
城市化对南亚热带常绿阔叶林土壤生物群落结构的影响
2015-03-12 01:29符方艳陆宏芳
生态环境学报 2015年6期
符方艳,陆宏芳
1. 中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049
城市化对南亚热带常绿阔叶林土壤生物群落结构的影响
符方艳1, 2,陆宏芳1*
1. 中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049
摘要:以广州市城郊梯度上存留的南亚热带常绿阔叶林为对象,量化分析城市化对南亚热带常绿阔叶林土壤微生物和土壤动物群落结构的影响,运用生态㶲(Eco-exergy)理论方法,量化揭示土壤生物群落生物热力学结构对城市化的综合响应规律。结果显示,城市化对南亚热带常绿阔叶林土壤动物和微生物群落结构影响的方向、速率和机制各不相同,可能的影响主要有土壤含水率、铵态氮、有机质、硝态氮以及草本层生物量等。城市化会降低南亚热带常绿阔叶林土壤微生物群落生物量和生态㶲,并加剧干、湿季差异;而适度的城市化干扰可增加南亚热带常绿阔叶林土壤动物类群数和生态㶲,并减少干、湿季的差异。综合而言,广州近郊和城区残留的南亚热带常绿阔叶林土壤生物群落生态㶲与结构㶲值均高于远郊区的水平(2.39~2.99倍);南亚热带常绿阔叶土壤生物群落生态㶲主要由大型土壤动物贡献(>64.41%),而中小型土壤动物则是结构㶲的主要贡献者(>50.00%);不考虑生命周期,土壤微生物对于土壤生物群落生态㶲与结构㶲值的做贡献较小。生态㶲理论方法可以综合度量土壤生物群落的自组织程度,但在具体计量中受到权重因子的有限性制约。
关键词:土壤微生物;土壤动物;群落结构;生态㶲;结构㶲
引用格式:符方艳,陆宏芳. 城市化对南亚热带常绿阔叶林土壤生物群落结构的影响[J]. 生态环境学报, 2015, 24(6): 938-946. FU Fangyan, LU Hongfang. Effects of Urbanization on Soil Community Structure under Subtropical Evergreen Broad-leaved Forests [J]. Ecology and Environmental Sciences, 2015, 24(6): 938-946.
南亚热带常绿阔叶林具有生物多样性高、生态系统服务功能强和对环境变化敏感性高的特点,在调解生态平衡、改善自然环境和保障区域可持续发展方面发挥着重要的作用(彭少麟,2003;Zhou等,2006)。华南地区是全球南亚热带常绿阔叶林重要分布区,但同时也是我国社会经济发展的重要先导区域,人口密度高、经济活跃、城市化进程快。量化揭示城市化对南亚热带常绿阔叶林结构与功能的影响规律,是科学制定区域森林保育与恢复策略、保障区域可持续发展的前提,具重要现实意义(Huang等,2013)。土壤生物作为森林地下生态系统最活跃的部分,对环境波动敏感性强、反应迅速,是森林生态系统重要的结构与功能单元和研究森林对干扰响应的理想生物类群(王绍军和阮宏华,2008;傅声雷,2007)。目前,关于城市化对森林生态系统中土壤微生物群落特征影响的研究较多(Zhao和Guo,2010;赵哲霞和郭恢财,2010;陈帅等,2012),而对土壤动物的研究较少(秦钟等,2009;杨冬青和高峻,2002),联立两者的土壤生物综合研究则更少(李俊生等,2003),城市化对南亚热带常绿阔叶林土壤生物群落结构的综合影响规律尚未被揭示。
生态系统是开放的热耗散系统,其各种结构与功能项复杂的非线性发展动态背后可能遵循统一的生物热力学自组织规律,如最大功率原则(Lotka,1992a,1992b)。生态㶲(Eco-exergy)理论方法,可用以度量生态系统的生物热力学组织程度和健康水平(Jørgensen,2006)。自提出以来,主要被应用于湿地和水生生态系健康评价的研究(Libralato等,2006;Jørgensen,2007;Austoni等,2007;Xu等,2011;Molozzi等,2013);森林生态系统的相关研究尚处于起步阶段,现有研究主要集中在植被群落结构部分(Lu等,2011;李浩等,2012;黄柳菁,2012),土壤生物群落的生态㶲研究未见报道。
