
SIRT1和SIRT2基因多态性及合并基因型与秦川牛肉用性状的关联分析
2015-03-07 08:05:16桂林生昝林森王洪宝王洪程
畜牧兽医学报 2015年10期
桂林生,昝林森,2*,王洪宝,2,王洪程
(1.西北农林科技大学动物科技学院,杨凌712100;2.国家肉牛改良中心,杨凌712100)
信息沉默调节因子1(Silent mating type information regulator 1,SIRT1)属于Sirtuin家族,是一类依赖烟碱胺腺嘌呤(NAD+)的第Ⅲ类组蛋白去乙酰化酶,定位于细胞核中且具有高度保守性[1-2]。在人和动物机体的糖代谢、脂代谢和胰岛素分泌等生理过程中,SIRT1发挥着重要的调节作用。作为调节脂肪形成最主要的转录因子,PPARγ基因被证实可以调控脂肪酸的储存和葡萄糖的代谢[3],研究表明,SIRT1基因可以通过与SMRT基因相结合,抑制PPARγ的活性,进而降低脂肪细胞分化速度,减少脂肪沉积[4]。此外,作为SIRT1基因的有效激活剂,白藜芦醇饲喂的小白鼠在抵抗高脂饮食引起的肥胖效果显著高于对照组,这说明SIRT1基因可以调节脂质代谢过程[5-7]。信息沉默调节因子2(Silent mating type information regulator 2,SIRT2)同样属于Sirtuin家族,主要定位于细胞质中,与微管共定位,完成对α-微管蛋白的去乙酰化作用,在结构上与SIRT1具有高度同源性[8-9]。在小鼠3T3-L1细胞上的研究发现,超表达SIRT2基因可以有效抑制前体脂肪细胞的分化[10],这可能是因为SIRT2基因可以结合并去乙酰化Foxo1基因,促进Foxo1基因对PPARγ基因的抑制作用[11]。类似的结果在猪的前体脂肪细胞中同样被证实[12]。
肉用性状是畜牧业生产过程中一个重要的经济性状,是衡量育种价值及经济效益的重要指标。鉴于这两个基因在脂质代谢过程中的重要作用,推测其在畜禽的肉用方面可能发挥直接或间接的作用,目前尚未见到相关报道。因此,本研究以SIRT1和SIRT 2基因为目标基因,采用PCR-PFLP方法分析不同标记基因型及合并基因型对秦川牛肉用性状的影响,以期为秦川牛分子育种提供新的理论方法和技术途径。
1 材料与方法
1.1 试验材料
血样采自陕西省良种肉牛繁育中心、陕西省秦川牛保种场和西北农林科技大学良种肉牛繁育中心3个场区,随机选择505头同等饲养条件下无亲缘关系的18~24月龄健康的秦川牛,对每头牛颈静脉采血10 m L,ACD抗凝(V(ACD)∶V(血液)=1∶6),置于-80℃ 保存备用。肉用性状包括背膘厚、眼肌面积和肌内脂肪含量3个指标。本试验肉用性状使用超声波背膘仪测定,测定之前在牛背部第12肋骨附近上下均匀涂上清油,等待1~2 min后开始检测。
