
壮精合剂对初老大鼠行为学及脑单胺类神经递质的影响*
2015-03-08 02:10蔡浩斌黄忠仕黄岑汉
重庆医学 2015年2期
蔡浩斌,黄忠仕,黄岑汉△
(1.广州中医药大学临床药理研究所,广州510405;2.右江民族医学院,广西百色533000)
壮药补精合剂(简称壮精合剂)是壮族民间医生根据壮医学理论的治疗原则,从精血论治的角度,用壮药(土人参、绞股蓝、扶芳藤、大地棕根等)配伍而成的经验方。方中重用土人参补精血、清虚热、调经带,为填精补血之要药;扶芳藤通龙路火路,舒筋脉,散淤血、止出血,通路活血以强土人参补精血之势;绞股蓝能除虚热、促食欲,促睡眠,抗疲劳、抗衰老,含有多种人参皂苷和氨基酸,有“南方人参”之称;大地棕根能补肾填精,活血化瘀,具有补虚通路之效。全方用药精专,配伍得当,剂量适宜,共奏补精益气、养血调经之功效,从而使精气盛、气血调、龙路通。本研究主要根据最新的衰老发生机制及新时期探讨衰老机制的新途径,以壮精合剂作用于初老大鼠模型,研究壮精合剂对初老大鼠行为学、单胺类神经递质等因素的影响,探讨壮精合剂抗衰老的作用及其机制,为开发广西的抗衰老中草药及申报抗衰老新药提供理论依据。现将实验方法及结果报道如下。
1 材料与方法
1.1 材料
1.1.1 实验动物 选用12月龄健康雌性SD大鼠50只,体质量平均(300±20)g,由右江民族医学院实验动物中心提供(许可证:SCXK桂2012-0003)。自动控制光照,在通风、温度及湿度适宜的动物室内常规饲养。
1.1.2 药物及试剂 壮精合剂(土人参30g,绞股蓝25g,扶芳藤20g,大地椋根10g);阳性对照药:古汉养生精(批号902630-126904);标准品:去甲肾上腺素(NE),批号100169-201103;多巴胺(DA),批号100070-201006;5-羟色胺(5-HT),批号111656-50679,均购自中国药品生物制品检定所;流动相:乙腈、纯净水;磷酸二氢钾;EDTA-2Na;样品前处理试剂:高氯酸。
1.1.3 仪器 Morris水迷宫(中国科学院药研所制),微电极脑立体定位仪(WDT-11,西北光明仪器厂)。FJ-200高速分散均质机(上海标本模型厂),高速冷冻离心机(BACKMAN公司,美国),高效液相-荧光检测器(Waters公司,美国),电子分析天平(Sartorius公司,德国)。
1.2 方法
1.2.1 模型建立 采用自然老化法。初老大鼠模型:选12月龄雌性大鼠,采取阴道脱落细胞涂片巴氏染色,连续观察4个动情周期,以阴道细胞学表现动情周期长。之后持续动情、反复假妊娠时,作为自然衰老的初老大鼠模型[1-2]。
1.2.2 分组与给药 将造模成功的50只龄雌性SD初老大鼠随机分为5组,分别为空白对照组、壮精合剂高剂量组、壮精合剂中剂量组、壮精合剂组低剂量组、阳性对照组,每组10只。空白对照组给予纯净水10mL/kg,阳性对照组给予古汉养生精4.8g/kg,壮精合剂低、中、高剂量组分别给予壮精合剂6 g/kg、15g/kg、30g/kg。以上各组每天灌胃给药1次,连续8周。
1.2.3 观察项目及方法
1.2.3.1 学习记忆能力测试[3]采用 Morris水迷宫检测各组大鼠的空间学习记忆行为能力,包括定位航行实验和空间探索实验。其中定位航行实验历时5d,每天将大鼠面向池壁分别从不同象限(靶象限除外)置入池中,记录其寻找到隐藏在水面下平台的时间即逃避潜伏期。大鼠登上站台1s后终止记录,最长记录时间为100s,若大鼠在100s内不能上台,则引导其登上站台适应20s。空间探索实验是在定位航行实验后撤掉平台,然后将大鼠随机从不同象限(靶象限除外)面壁置入池中,记录其在100s内的游泳轨迹,穿过站台的次数,考察大鼠对原平台的记忆。