本研究分析了广州市城区-近郊区-远郊区梯度上,南亚热带常绿阔叶林林下土壤微生物和土壤动物群落结构,并引入生态㶲理论方法,综合量化各土壤生物群落的生物热力学结构水平,分析揭示城市化对南亚热带森林生态系统土壤生物群落结构的影响规律;联立各样地土壤理化性状,探讨可能的影响机理,为保育和恢复南亚热带城市森林生态系统提供科学依据。

图1 广州市城郊梯度上研究样地Fig. 1 The location of studied areas situated along an urban-rural gradient in Guangzhou
1 研究区概况
广东省广州市位于中国华南沿海,属于海洋性亚热带典型季风气候,全年有明显的干湿季交替,地带性植被和土壤分别是南亚热带常绿阔叶林和赤红壤。本研究以广州市城郊梯度上残存的天然恢复50年或以上的南亚热带常绿阔叶林为研究对象(图1),根据研究样地距离城市中心距离以及周围土地利用类型和人口密度划分研究样地,构建了城区蒲岗、近郊区萝岗以及远郊区大岭山3个森林退化样地(Huang等,2013)。
(a)城区蒲岗样地,PG(E113°21′,N23°11′)
位于广州市东北部的华南植物园内,是由“风水林”演化而成的亚热带次生常绿阔叶林,植物群落结构复杂,种类丰富,主要植被有木荷Schima superba、牛矢果Osmanthus matsumuranus Hayata、降真香Dalbergia odorifera等,土壤类型为赤红壤。蒲岗周围交通发达,居民房屋较多。
(b)近郊区萝岗样地,LG(E113°18′,N23°06′)
位于广州市东南部萝岗区长龙村,是目前广州市保存完好的、由“风水林”发育而成的南亚热带次生常绿阔叶林。萝岗区长龙村风水林周围主要为乡镇居民点、园地和耕地,邻近高速公路。
(c)远郊区大岭山样地,DL(E113°48′,N23°37′)
位于东北部距离广州市中心约75 km的从化区,属于大岭山林场和石门国家森林自然保护区,主要植被为南亚热带天然次生林常绿阔叶林。大岭山作为广州市域内的远郊区,区域经济发展以生态林业为主,受人为干扰较少。
2 研究方法
2.1土壤生物群落采样分析
野外采样时间:分别于2013年12月份和2014 年6月份进行采样,这两个时间点分别代表了我国南亚热带地区的干季和湿季。每个研究样地中,设立2条5 m×25 m的平行样带,每条样带间隔10 m。每条样带划分为5个5 m×5 m的样方,在每个样方内在设立一个1 m×1 m的小样方。
2.1.1土壤微生物群落的采样与分析
在样带中随机选取6个不相邻的1 m×1 m小样方,每个样方内按对角线法取5个点。5个点的土样混合为一个土壤样品,用直径3 cm的土钻进行采样,采土深度0~15 cm。运用PLFA(Phospholipid Fatty Acid)方法测定。
2.1.2土壤动物群落的采样与分析
在样带中随机选取6个不相邻的1 m×1 m小样方,每个样方采用直线法取3个混合为一个样品。干漏斗用直径5 cm采集土壤样品,采土深度0~15 cm。采用手拣法收集大型土壤动物,30 cm×30 cm,深度15 cm,保存在装有75%酒精的小瓶中,带回室内测定。
采用手拣法、干漏斗法(Tullgren法)分离后,于光学显微镜下,依据《中国土壤动物检索图鉴》进行分类鉴定,统计种类、数量,采用真空烘箱烘至恒质量,测定其生物量。
2.2土壤理化性状的测定
采样时间和方法与土壤微生物的采样相同。环刀法测定土壤容重、采用烘干法测定土壤含水量、酸度计法测定pH值、重铬酸钾氧化-外加热法测定有机碳、过硫酸钾氧化-紫外分光光度计法测定全氮(TN)、氯化钾浸提-靓酚蓝比色法测定土壤铵态氮、镀铜镉还原-重氮化偶合比色法测定硝态氮、氢氧化钠-钼锑抗比色法测定全磷(TP)、火焰光度计法测定全钾(TK)。
