
环境因子对胡杨树干液流动态的影响
2015-03-15 02:41赵天宇关东海苏里坦时山良
水土保持通报 2015年6期
赵天宇, 关东海, 苏里坦, 时山良
(1.中国科学院 新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室, 新疆 乌鲁木齐 830011;
2.中国科学院大学, 北京 100049; 3.北京七兆科技有限公司, 北京 100070; 4.新疆水利水电科学
研究院 水资源规化研究所, 新疆 乌鲁木齐 830049; 5.伊犁花城勘测设计研究有限责任公司, 新疆 伊犁 835000)
环境因子对胡杨树干液流动态的影响
赵天宇1,2,3, 关东海4, 苏里坦1, 时山良5
(1.中国科学院 新疆生态与地理研究所 荒漠与绿洲生态国家重点实验室, 新疆 乌鲁木齐 830011;
2.中国科学院大学, 北京 100049; 3.北京七兆科技有限公司, 北京 100070; 4.新疆水利水电科学
研究院 水资源规化研究所, 新疆 乌鲁木齐 830049; 5.伊犁花城勘测设计研究有限责任公司, 新疆 伊犁 835000)
摘要:[目的] 分析胡杨生长期内各环境因子对树干液流速率的影响程度,为干旱区自然植被耗水量的计算提供了新的方法和科学依据。 [方法] 对塔里木河下游2012单木胡杨整个生长季内树干液流以及空气温度、风速、有效光合辐射强度、空气湿度、土壤温湿度等环境因子进行连续观测。 [结果] 胡杨在整个生长季内的液流呈单峰波动曲线,有明显的昼夜变化规律;在生长期内,不同的观测尺度下,影响胡杨液流速率的环境因子不同。回归分析结果表明,对于瞬时液流流速,相关性较大的环境因子主要是大气温度、10 cm深处的土壤温度;对于日液流量,主要影响因子为太阳净辐射;对于月液流量,主要影响因子为大气温度和太阳净辐射。[结论] 研究区内胡杨生长受大气温度和太阳净辐射影响程度较其他因素大,并且回归方程具有很高的拟合精度。
关键词:塔里木河下游; 胡杨; 液流速率; 环境因子
塔里木河流域土地资源丰富,流域总面积1.02×106km2,灌区面积1.26×106hm2,其中农田灌溉面积9.56×105hm2,人工林草灌溉面积3.06×105hm2。其森林资源按起源分为天然林和人工林两大类。近年来,塔里木河流域沙漠化十分严重,根据1959和1983年航片资料统计分析,24 a间塔里木河干流区沙漠化土地面积上升了15.6%,下游土地沙漠化发展最为严重,上升了22.05%,导致气温上升,大风、沙尘暴日数增加,植被衰败,交通道路、农田及村庄埋设,严重威胁绿洲生存和发展,合理的治理沙漠化问题亟需解决。有研究[1]指出,种植防护林可以有效的治理沙漠化,防治沙埋问题。但是,关于极端干旱条件下植物耗水的研究资料很少。因此,沙漠腹地防护林植物蒸腾耗水的研究有十分重要的意义。
胡杨(Populuseuphratica)主要分布我国西北荒漠半荒漠地区—尤其是塔里木河下游,具有耐盐碱、耐贫瘠、耐风蚀和沙埋等特点,是水土保持和荒漠化防治和不同断面荒漠河岸林主要建群物种同时也常用于土壤改良,也是塔里木沙漠公路防护林的一个重要构成树种。液流是植物的一个重要生理生态参数,可反映植物体内的水分传输状况[2-3]、植物对水分的利用特征[4-5]及其对于环境的响应[6-7],也是验证、修正蒸腾(蒸散)模型、根系吸水模型的重要参数[8],具有连续观测,持续测定等优点[9-10]。