1.2.3.2 脑组织内单胺类神经递质含量的测定[4-5]采用高效液相-荧光测定法检测大鼠大脑皮层、海马区单胺类神经递质含量。
1.2.3.2.1 色谱条件 色谱柱:Kromasil C18反相色谱柱(250mm×4.6mm,5μm)。流动相为水:乙腈(90∶10),其中水相每升含磷酸二氢钾0.2mol、EDTA-2Na 0.1mol,用磷酸调pH值至3.0,流速0.5mL/min。发射波长338nm,激发波长220nm。柱温:28℃,进样量10μL。在本色谱条件下,NE,DA和5-HT各峰均达基线分离的效果。色谱图见图1。
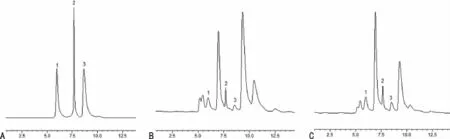
图1 色谱图
1.2.3.2.2 对照品溶液的制备 取混合对照品贮备液适量,置6个容量瓶中,各加入5%HCLO4稀释成不同质量浓度的混合对照品溶液,使所含对照品的质量浓度分别为0.002、0.010、0.050、0.500、1.000、5.000μg/mL,分别精密吸取10 μL进样。以各组分的质量浓度为横坐标,各组分的峰面积为纵坐标进行线性回归,并计算相关系数r。结果表明,各标准曲线线性良好。各回归方程如下:DA;Y=230 178X+3 227.2,r=0.999 9;NE:Y=219 207X+2 087.9,r=0.999 9;5-HT:Y=386 571X-7 865.5,r=0.999 9。
1.2.3.2.3 样品处理 实验大鼠断头处死后,迅速取出全脑,用冰冷生理盐水冲洗除去血液,滤纸拭干,冰上迅速分离出大鼠大脑皮层、海马组织,称质量后,加入1mL 5%HClO4,超声粉碎成10%匀浆,冷冻高速离心14 000r/min 15min,取上清液,即得脑组织样品。取上清液进样10μL。记录各样品的峰面积,代入相应标准曲线计算出相应浓度,统计结果。
1.3 统计学处理 采用SPSS17.0软件处理,计量资料用±s表示,组间比较采用t检验,检验水准α=0.05,以P<0.05为差异有统计学意义。
2 结 果
2.1 Morris水迷宫实验结果 定位航行:壮精合剂中剂量组及阳性对照组与空白对照组比较所用时间更少(P<0.05)。壮精合剂高剂量组比空白对照组、阳性对照组所用时间更少(P<0.05)。空间探索实验:壮精合剂中剂量组及阳性对照组与空白对照组相比次数更多(P<0.05)。壮精合剂高剂量组与空白对照组、阳性对照组相比次数较多(P<0.05)。结果说明壮精合剂能明显提高初老大鼠的学习能力和记忆能力,且高剂量组效果最为显著,见表1。
表1 隐秘平台实验与空间探索实验结果(±s)

表1 隐秘平台实验与空间探索实验结果(±s)
a:P<0.05,b:P<0.01,与空白对照组比较;c:P<0.05,与阳性对照组比较。
组别 定位航行(潜伏期,s)穿越平台次数(n)空白对照组22.62±4.18 5.15±2.57壮精合剂低剂量组 21.67±3.49 4.51±2.50壮精合剂中剂量组 18.84±3.50a 7.50±1.85a壮精合剂高剂量组 15.14±3.64bc 9.42±2.27bc阳性对照组 18.77±3.90a 7.40±1.88a
大鼠Morris水迷宫实验轨迹类型分为4种:趋向型、直线型、随机型、边缘型。其中每只大鼠共游泳10次,对比各组大鼠游泳轨迹,阳性对照组大鼠趋向型、直线型游泳轨迹有63次,空白对照组大鼠趋向型、直线型游泳轨迹有44次,两者相比差别显著(P<0.05)。壮精合剂中剂量组大鼠趋向型、直线型游泳轨迹有68次,与空白对照组相比差异有统计学意义(P<0.05)。壮精合剂高剂量组大鼠趋向型、直线型游泳轨迹有80次,与空白对照组、阳性对照组相比差异有统计学意义(P<0.01)。结果说明壮精合剂能明显提高衰老大鼠的学习能力和记忆能力,且高剂量组效果最为显著(表2、图2).