2.3数据处理
2.3.1土壤动物多样性
不同植被类型中土壤动物群落结构和多样性均有不同的表现,反映了土壤动物对不同生境类型的响应,在此以类群(目)丰富度为指标,选择DG多样性指数(DG)、Simpson优势度指数(C)、Pielou均匀度指数(J)和Margalef丰富度指数(D),反映土壤动物结构多样性特征。
土壤动物的多样性采用密度-类群指数,即DG指数(廖崇惠等,1997)表示:
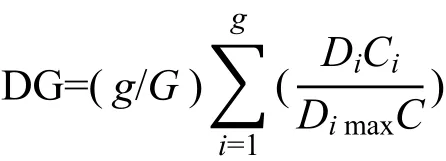
式中Di为第i类群个体数;Dimax为各群落中第i类群的最大个体数;g为群落中的类群数;G为各群落所包含的总类群数,Ci/C为C个群落中第i个类群出现的比率,以显示该类群在群落中的重要性。
Simpson优势度指数:C=∑(ni/N)2
Pielou均匀性指数:J=(-∑PilnPi)/lnS
Margalef丰富度指数:D=(S-1)/lnN
式中S为样方的土壤动物种类总和;ni为种i的个体数;N为样方所有物种的个体数之和;Pi为种i的个体数占所有种的个体数的比率。
2.3.2生态㶲(Eco-exergy)与结构㶲(Specific Eco-exergy)
生态㶲(Ex)理论方法由丹麦学者Jørgensen 于20世纪70年代提出,被定义为生态系统生物组分相对环境的自由能(Jørgensen和Mejer,1977,1979),计算公式如下:
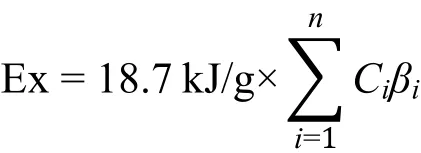
其中,18.7 kJ·g-1为分解1 g碎屑物所含的平均㶲值,Ci为i种的生物量(g·m-2),βi为物种相对于碎屑物的权重转换因子(Jørgensen等,2002)。
本研究的中的β值来源于Jørgensen等(2005)和Jørgensen(2007)的研究,引用数据尽可能在目的水平上估算,若不能精确到目则以同纲动物的β值计算,不能精确到纲则以同门动物的β值计算。
结构㶲(Specific Eco-exergy)即单位生物量的生态㶲值,指示系统内生物遗传信息量水平,与生物量共同决定系统生态㶲水平,分别表征系统生物热力学结构发展的两条主要路径,即生物量和遗传信息量的变化。具体计算公式如下(符号意义同前文生态㶲公式):
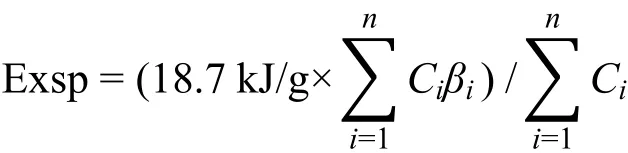
2.3.3统计分析
采用SPSS16.0 Software进行单因素方差分析,运用Canoco for windows 4.5软件的冗余分析(Canonical Redundancy Analysis,RDA)对各类群土壤生物及其生态㶲值以生境条件为背景进行相关性排序。
3 结果与分析
3.1土壤微生物
测定结果显示,在干季,所有微生物类群生物量皆呈现同一规律,即远郊区>城区>近郊区;而在湿季,则是远郊区>近郊区>城区。微生物群落中,细菌的生物量最高,真菌与放线菌次之,原生生物最低;说明细菌是常绿阔叶林退化样地土壤微生物群落中的绝对优势种群(图2)。
单因素方差分析表明,远郊区林下土壤微生物总生物量及所有类群在干季均与其它两个样地存在极显著性差异,而近郊区与城区间则无显著差异。与干季规律不同,湿季中,城区林下土壤微生物总生物量及细菌、放线菌生物量较低,与近郊与远郊区分别存在极显著性差异,而近郊与远郊区差异只在真菌、原生生物生物量上存在显著差异,其他类群差异不显著。
干湿季对比,近郊区与城区林下土壤微生物生物量表现为湿季显著高于干季;而远郊区林下土壤微生物各类群以及总微生物生物量则无显著的干、湿季差异。
3.2土壤动物
干季,共采集到土壤动物5795只,隶属2门,共有27个类群,其中远郊区、近郊区和城区样地捕获土壤动物类群数分别为22、24、22个(表1),各样地之间类群数目相近。在个体数量上,近郊区>远郊区>城区;中小型土壤动物占各样地土壤动物总数的96.17%~99.08%;以节肢动物门的真螨目、寄螨目、弹尾目、膜翅目及双翅目幼虫为优势类群,后孔寡毛目亦是近郊区和城区样地土壤动物个体数量的优势类群。在生物量上,近郊区>城区>远郊区,生物量的贡献主要来自大型土壤动物类群,占总生物量的78.79%~85.20%,其中,节肢动物门后孔寡毛目为各样地生物量上的优势类群,占总生物量的55.18%~74.56%。