胡杨蒸腾变化规律,不仅与植物本身的生长状况,还受太阳有效光辐射、水汽压亏缺、土壤湿度、降雨、温度、风速等环境因子的影响[11],国内很多学者研究结果也表明不同树种对不同环境因子的响应不一致,如沙木蓼的液流大小与太阳辐射、空气温度、风速和相对湿度显著相关[12];空气水汽压差和气温等环境因子的变化则对梭梭的生长影响较敏感,太阳辐射则主要影响着对尾巨桉的生长[13];太阳辐射、空气湿度、空气温度以及风速等环境因子显著影响着柠条的液流速率[14-15]。对于树型较大的胡杨,如何准确的测定其蒸腾作用还没有合理的方法。郭树江[11]、乌日娜[12]等、蒋文伟[16]等、赵敏丽[17]等、王文杰[18]等都选择了具有代表性的单株样本进行树木液流的相关研究,而塔里木河下游拥有胡杨林面积约3.52×105hm2,选择具有代表性的单木胡杨,研究其树木液流动态规律对河岸胡杨林的正常生长以及合理保护种群结构具有重要价值。因此,本研究通过对塔里木河下游荒漠河岸胡杨单木树干液流的监测,结合实测空气温度、风速、有效光合辐射强度、空气湿度以及土壤温湿度等环境因子,分析在不同研究尺度下,各环境因子对胡杨液流的动态影响,探讨维系塔里木河下游天然胡杨的生长状况,为干旱内陆河流域受损生态系统的恢复重建提供科学依据。
1研究区概况
研究区位于塔里木河下游上端达西库勒胡杨林管护站附近的天然胡杨林生长带(85°32′48″E,40°59′19″N,海拔906 m),多年平均降水量为50 mm,最高日均气温达30 ℃,最低日均气温为-15 ℃,夏季日均太阳辐射在260~350 W/m2,而冬季日均太阳辐射在80~100 W/m2之间,具有很好的代表性。管护区内胡杨林平均高约12 m,幼龄胡杨、中龄胡杨和成熟胡杨的平均胸径分别为7.4和19.2,25.8 cm,分布密度为0.06棵/m2,胡杨林覆盖度为32%。土壤质地类型为粉砂壤土至砂土。地下水位在0.5~4.0 m之间,生态输水后(9月份)地下水位太高至0.5 m。林下有少量的红柳和1年生植物。
2观测要素的测定方法
2.1 观测要素
供试植物为胸径为23 cm的生长良好、无病虫害的单株胡杨,试验观测期为2012年1—12月。(1) 环境因子的测定。地表蒸发量采用微型蒸发器测定;太阳净辐射、冠层顶大气温度、湿度、降雨量、风速等气象数据采用Hobo自动气象站测定;胡杨胸径等采用卷尺测定;土壤温度及湿度采用TDR进行测定,观测时间步长为30 min。 (2) 地下水位埋深的测定。采用定位法测定,垂直于河道从河岸到沙丘布设了3个监测断面,利用水位管和钢尺水位计,配合水准测量,确定地下水位高程,通过各观测期水位管内水面高程的变化,监测地下水位的变化量。(3) 树干液流的测定。胡杨边材液流速率用TDP插针式径流计。该方法的数据采集具有准确和稳定的特点,且可以连续不间断地读取数据,因而数据具有系统性。该测定系统由一对长20 mm含通镍合金热电偶的圆柱形热消散探针(Granier径向型探针)组成,安装时将探针上下相隔10~15 cm插入树木的边材中,上方探针处缠绕电阻丝,供以直流电进行加热,下方的探针不加热,保持与周围边材组织的温度相同,两探针的温差变化反映树木的液流密度(sap flux density),Granier建立了反映两者关系的标准方程:
(1)
式中:Js——树干液流速率〔g/(cm2·min)〕;Tm——分析日24 h内上下探针的最大温差值(℃);T——某时刻瞬时温差值(℃),即当时测定的温差值。
2.2 测定方法和步骤
(1) 选定样木同时编号纪录; (2) 在东西2个方位胸高处刮去粗树皮用钻孔模板贴在去皮部位用钻头钻孔直至探针可以方便插入为止; (3) 将TDP30探针小心插入孔中含有红色导线的探针在上,2线探针在下,探针外围裹一圈胶泥; (4) 将探针用泡沫块夹住后胶带固定之后在外面包裹绝缘防辐射材料最后用胶带密封以防止雨水进入; (5) 将TDP馈线与数据采集器连接接通电源设定并输入仪器工作参数树号日期时间等。数据采集间隔期为30 min。假定树干边材中液流密度处处相等[12],则单木液流速率J(g/min)计算公式为:
