusEchinococcusinferred from complete mitochondrial genomes[J].Parasitology,2007,134(Pt5):713-722.
[26] HÜTTNER M,NAKAO M,WASSERMANN T,et al.Genetic characterization and phylogenetic position ofEchinococcusfelidis(Cestoda:Taeniidae) from the African lion[J].IntJParasitol,2008,38(7):861-868.
[27] THOMPSON R C,MCMANUS D P.Towards a taxonomic revision of the genusEchinococcus[J].TrendsParasitol,2002,18(10):452-457.
[28] THOMPSON R C.The taxonomy,phylogeny and transmission ofEchinococcus[J].ExpParasitol,2008,119(4):439-446.
[30] BADARACO J L,AYALA F J,BART J M,et al.Using mitochondrial and nuclear markers to evaluate the degree of genetic cohesion amongEchinococcuspopulations[J].ExpParasitol,2008,119(4):453-459.
[31] HAAG K L,GOTTSTEIN B,AYALA F J.The EG95 antigen ofEchinococcusspp.contains positively selected amino acids,which may influence host specificity and vaccine efficacy[J].PLoSOne,2009,4(4):e5362.
[32] OKAMOTO M,NAKAO M,BLAIR D,et al.Evidence of hybridization betweenTaeniasaginataandTaeniaasiatica[J].ParasitolInt,2010,59(1):70-74.
[33] NARAYANAN K,SRINIVAS R,RAMACHANDRAN A,et al.Differentiation of embryonic mesenchymal cells to odontoblast-like cells by overexpression of dentin matrix protein 1 [J].ProcNatlAcadSciUSA,2001,98(8):4516-4521.
(编辑 白永平)
The Screening and Analysis of Differentially Expressed Genes in Suppression Subtractive Hybridization Library with either Cyst (CW) or Adult Worm (AM) Development fromEchinococcusgranulosusLarval Protoscolex (PSC)
ZHAO Li1#,SHI Bao-xin1#,LI Jun3,ZHANG Zhuang-zhi1,MA Zheng-hai2,ZHANG Xu1,XUE Jing1,JIN Ying-hong1,ZHANG Wen-bao3*
(1.VeterinaryResearchInstituteofAnimalScienceAcademyofXinjiangUygurAutonomousRegion,Urumqi830000,China;2.XinjiangKeyLaboratoryofBiologicalResourcesandGeneticEngineering,CollegeofLifeScienceandTechnology,XinjiangUniversity,Urumqi830046,China;3.XinjiangHydatidFundamentalMedicalKeyLab,FirstTeachingHospitalofXinjiangMedicalUniversity,Urumqi830054,China)
According to the bidirectional development of the protoscolex (PSC) ofEchinococcusgranulosus,the study was conducted to screen the specifical genes ofE.granulosusPSC development between a larval cyst and adult worm.The subtractive PCR products of the SSH library ofE.granulosusPSC-adult (AW) andE.granulosusPSC-cyst (CW) were connected to the pGEM-T easy vector,and were cloned and sequenced.Totally,280 and 200 clones were obtained from PSC-CW and PSC-AM libraries,respectively,with the length of fragments 200 to 1000 bp.The clones were sequenced.Sequence alignment analysis showed that these sequences belong to 16 and 10 gene clusters representing 16 and 10 unique genes from PSC-CM and PSC-AW libraries respectively.As the genes are specifically or differentially expressed during the development of PSC to either adult worm or cyst,they are important for the parasite development and differentiation,thus can be candidates for drug and vaccine development.
Echinococcusgranulosus;suppression subtractive hybridization (SSH);the specifical genes
10.11843/j.issn.0366-6964.2015.07.019
2014-09-26
公益性行业(农业)科研专项项目(201103008)
赵 莉(1982-),女,江苏宿迁人,硕士,助理研究员,主要从事生物化学与分子生物学研究,E-mail:82285053@qq.com;石保新(1963-),男,河北馆陶人,研究员,E-mail:2423930978@qq.com。石保新与赵莉为同等贡献的第一作者
*通信作者:张文宝,研究员,E-mail:wenbaozhang2013@163.com
S852.734
A
0366-6964(2015)07-1215-09
猜你喜欢
选煤技术(2021年3期)2021-10-13 07:33:36
有色金属(矿山部分)(2021年4期)2021-08-30 06:10:26
猪业科学(2021年3期)2021-05-21 02:05:36
幽默大师(2020年10期)2020-11-10 09:07:22
中华诗词(2019年1期)2019-11-14 23:33:56
天津科技大学学报(2019年5期)2019-10-23 01:21:14
猪业科学(2018年4期)2018-05-19 02:04:31
西南石油大学学报(自然科学版)(2016年6期)2017-01-15 14:14:06
中国资源综合利用(2016年9期)2016-01-22 08:35:21
西藏科技(2015年6期)2015-09-26 12:12:12