湿季,取样采集到土壤动物1256只,隶属2门,共有25个类群,其中远郊区、近郊区和城区样地捕获土壤动物类群数分别为12、22、16个(表1),远郊区的土壤动物类群数最少,城区次之,近郊区最多。在个体数量上,近郊区>城区>远郊区,中小型土壤动物占总个体数的66.39%~91.42%,其中,以节肢动物门的真螨目、寄螨目、弹尾目、后孔寡毛目、膜翅目为优势类群。在生物量上城区>近郊区>远郊区,总生物量仍是以大型土壤动物类群为主,占总生物量的93.03%~97.90%,而大型动物中以环节动物门后孔寡毛目的生物量为主,各总生物量的86.27%~90.43%。
干季取样采集捕获的土壤动物总数远比湿季多,约为湿季总数的5倍。造成此差异的原因主要是中小型动物真螨目、寄螨目、弹尾目3个类群的数量在干季远多于湿季;然而,湿季大型动物在数量上则比干季要多。湿季总生物量比干季高,其中城区样地总生物量的干湿季差异最为显著。干湿两季,土壤动物生物量总量最小的都是远郊区样地。大型土壤动物生物量比重最大,说明它们在森林生态系统的物质循环和能量转换中起着重要的作用。
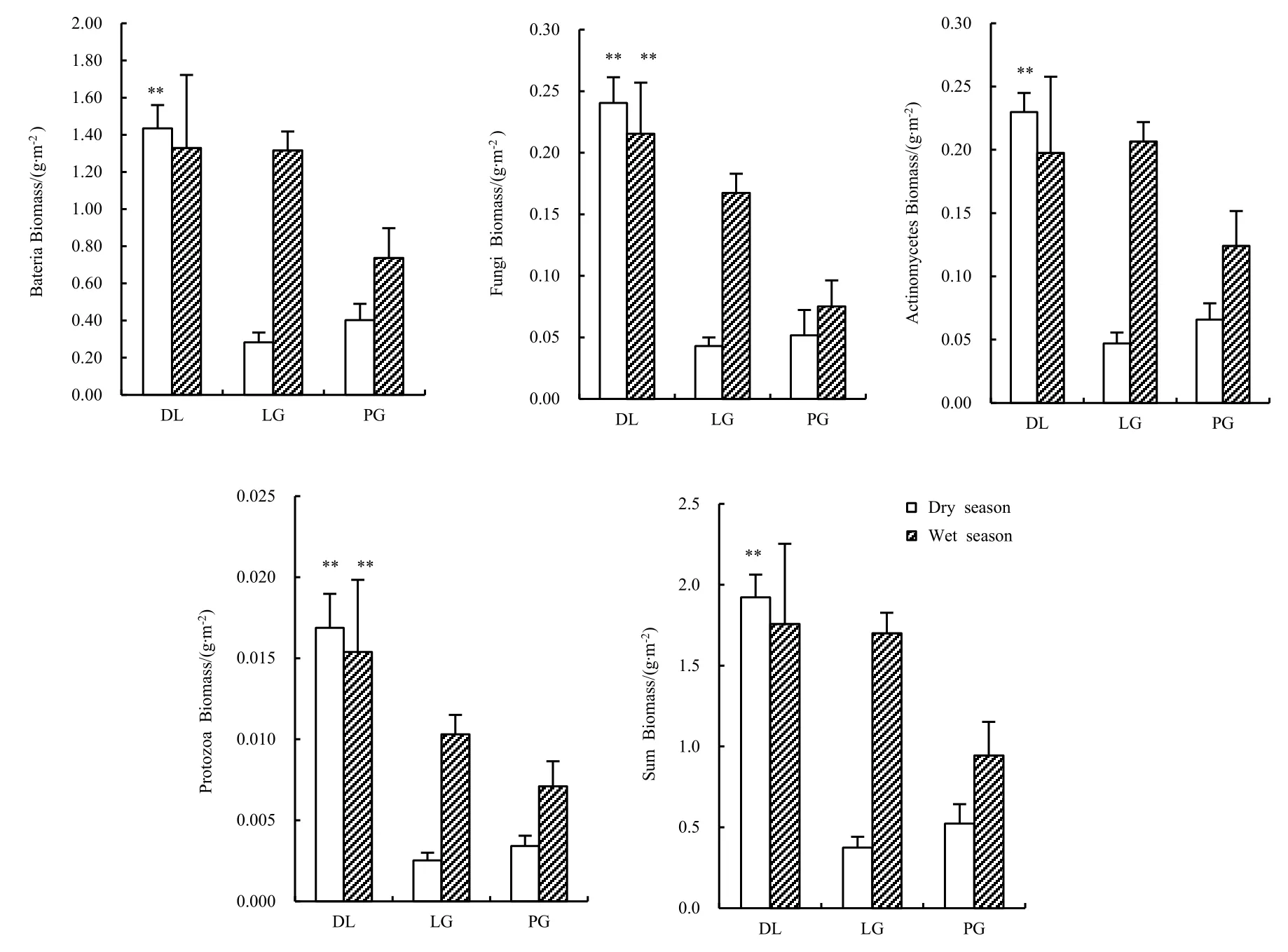
图2 干湿两季各类群微生物生物量Fig. 2 Biomass of different group microbe at each sampling site
干季,各样地间土壤动物各多样性指数均无显著差异。湿季,远郊区样地的DG指数和Pielou均匀度指数显著高于其他两个样地,而近郊区样地土壤动物群落的Simpson优势度指数显著高于远郊区和城区样地(表2)。
3.3土壤生物群落生态㶲和结构㶲
干季以近郊区林下最高,湿季以城区林下最高,但近郊区与城区总生态㶲值在干、湿季均无显著差异。远郊区样地生态㶲值在干、湿季均显著低于近郊区和城区样地水平,表明后两个森林土壤生物群落的生物热力学组织程度更高,一旦遭到破坏,恢复所需能量也更多。各样地湿季的生态㶲总值均高于干季,表明土壤生物群落受气候的影响较大,湿季土壤生物生命力旺盛,系统结构更稳定。
后孔寡毛目的生态㶲对各样地土壤生物群落总生态㶲的贡献率最大,马陆类的生态㶲值也占有一定比重,微生物仅在远郊区样地对土壤生物群落总生态㶲值具较大贡献(大于1%)。
干湿季城郊梯度上的结构㶲均以城区林下最高,近郊区林下次之,远郊区林下最小。表明单位生物量上城区林下生态系统复杂程度高。近郊区与城区林下系统结构㶲季节变化表现为湿季>干季,而远郊区林下表现为干季>湿季。不同体型土壤生物对系统结构㶲贡献程度不一,中小型土壤生物在群落结构㶲所占比例均大于50%(图3B),其次为大型土壤动物,微生物所占比例最小。
各生境土壤生物群落的生态㶲主要由大型动物决定(大于64.41%),总生态㶲的分配趋势与大型动物分配趋势大致相同,而中小型生物在群落结构㶲所占比例最大(表3),表明大型动物与中小型动物在南亚热带森林地下生态系统结构稳定性和复杂性中发挥较大作用。
3.4生态㶲与土壤理化性状因子的RDA分析
各个样地的土壤理化性质如表4所示。单因素方差分析结果表明,远郊区林下土壤干湿季的铵态氮、pH、TK以及湿季的有机质含量显著高于近郊区与城区,而城区和近郊区林下大部分土壤的理化性状指标间无显著差异。干季,远郊区林下土壤的含水率、有机质、硝态氮的含量均最低,但铵态氮含量最高;近郊区土壤含水率最高;城区土壤的有机质、硝态氮含量最高。湿季,远郊区林下土壤的含水率、有机质、铵态氮、硝态氮的含量均最高,近郊区的有机质、铵态氮的含量最低,城区土壤的含水率与硝态氮最低。

表2 土壤动物群落多样性指数Table 2 Diversities of soil fauna at different sites on the urban-rural gradient