J=Js·SA·ρ
(2)
式中:ρ——水的密度(g/cm2);SA——边裁面积(cm2)。
单木日液流量Jd(L/d)计算公式为:
(3)
式中:ΔT——采样时间间隔(min),在本研究中为30 min;Jsi——第i时刻的单木液流速率(g/min)。
单木月液流量Jm(L)计算公式为:
(4)
式中:n——每月的天数;Jdi——该月第j天的单木日液流量(L/d)。
试验数据均采用Excel和SPSS软件对液流与各环境影响因子分别进行制图分析,曲线拟合和相关性分析。
3结果与分析
3.1 胡杨液流的连日变化及其与环境因子的相关分析
胡杨液流速率有明显的昼夜节律性,白天液流速率大,晚上液流速率小。图1为一株胸径为27 cm的单木胡杨在9月4—6日胡杨液流速率与太阳净辐射量的日变化。由图1可以看出,胡杨的液流日变化由于受到环境因子的影响而呈单峰波动曲线;胡杨的日平均液流速率为1.26 g/min,树干液流速率在7:30呈明显的上升趋势,并在10:30—12:00出现液流的高峰值;随后有短暂的液流急骤减少现象,说明植物为了适应严酷的沙漠环境,保持体内水分而出现特有的“午休”现象[19]。在15:30左右时,液流开始明显下降,直到22:00—1:00出现最小值,可以发现夜间的液流速率极小,这是由于在干旱条件下,白天较高的太阳净辐射和大气温度导致植物损失大量的水分,因此在夜间仍需要一定的液流以补充水分,同时空气干燥也可能是植物水分散失的一个重要原因[20]。
3.1.1液流速率与太阳净辐射量的关系经过对连续3 d的144个数据进行相关系数分析得出,胡杨液流速率与太阳净辐射量呈显著正相关(R2=0.488)。从图1可以看出,太阳辐射上升2 h后,液流速率迅速增大并达到峰值,之后开始下降并趋于平缓。随着太阳净辐射量达到最大值,速率会出现小的波动。随着太阳净辐射强度的增大,气温升高,树木生理活动增强,空气导度增大,叶片蒸腾增强,进而使液流速率增大。当叶片表面温度过高(超过32 ℃)时,气孔大量关闭[21],从而导致液流速率下降,因此液流速率和太阳辐射的峰值出现的时间并不完全一致。夜间太阳辐射为0,叶片气孔关闭,树木液流速率停止,但这段时间依然存在较小的树干液流,一般认为是由于白天的蒸腾耗水较多,树干出现水分亏缺,晚上通过根系吸水补充,从而出现树干液流。
3.1.2液流速率与大气温度的关系随着大气温度的升高,胡杨液流速率呈逐渐增大的趋势,二者呈显著正相关(R2=0.303)。当大气温度开始升高时,液流速率明显增大,然后开始下降并趋于平缓(图1);在空气温度达到最大值之前出现一个小的波峰,之后平缓下降,即开始出现“午休”现象,此时树干液流速率呈小的波动变化。
3.1.3液流速率与相对湿度的关系空气相对湿度与液流速率呈负相关(R2=0.346),其变化趋势是随着湿度的减小而逐渐上升(图1)。当空气湿度开始减小时,树干液流速率开始迅速增大并达到最大值,之后迅速下降趋于平缓;在空气相对湿度接近最小值时出现一个小的波峰。大气温度和相对湿度具有协同效应。当环境中的空气相对湿度增加时,将导致液流速率下降,甚至会出现为零的情况。
3.1.4液流速率与风速的关系风速与液流速率呈显著正相关(R2=0.296),其作用机理比较复杂。微风可以将气孔外围相对湿度较高的空气吹走,使蒸气扩散层变薄或消失,外部营力减小,从而促进蒸腾;而风速太大则会导致气孔开度降低,甚至导致气孔关闭,液流速率变小,从而抑制液流速率(图1)。