2.2 各组大鼠大脑皮层内单胺类神经递质含量 各组大鼠大脑皮层内单胺类神经递质NE、DA、5-HT含量均比空白对照组高,且壮精合剂高剂量组效果显著(与空白对照组比较,P<0.01;与阳性对照组比较,P<0.05),见表3。
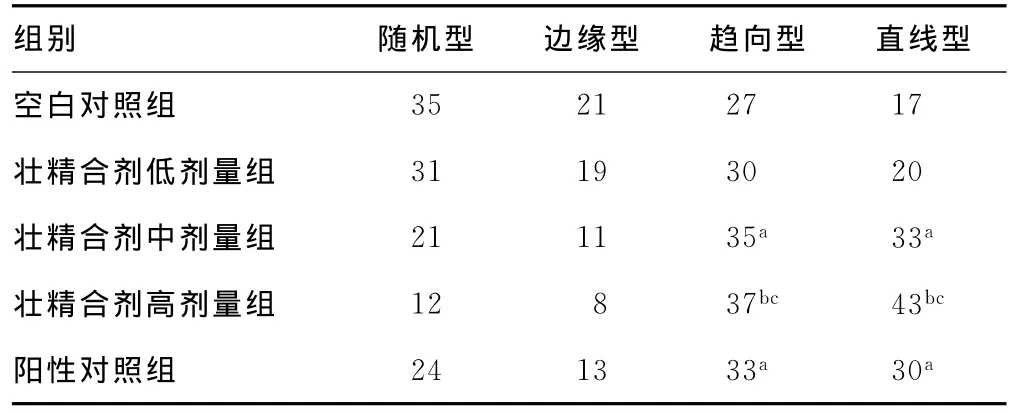
表2 大鼠Morris水迷宫游泳轨迹类型(n,n=10)

图2 Morris水迷宫轨迹图
表3 各组大鼠大脑皮层内单胺类神经递质含量比较(±s,ng/g,n=10)

表3 各组大鼠大脑皮层内单胺类神经递质含量比较(±s,ng/g,n=10)
a:P<0.05,b:P<0.01,与空白对照组比较;c:P<0.05,与阳性对照组比较。
组别DA NE 5-HT空白对照组387.3±84.5 253.6±46.3 114.3±29.1壮精合剂低剂量组 403.8±64.1 287.1±36.5 136.2±25.5壮精合剂中剂量组 468.3±73.8a313.5±41.9b 168.4±33.2b壮精合剂高剂量组 527.3±81.5bc 373.6±46.3bc 211.8±27.7bd阳性对照组 457.5±61.6a328.4±39.1b 166.7±31.1b
2.3 各组大鼠海马单胺类神经递质含量 各组大鼠海马内单胺类神经递质NE、DA、5-HT含量均比空白对照组高,且壮精合剂高剂量组效果显著(与空白对照组比较P<0.01;与阳性对照组比较P<0.05),见表4。
表4 各组大鼠海马单胺类神经递质含量比较(±s,ng/g,n=10)
表4 各组大鼠海马单胺类神经递质含量比较(±s,ng/g,n=10)
a:P<0.05,b:P<0.01,与空白对照组比较;c:P<0.05,与阳性对照组比较。
组别DA NE 5-HT空白对照组117.4±32.3 53.6±16.6 44.9±11.5壮精合剂低剂量组 133.1±24.2 67.3±19.2 49.3±15.7壮精合剂中剂量组 168.6±23.1b 74.6±19.3a 63.8±13.1b壮精合剂高剂量组 187.9±21.5bd 108.5±26.4bd 77.4±17.9bd阳性对照组 152.1±31.5a73.5±19.1a56.3±12.6a
3 讨 论
衰老是指在正常状况下生物发育成熟后,随年龄增加,自身机能减退,内环境稳定能力与应激能力下降,结构、组分逐步退行性变,趋向死亡的不可逆转现象[6]。中医药以其独特疗效,在抗衰老抗氧化的研究中占有越来越重要的地位。中医认为衰老主要是肾虚,其次是脾虚,再次是气血两虚。所以补肾、健脾、益气是延缓衰老的基本途径,活血化淤是延缓衰老的主要方法[7]。
学习记忆是大脑的高级神经功能之一,中枢单胺类神经递质在此过程中发挥了重要的作用[8]。其过程伴随着复杂的神经生理和生化机制,其作用机制与单胺类神经递质含量密切相关[9]。研究表明,去甲肾上腺素在记忆保持方面起着重要作用,而改善记忆与多巴胺的释放有关[10]。去甲肾上腺素和多巴胺含量的减少是造成学习记忆能力减退的重要原因[11]。它们的前体是酪氨酸和色氨酸,酸性代谢产物有3,4-二羟苯乙酸、5-羟吲哚乙酸等。这些物质在正常生物体内存在动态平衡,参与中枢神经系统中多种生理过程。分析单胺类神经递质及其相关物质可为多种疾病的诊断和治疗以及疾病机制的研究提供重要依据[12-15]。
在正常情况下,中枢内神经递质的分泌保持在一定水平,并且它们相互之间比例协调,从而维持功能的稳定;随着机体的老化,学习记忆能力的下降,脑内单胺类神经递质的代谢亦发生紊乱,出现NE、DA、5-HT含量的下降,提示衰老过程中脑内单胺类神经递质的平衡遭到破坏,这与一些老年性疾病如帕金森病、老年痴呆及脑功能的减退也有关[16]。
本实验结果显示给予壮精合剂灌胃8周以后,各组大鼠大脑皮层及海马内单胺类神经递质NE、DA、5-HT含量均比空白对照组高,且壮精合剂高剂量组效果显著。结合Morris水迷宫实验结果说明壮精合剂可明显改善初老大鼠的学习记忆能力及提高大脑皮层、海马内单胺类神经递质NE、DA、5-HT含量,提示壮精合剂可有效调整中枢神经递质的合成,恢复大脑皮层、海马内单胺类神经递质的平衡,改善大脑皮层、海马神经内分泌细胞的功能,且壮精合剂高剂量效果较好。
[1] 谢启文.现代神经内分泌学[M].上海:上海医科大学出版社,1999:385-389.