图3 各植被类型的不同体型土壤生物的生态㶲与结构㶲Fig. 3 Eco-exergy and Specific eco-exergy of different sizes’ soil organisms at the three sites on the urban-rural gradient
土壤生物生态㶲和9个土壤理化因子以及地上部分乔、灌、草层植物生物量(黄柳菁,2012)共12个因子之间相关关系的冗余分析(RDA)结果(表5)显示,第一排序轴主要反映土壤含水率、铵态氮、有机质,第二排序轴主要反映硝态氮、草本层生物量。其中第一和第二轴分别解释了29.60%和26.00%土壤生物生态㶲和生境因子关系的变异量,足够解释土壤生物群落生态㶲和土壤因子、地上植物的相关性。
城郊梯度森林生态系统地下生物类群中,后孔寡毛目的生态㶲受土壤含水率、有机质、铵态氮的影响较大,呈现正相关。中小型的鞘翅目幼虫、马陆类受硝态氮、TN以及有机质的影响较大。膜翅目、真螨目、寄瞒目与弹尾目则受土壤容重、乔木层生物量的影响较大。微型土壤生物(细菌、真菌、放线菌、原生动物)以及地蜈蚣目、石蜈蚣目等受草本、灌木层生物量影响大,此外,土壤微生物与土壤pH、TK、TP等呈现正相关。
远郊区土壤含水率、有机质、铵态氮、硝态氮等理化性状干湿季差异显著,说明远郊区土壤理化性状等受季节影响波动大。与干季相比,湿季的后孔寡毛目生态㶲虽有增加,但其在城区与近郊区森林环境中增加的幅度更大,说明城区与近郊区森林的土壤含水率等理化性状更稳定,而稳定的土壤环境更适合地下生物后孔寡毛目、马陆类的生长。受城市化影响最少的远郊区森林中的草本层植物群落的生物量最高(黄柳菁,2012),而土壤微生物受草本层的正相关影响最大,其生态㶲在远郊区也相应最高(图4)。
4 结论与讨论
4.1讨论
本研究表明,除了气候条件和人为干扰等可能影响因子之外(易志刚等,2005;徐国良等,2006;Murrary等,2006),城市化对南亚热带常绿阔叶林土壤生物的影响因子主要有土壤含水率、铵态氮、有机质、硝态氮以及草本层生物量等。
城市化会降低南亚热带常绿阔叶林土壤微生物群落生物量和生态㶲,并加剧干、湿季差异(易志刚等,2005;吴艺雪等,2009)。广州市城郊梯度上残存的南亚热带常绿阔叶林中,城市化可能通过影响草本层、土壤酸度等来影响土壤微生物,结果表明草本层生物量大、土壤酸度较小的远郊区土壤微生物生物量最高。在本研究城郊梯度上,远郊区林下土壤微生物生物量在干季显著高于近郊区和城区,在湿季显著高于城区;近郊区土壤微生物量在湿季显著高于城区,在湿季不显著低于城区。表明城市化会降低南亚热带常绿阔叶林土壤微生物生物量,这与陈帅等(2012)对北京城-郊温带森林土壤微生物群落的研究结果一致。

表3 各植被类型的主要类群的生态㶲及其所占比例Table 3 Eco-exergy and proportions of main groups

表4 各样点干湿两季土壤理化性质特征Table 4 Soil physicochemical properties at the three sites on the urban-rural gradient
适度的城市化干扰可增加南亚热带常绿阔叶林土壤动物类群数和生态㶲,并降低干、湿季差异(Connell,1978)。本研究结果表明,在广州市城郊梯度上,南亚热带常绿阔叶林土壤动物个体数目无显著差异,但近郊区的类群数显著高于远郊区和城区。同时,近郊区林下土壤动物个体数和类群数的差异均不显著,而远郊区和城区则是干季显著高于湿季。中度城市化干扰下的近郊区林下土壤动物群落类群数和生态㶲土壤动物中后孔寡毛目对森林土壤含水率、有机质、铵态氮比较敏感,上述土壤理化指标相对高的湿季更适合后孔寡毛目的生存,而土壤中后孔寡毛目的增多,使枯枝落叶的分解过程加速,促进土壤有机质含量的累积。
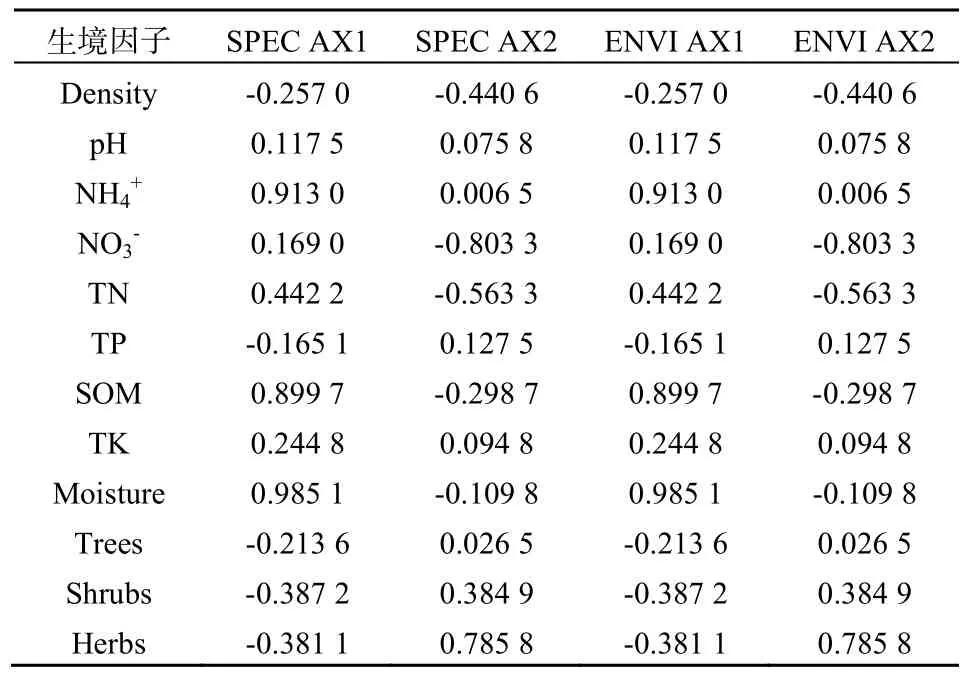