图1 胡杨液流速率与各影响因子之间的日变化(2012年9月4-6日)
3.1.5液流速率与土壤温度及土壤含水量的关系土壤温度和土壤水分含量呈负相关,但相关性极小,R2分别为0.003 0,0.000 5,主要原因是由于在连续3 d内,土壤温度和土壤水分含量变化率分别为-0.001 3和-0.000 1,因此对胡杨液流速率的影响可以忽略不计。
3.2 单木日液流量的月变化
胡杨单木月液流量表现为1月份最小,只有131.19 L,7月最大,达765.94 L。冬季(2012年1,2,12月)、春季(2012年3—5月)、夏季(2012年6—8月)、秋季(2012年9—11月)胡杨单木季液流量分别为464.95,739.72,1 662.04,995.80 L(图2)。
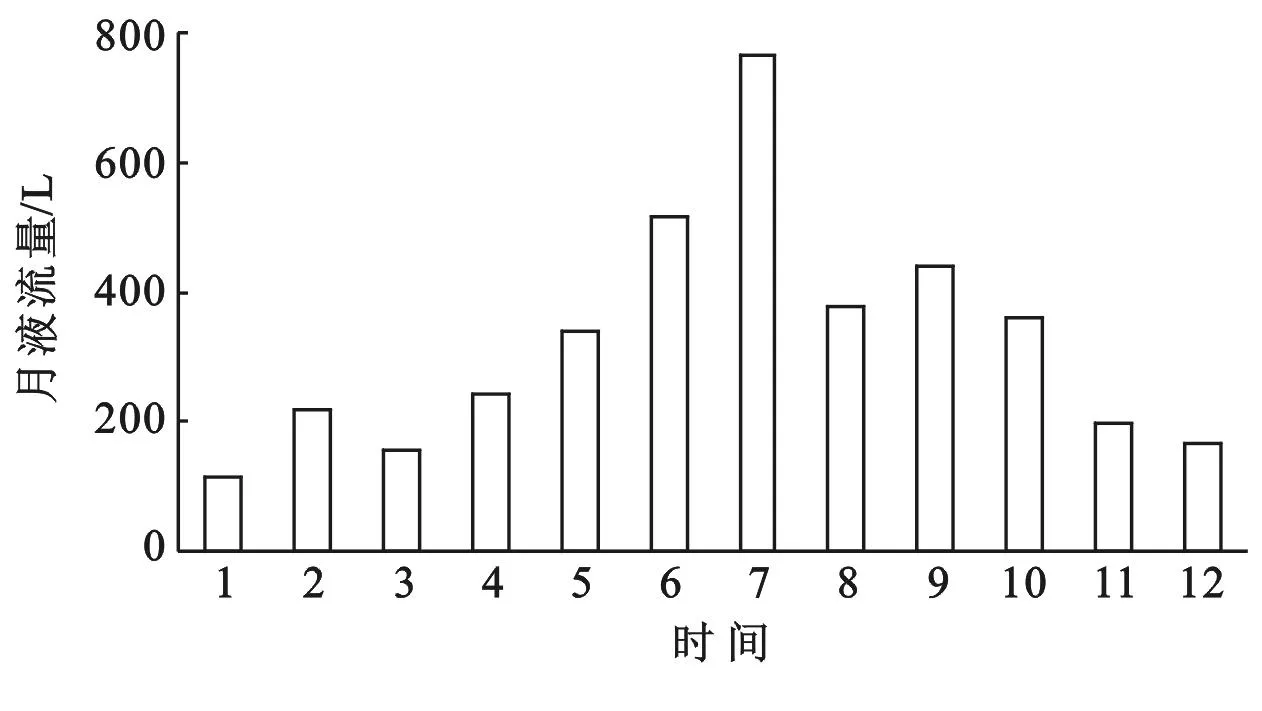
图2 胡杨2012年1-12月液流量变化
3.3 单木日液流量的年变化
由图3可知,2012年胡杨在冬季即1,2,12月基本不生长,日液流量平均值为(5.37±0.3) L/d,2月13日日液流量平均值为16.12 L/d,为最大;12月18日日液流量平均值为0.98 L/d,为最小;该时期平均温度-7.88 ℃,而导致这一时期液流量小的主要限制因子可能是较低的温度;春季即3月1日至5月31日,日液流量在(8.04±0.45) L/d变化,5月10日最大(17.85 L/d),3月8日最小(2.7 L/d),该时期太阳平均净辐射强度最高(176 W/m2)、大气平均温度也开始升高(14.7 ℃)同时土壤水分含量日际变幅增大;夏季即6月1日至8月31日,日液流量呈波动上升趋势,平均值为(18.07±0.8) L/d,7月14日最大(53.16 L/d),8月24日最小(6.7 L/d),土壤水分含量开始降低,大气湿度则相对较高,温度为该时期为一年中最高(28.8 ℃)、太阳辐射强度为次高(155.05 W/m2);秋季即9月1日至11月30日,日液流量平均值为(10.94±0.6) L/d,9月16日最大(20.44 L/d),11月6日最小(3.01 L/d),该时期大气温度开始慢慢降低,太阳辐射也达到了全年最低(40.61 W/m2),导致液流速率下降的主要原因是土壤水分含量的持续下降。
3.4 胡杨液流与环境因子的关系
3.4.1胡杨液流速率与环境因子的关系以胡杨单木液流速率为因变量,以太阳净辐射日总量、及气温、相对湿度、风速、土壤温度、土壤水分含量等日均值为因变量,通过逐步线性回归分析,得到回归方程为:
Js=26.585+0.032Rs+12.667T-
0.563Rh-9.588T10+1.236CS
(F=15.126,Df=8.30,p<0.01)
式中:Rs——太阳净辐射(W/m2);T——空气温度(℃);Rh——空气相对湿度(%);T10——10 cm深处的土壤温度;v——风速;CS——土壤水分含量。
回归分析表明,胡杨液流速率与大气温湿度、土壤水分含量、太阳净辐射和10 cm深处的土壤温度均相关,与风速相关性不显著。其中,液流速率与大气温度及10 cm深处的土壤温度最为显著,相关性分别为0.868,0.831。

图3 胡杨2012年1-12月日液流量变化
3.4.2胡杨日液流量与环境因子的关系以胡杨单木日液流量为因变量,以太阳净辐射日总量、及其他环境因子的日均值为因变量,通过逐步线性回归分析,得到回归方程为:
Jd=104.169+0.004Rs+12.931T-
0.865Rh-4.568Ts
(F=3.140,Df=7,358,p<0.01)