[2] 程化奇,熊舜华,殷新尤,等.大鼠更年期实验模型研究[J].上海实验动物科学,1993,13(4):192-196.
[3] 陈罗西,郭玲玲,李亮.Morris圆形水迷宫的应用及其相关检测指标分析[J].辽宁中医药大学学报,2008,10(8):55-57.
[4] 魏元锋,汪益涵,张宁,等.HPLC-荧光检测法测定大鼠血浆及脑组织中5种单胺类神经递质[J].中华中医药学刊,2009,27(9):1942-1943.
[5] 张璐璐,冯芳,叶小敏,等.高效液相色谱-荧光法测定小鼠脑组织中单胺递质及其相关物质[J].中国医科大学学报,2010,41(4):367-371.
[6] 邹承鲁.当代生物学[M].北京:中国致公出版社,2000:394-396.
[7] 石雪蓉.衰老机制和抗衰老药物的研究现状[J].四川解剖学杂志,2008,16(3):43-45.
[8] 朱蕾,张茹,李廷利.刺五加对睡眠剥夺大鼠学习记忆及海马单胺类神经递质的影响[J].中国实验方剂学杂志,2012,18(4):219-222.
[9] Birthelmer A,Stemmelin J,Jackisch R,et al.Presynaptic modulation of acetylcholine,noradrenalin,and serotonin release in the hippocampus of aged rats with various levels of memory impairments[J].Brain Res Bull,2003,60(3):283-296.
[10] Nowakowska E,Chodera A,Kus K,et al.Reversal of stress-induced memory changes by moclobemide:the role of neurotransmitters[J].Pol J Pharmacology,2001,53(3):227-233.
[11] De la Fuente M,Hernanz A,Medina S,et al.Characterization of monoamine systems in brain regions of prematurely ageing mice[J].Neurochemistry Int,2003,43(2):165-172.
[12] Hamaue N,Ogata A,Terado M,et al.Entacapone,a catechol-O-methyltransferase inhibitor,improves the motor activity and dopamine content of basal ganglia in a rat model of Parkinson′s disease induced by Japanese encephalitis virus[J].Brain Res,2010,1309:110-115.
[13] Meyer U,Feldon J.Neural basis of psychosis-related behavior in the infection model of schizophrenia[J].Behav Brain Res,2009,204(2):322-334.
[14] Moreno FA,Parkinson D,Palmer C,et al.CSF neurochemicals during tryptophan depletion in individuals with remitted depression and healthy controls[J].Eur Neuropsychopharm,2010,20(1):18-24.
[15] Hirschfeld RM.History and evolution of the monoamine hypothesis of depression[J].J Clin Psychiatry,2000,61(Suppl 6):4-6.
[16] Rehman Hu,Masson EA.Neuroendocrinology of aging[J].Age Ageing,2001,30(4):279-287.
猜你喜欢
中国图书评论(2022年4期)2022-11-22
世界科学技术-中医药现代化(2022年3期)2022-08-22
右江医学(2022年3期)2022-04-17
食品安全导刊(2021年21期)2021-08-30
大众健康(2021年2期)2021-03-09
河北果树(2020年2期)2020-05-25
中成药(2018年7期)2018-08-04
中成药(2018年5期)2018-06-06
中成药(2017年7期)2017-11-22
中国学术期刊文摘(2016年8期)2016-02-13