表5 环境因子与排序轴的相关性Table 5 Correlation coefficients between ordination axis and environment factors
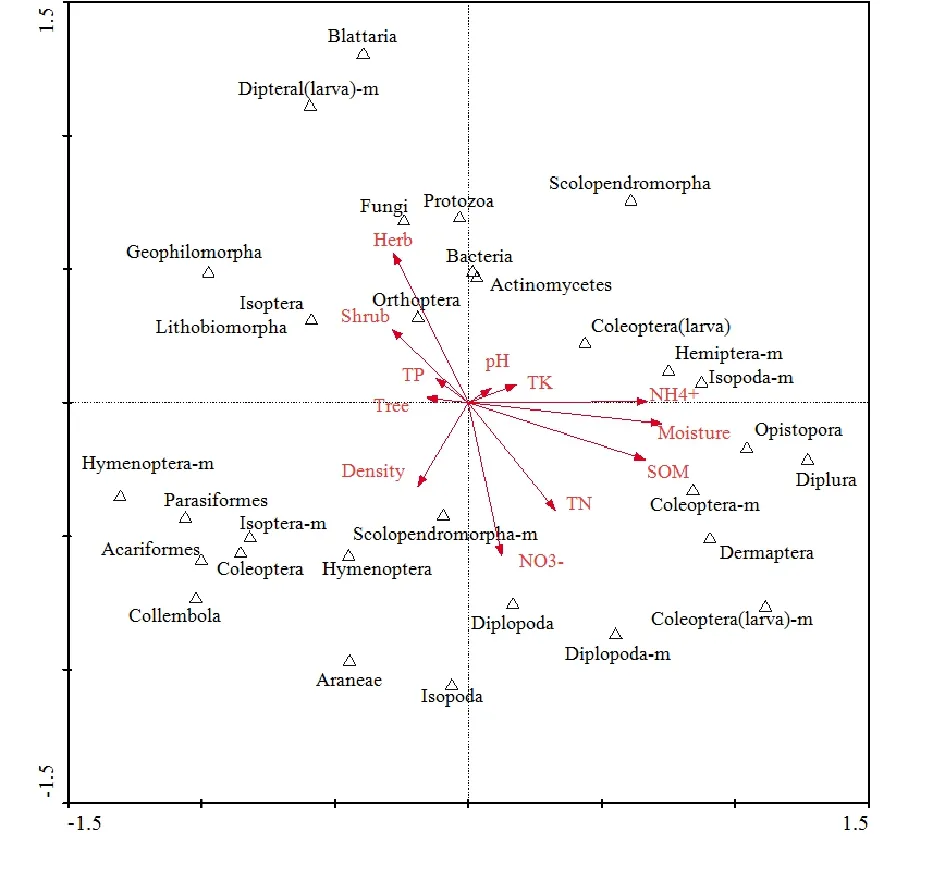
图4 土壤生物生态㶲与生境因子关系的RDA双序图Fig. 4 Biplot got by RDA illustrated the effects of environment factors and eco-exergy of soil organisms
综合而言,广州近郊和城区残留的南亚热带常绿阔叶林土壤生物群落生态㶲与结构㶲值均高于远郊区的水平;南亚热带常绿阔叶土壤生物群落生态㶲主要由大型土壤动物贡献,而中小型土壤动物则是结构㶲的主要贡献者;不考虑生命周期,土壤微生物对于土壤生物群落生态㶲与结构㶲值的贡献较小。相对于远郊区坡度较大的山林环境,近郊区与城区平缓地稳定的土壤理化环境更适宜林下土壤生物群落生态㶲的主要贡献者后孔寡毛目、马陆类等的生长。综合本研究结果与秦钟(2009)等对广州市天河区林地、人工绿地以及农田土壤中小型动物群落结构的研究结果,我们认为,南亚热带常绿阔叶林土壤生物群落生物热力学组织程度与健康水平较高,但如果保护不良,土地利用后的恢复难度亦较大。综合本研究结果与黄柳菁(2012)对广州市城郊梯度南亚热带常绿阔叶林植被群落结构的研究结果,我们认为,远郊区大岭山森林植被与土壤生物群落的生物热力学组织程度与健康水平均有待进一步提高,相关林业管理与保育工作任重道远。
生态㶲理论方法可以综合度量土壤生物群落的自组织程度与健康水平,但在具体计量中受到权重因子β的有限性制约。目前,大多数生态㶲权重因子β只细化到目的分类水平,仅少部分细至门、纲的分类水平,难以满足高生物多样性土壤生物群落生态㶲的精确计量需要,亟待补充、完善。
4.2结论
城市化对南亚热带常绿阔叶林土壤微生物和土壤动物群落结构均有影响,但影响的方向、速率和机制各不相同。在广州市城郊梯度上,远郊区的南亚热带常绿阔叶林土壤生物群落生物热力学组织程度与健康水平有待提高。综合而言,南亚热带常绿阔叶林土壤生物群落生物热力学组织程度较高,开发利用需谨慎考虑。
参考文献:
AUSTONI M, GIORDANI G, VIAROLI P, et al. 2007. Application of specific exergy to macrophytes as an integrated index of environment quality for coastal lagoons [J]. Ecological Indicators, 7(2): 229-238.
CONNELL J H. 1978. Diversity in tropical rain forest and coral reefs [J]. Science, 199: 1302-1310.
HUANG L J, CHEN H F, REN H, et al. 2013. Effect of urbanization on the structure and functional traits of remnant subtropical evergreen broad-leaved forests in South China [J]. Enviromental Monitoring and Assesment, 185(6): 5003-5018.
JØRGENSEN S E, LADEGAARD N, DEBELJAK M, et al. 2005. Calculations of exergy for organisms [J]. Ecological Modelling, 185(2-4): 165-175.
JØRGENSEN S E, MEJER H. 1977. Ecological buffer capacity [J]. Ecological Modelling, 3(1): 39-45, 47, 49, 51, 53-61.