回归分析表明,日液流量与太阳净辐射、空气温湿度以及10 cm深处的土壤温度相关性较大,与风速及土壤水分含量相关性不显著。其中,太阳净辐射与树干液流相关性最为显著,相关系数达到0.925。
3.4.3胡杨月液流量与环境因子的关系以胡杨单木月液流量为因变量,以太阳净辐射月总量、及其他环境因子的日均值为因变量,通过逐步线性回归分析,得到回归方程为:
Jm=14196.503+0.004Rs+12.931T-
5.031v+2.224Cs
(F=3.953,Df=6.5,p<0.01)
回归分析表明,单木月液流量与太阳净辐射、空气温度、土壤水分含量以及风速相关性较大,与10 cm深处的土壤温度和相对湿度相关性不显著。其中,日液流量与太阳净辐射和空气温度的相关性最为显著,相关系数为0.896,0.838。
4结 论
(1) 植物蒸腾过程变化规律明显。在干旱的沙漠地区,胡杨树干液流日变化呈单峰曲线,有明显的昼夜变化特征,液流受环境因子变化有所波动;在10:30—12:00出现液流的高峰值;在15:30左右时,液流开始明显下降,直到22:00—1:00出现最小值,对于阴雨天,蒸腾过程则无明显的变化特征[16],因此本文不作分析。
(2) 胡杨树干液流的变化规律不仅受到不仅与植物本身的生长状况,还受到各环境因子的影响。其中,影响着植物的耗水量大小的主要是太阳净辐射以及由其引起的空气温湿度变化。本研究结果发现,在研究观测期间(2012年1—12月)日照时间最长,太阳净辐射季总量最大(479 177.5 W/m2)的是夏季,其次为春季(440 088.2 W/m2)、秋季(419 523.8 W/m2),冬季最低(279 177.5 W/m2),因此在夏季树干液流速率高于其他季节。
(3) 胡杨在各个季节生长速率也有很大的差别,因此液流密度具有很大的差异,在冬春夏秋季,胡杨日液流量分别为(5.37±0.3) L/d,(8.04±0.45) L/d,(18.07±0.8) L/d,(10.94±0.6) L/d,而胡杨季液流量分别为468.95,739.72,1 662.04,995.80 L。
(4) 回归分析表明,环境因子对胡杨树干液流的影响程度随着研究尺度的不同而不同。对于连续3 d的液流速率,相关性较大的主要是太阳净辐射量,相对湿度以及温度,相关系数分别为0.488,0.346,0.303;对于瞬时液流流速,相关性较大的主要是温度、10 cm深处的土壤温度,相关系数分别为0.868,0.831;对于日液流量,相关性较大的因子主要是太阳净辐射,相关系数达到0.925;对于月液流量,相关性较大的因子主要是空气温度和太阳净辐射,相关系数分别为0.896,0.838。
[参考文献]
[1]许浩,张希明,闫海龙,等.塔克拉玛干沙漠腹地多枝柽柳茎干液流及耗水量[J].应用生态学报,2007,18(4):735-741.
[2]Nadezhdina N. Sap flow index as an indicator of plant water status[J]. Tree Physiology, 1999,19(13): 885-891.
[3]Cohen M, Goldhamer D A, Fereres E, et al. Assessment of peach tree responses to irrigation water deficits by continuous monitoring of trunk diameter changes[J]. The Journal of Horticultural Science and Biotechnology, 2001,76(1): 55-60.
[4]Andrade J L,Meinzer F C, Goldstein G, et al. Regulation of water flux through trunks,branches, and leaves in trees of a lowland tropical forest[J]. Oecologia, 1998,115(4):463-471.
[5]Wullschleger S D, Meinzer F C, Vertessy R A. A review of whole-plant water use studies in trees[J]. Tree Physiology, 1998,18(8/9): 499-512.
[6]Kellomki S, Wang K Y. Sap flow in Scots pines growing under conditions of year-round carbon dioxide enrichment and temperature elevation[J]. Plant Cell and Environment, 1998,21(10): 969-981.
[7]Wullschleger S D, Norby R J. Sap velocity and canopy transpiration in a sweetgum stand exposed to free-air CO2enrichment(FACE)[J]. New Phytologist, 2001, 150(2): 489-498.
[8]Gong Daozhi, Kang Shaozhong, Lu Zhang, et al. A two-dimensional model of root water uptake for single apple trees and its verification with sap flow and soil water content measurements[J]. Agricultural Water Management, 2006,83(1): 119-129.
[9]Smith D M, Allen S J. Measurement of sap flow in plant stems[J]. Journal of Experimental Botany, 1996, 47(305): 1833-1844.
[10]Gazal R M, Scott R L, Goodrich D C, et al. Controls on transpiration in a semiarid riparian cotton wood forest[J]. Agricultural and Forest Meteorology, 2006, 137(1): 56-67.
[11]郭树江,徐先英,杨自辉,等.干旱荒漠区沙冬青茎干液流变化特征及其与气象因子的关系[J].西北植物学报,2011,31(5):1003-1010.
[12]乌日娜,丁国栋,吴斌,等.沙木蓼蒸腾耗水特性及环境影响因子研究[J].水土保持通报,2012,32(3):68-72.
[13]王文,朱烨,诸葛绪霞,等.尾巨桉树干液流特性及其影响因子分析[J].水土保持通报,2013,33(3):159-164.
[14]黄磊,张志山,周小琨,等.沙漠人工植被区柠条树干液流变化及影响因子分析[J].中国沙漠,2011(2):415-419.
[15]苏里坦,玉米提,宋郁东.基于改进BP神经网络的干旱区李伟腾发量预测模型[J].干旱区地理,2011,34(4):551-557.