JØRGENSEN S E, MEJER H. 1979. A holistic approach to ecological modeling [J]. Ecological Modelling, 7(3): 169-189.
JØRGENSEN S E, VERDONSCHOT P, LEK S. 2002. Explanation of the observed structure of functional feeding groups of aquatic macro-invertebrates by an ecological model and the maximum exergyprinciple [J]. Ecological Modelling, 158(3): 223-231.
JØRGENSEN S E. 2006. Eco-exergy as Sustainability [M]. Southampton Boston, WIT Press: 8-20.
JØRGENSEN S E. 2007. Description of aquatic ecosystem’s development by eco-exergy and exergy destruction [J]. Ecological Modelling, 204(1-2): 22-28.
LIBRALATO S, TORRICELLI P, PRANOVI F. 2006. Exergy as ecosystem indicator: an application to the recovery process of marine benthic communities [J]. Ecological Modelling, 192(3-4): 571-585.
LOTKA A J. 1922a. Contribution to the energetic of evolution [J]. Proceedings of the National Academy of Sciences, 8(6): 147-151.
LOTKA A J. 1922b. Natural selection as a physical principle [J]. Proceedings of the National Academy of Sciences, 8(6): 151-154.
LU H F, WANG Z H, CAMPBELL D E, et al. 2011. Emergy and eco-exergy evaluation of four restoration modes in southeast China [J]. Ecological Engineering, 37(2): 277-285.
MOLOZZI J, SALAS F, CALLISTO M, et al. 2013. Thermodynamic oriented ecological indicators: application of eco-exergy and specific eco-exergy in capturing environmental changes between disturbed and no-disturbed tropical reservoirs [J]. Ecological Indicators, 24: 543-551.
MURRARY P J, COOK R, CURRIE A F, et al. 2006. Interactions between fertilizer addition,plants and soil environment : implications for soil faunal structure and diversity [J]. Applied Soil Ecology, 33(2): 199-207.