[16]蒋文伟,郭运雪,杨淑贞,等.天目山柳杉树干液流动态及其与环境因子的关系[J].江西农业大学学报,2011,35(5):899-905.
[17]赵敏丽,刘普幸,朱小娟,等.近51年来额济纳旗绿洲胡杨年生长期对气候变化的响应[J].水土保持通报,2012,32(5):205-209.
[18]王文杰,孙伟,邱岭,等.不同时间尺度下兴安落叶松树干液流密度与环境因子的关系[J].林业科学,2012,48(1):77-85.
[19]郭跃,丁国栋,吴斌,等.毛乌素沙地沙木蓼茎干液流规律研究[J].水土保持通报,2010,30(5):22-26.
[20]曹晓明,陈曦,王卷乐,等.古尔班通古特沙漠南缘非灌溉条件下梭梭(Haloxylonammodendron)蒸腾耗水特征[J].干旱区地理,2013,36(2):292-302.
[21]司建华,常宗强,苏永红,等.胡杨叶片气孔导度特征及其对环境因子的响应[J].西北植物学报,2008,28(1):125-130.
Effects of Environmental Factors on Trunk Sap Flow ofPopulusEuphratica
ZHAO Tianyu1,2,3, GUAN Donghai4, SU Litan1, SHI Shanliang5
(1.StateKeyLaboratoryofDesertandOasisEcology,XinjiangInstituteofEcologyandGeography,CAS,Urumqi,Xinjiang830011,China; 2.GraduateUniversity,ChineseAcademyofSciences,Beijing100049,China; 3.BeijingSevenTrillionTechnologyCoLtd,Beijang100070,China; 4.InstituteofWaterResourcesofXinjiangWaterResourcesandHydropowerResearch,Urumqi,Xinjiang830049,China; 5.LimitedLiabilityCompanyofYiliHuachengSurveyandDesignResearch,Yili,Xinjiang835000,China)
Abstract:[Objective] The influences of environmental factors on sap flow rate of Populus euphratica were analyzed to provid method data base for the calculation of water consumption by natural vegetation in arid area. [Methods] The trunk sap flow of Populous euphratica at the lower reaches of Tarim River was measured continuously during the whole growth season in 2012 and its responses to environmental factors were analyzed. [Results] The sap flow of Populus euphratica presented an unimodal curve, with obvious day and night variation rhythm. During the growth season, the relations between sap flow rate and environmental factors depended upon the temporal scales. Regress analysis showed that when observed at less hour scale, sap flow were associated closely with air temperature and soil temperature at depth of 10 cm. However, on daily scale, solar radiation was the major influencing factors; and on monthly scale solar radiation and air temperature were the major influencing factors. [Conclusion] The major factors assocated with sap flow of Populus euphratica are air temperature and solar net radiation, and the regressions of them with the sap flow rate were fitted wells.
Keywords:the lower reaches of Tarim River; Populus euphratica; sap flow rate; environmental factor
文献标识码:A
文章编号:1000-288X(2015)06-0015-06
中图分类号:Q948.1
通信作者:苏里坦(1972—),男(哈萨克族),新疆自治区霍城县人,博士,副研究员,主要从事地表水文过程研究。E-mail:sulitan@ms.xjb.ac.cn。
收稿日期:2014-09-23修回日期:2014-10-01
资助项目:国家自然科学基金项目“塔里木河下游断面尺度水循环过程模拟研究”(U1403182),“地下变水位条件下塔里木河下游河岸胡杨林耗水模型研究”(41071026)
第一作者:赵天宇(1990—),男(汉族),安徽省亳州市人,硕士研究生,研究方向为地表水文过程。E-mail:zhaoskyu@foxmail.com。
猜你喜欢
金桥(2021年11期)2021-11-20
北广人物(2020年47期)2020-12-09
音乐教育与创作(2020年11期)2020-11-17
创新作文(1-2年级)(2018年6期)2018-01-22
甘肃林业(2016年4期)2016-11-07
中国市场(2016年36期)2016-10-19
天津农业科学(2015年12期)2015-12-03
河北渔业(2015年10期)2015-10-15
塔里木大学学报(2014年3期)2014-03-11