XU F, YANG Z F, CHEN B, et al. 2011. Ecosystem health assessment of the plant-dominated Baiyangdian lake based on eco-exergy [J]. Ecological Modelling, 222(1): 201-209.
ZHAO Z X, GUO H C. 2010. Effects of urbanization on the quantity changes of microbes in urban-to-rural gradient forest soil [J]. Agricultural Science & Technology, 11(3): 118-122.
ZHOU G Y, LIU S G, LI Z A, et al. 2006. Old-growth forests can accumulate carbon in soils [J]. Science, 314(5804): 1417.
陈帅, 王效科, 逯非. 2012. 城市与郊区森林土壤微生物群落特征差异研究[J]. 土壤通报, 43(3): 614-620.
傅声雷. 2007. 土壤生物多样性的研究概况与发展趋势[J]. 生物多样性, 15(2): 109-115.
黄柳菁. 2012. 城市化影响下存留南亚热带常绿阔叶林群落特征及其环境因子关联研究[D]. 中国科学院华南植物园: 77-81.
李浩, 曾宪曙, 陆宏芳. 2012. 基于Eco-exergy的鹤山红木荷人工林群落结构发展动态[J]. 生态环境学报, 21(12): 1822-1829.
李俊生, 高吉喜, 张晓岚, 等. 2003. 城市化对生物多样性的影响研究综述[J]. 生态学杂志, 24(8): 953-957.
廖崇惠, 李健雄, 黄海涛. 1997. 南亚热带森林土壤动物群落多样性研究[J]. 生态学报, 17(5): 549-555.
彭少麟. 2003. 热带亚热带恢复生态学研究进展与实践[M]. 北京: 科学出版社: 7-18.
秦钟, 章家恩, 李庆芳. 2009. 城市化地区不同生境中中小型土壤动物群落结构特征[J]. 应用生态学报, 20(12): 3049-3056.
王绍军, 阮宏华. 2008. 土壤生物对地上生物的反馈作用及其机制[J].生物多样性, 16(4): 407-416.
吴艺雪, 杨晓东, 余广彬. 2009. 两种热带雨林微生物生物量碳季节动态及其影响因素[J]. 生态环境学报, 18(2): 658-663
徐国良, 周国逸, 莫江明, 等. 2006. 南亚热带退化植被重建中土壤动物群落变化[J]. 动物学研究, 27(1): 23-28.
杨冬青, 高峻. 2002. 城市化生态系统中土壤动物研究及应用进展[J].生态学杂志, 21(5): 54-57.
易志刚, 蚁伟民, 周丽霞, 等. 2005. 鼎湖山主要植被类型土壤微生物生物量研究[J]. 生态环境, 14(5): 727-729.
赵哲霞, 郭恢财. 2010. 城市化对城乡梯度森林土壤微生物数量变化的影响[J]. 安徽农业科学, 38(10): 5188-5190.
Effects of Urbanization on Soil Community Structure under Subtropical Evergreen Broad-leaved Forests
FU Fangyan1, 2, LU Hongfang1*
1. South China Botanical Garden, University of Chinese Academy of Sciences, Guangzhou 510650, China; 2. University of Chinese Academy of Sciences, Beijng 100049, China
Abstract:The impact of urbanization on community structure of soil microbes and soil fauna under Subtropical Evergreen Broadleaf Forest (SEBF) was measured along an urban-suburban-rural gradient in Guangzhou, China. Furthermore, the impact on the biophysical structure of the soil community was quantified using the eco-exergy method. The results showed that the impact of urbanization on soil microbes and soil fauna was different in both direction and degree, and that these differences were mainly related to soil moisture, ammonium and nitrate nitrogen, soil organic matter, and the biomass of herbs. Urbanization can decrease the biomass and eco-exergy of soil microbes under SEBF, and increase the differences between the dry and wet seasons; while a low degree of urbanization can increase the group richness as well as the eco-exergy of soil fauna, and decrease the differences between dry and wet seasons. After counting both the soil microbes and soil fauna, the eco-exergy of the whole soil community at the suburban study site was higher than that at the urban and rural study sites. Larger fauna is the main contributor to the eco-exergy of the soil community (>64.41%), while middle and small fauna contribute more to the specific eco-exergy (>50.00%). The eco-exergy method can measure the biophysical organization of the soil community, but its application is limited by the availability of weighting factors.
Key words:urbanization, soil microbes; soil fauna; community structure; eco-exergy
收稿日期:2015-03-25
*通信作者:陆宏芳,E-mail: luhf@scbg.ac.cn
作者简介:符方艳(1990年生),女,硕士研究生,研究方向为生态恢复与生态经济。E-mail: fufy5826@163.com
基金项目:国家自然科学基金项目(31170428)
中图分类号:X171.1; Q948
文献标志码:A
文章编号:1674-5906(2015)06-0938-09
DOI:10.16258/j.cnki.1674-5906.2015.06.005
猜你喜欢
哲学评论(2021年2期)2021-08-22
中华诗词(2019年7期)2019-11-25
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
影视与戏剧评论(2016年0期)2016-11-23
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
现代企业(2015年9期)2015-02-28