1.2 秦川牛基因组DNA的提取及检测
采用常规的酚-氯仿提取法[13]提取秦川牛血样基因组DNA,提取后的DNA用TE缓冲液溶解后,采用0.5%琼脂糖凝胶电泳检测DNA质量,紫外分光光度计测定DNA浓度,然后-20℃保存备用。
1.3 秦川牛SIRT1和SIRT2基因PCR扩增
根据GenBank公布的牛SIRT1(NM_001192980.1)和SIRT2的序列(NM_001113531.1),利用Primer 5.0软件设计引物(表1),引物均由上海生工生物工程技术服务有限公司合成。
PCR反应体系30.0μL,含有核酸染料的d NTPs、Taq DNA聚合酶、10×Buffer的Mix 15.0 μL,dd H2O 11.8μL,上、下游引物各(10 pmol·μL-1) 0.6μL、模板DNA(50 ng·μL-1)2.0μL。PCR反应程序:95℃预变性5 min;94℃30 s,退火30 s(退火温度见表1),72℃30 s,35个循环;72℃延伸10 min;4℃保存。PCR扩增产物用10 g·L-1琼脂糖凝胶电泳检测。
1.4 PCR产物纯化与测序
在扩增好的PCR产物中,每种随机选取8个样本使用凝胶回收试剂盒回收纯化,送至上海生工生物工程技术服务有限公司进行测序,测序结果使用DNAMAN软件进行对比分析,寻找突变位点。
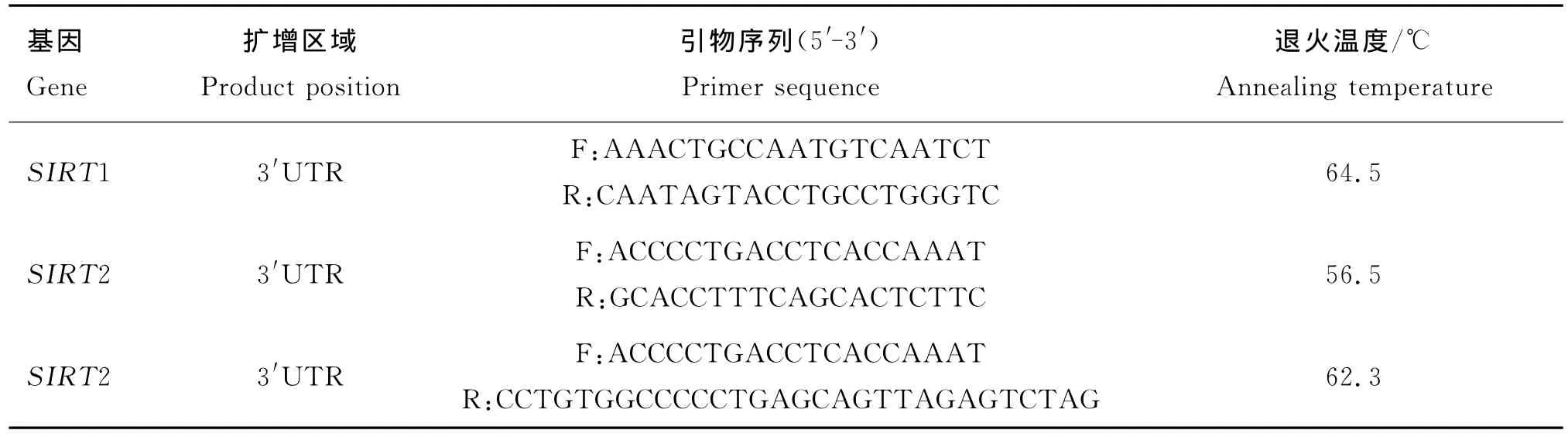
表1 秦川牛SIRT1和SIRT2基因引物序列信息Table 1 Sequence information of primers amplifying Qinchuan cattle in SIRT1 and SIRT2 genes
1.5 PCR-PFLP分析
经测序比对后,取含有突变位点的PCR扩增产物10μL,依照所测突变位点G25764A、A25846G、C19501T和C19518T的顺序,分别加入限制性内切酶Bfa 1(Fermentas,Canada)、Sat 1(Fermentas,Canada)、Tai 1(Fermentas,Canada)和Xba 1(Fermentas,Canada)1.0μL,酶切10×Buffer 2.0μL,加入去离子水至20μL,反应体系置于37℃恒温箱中过夜。然后用2%的琼脂糖凝胶电泳检测,凝胶成像系统照相分析。
1.6 数据统计分析
根据基因型统计分析结果计算基因型、等位基因频率、多态信息含量(PIC)、遗传杂合度(He)和有效等位基因数(Ne),并进行Hard-Weinberg平衡适应性检验。采用SPSS(13.0 version)软件[14]分别对SIRT 1和SIRT 2基因突变位点的合并基因型与秦川牛部分肉用性状(背膘厚、眼肌面积和肌间脂肪含量)进行相关性分析。同时,分析SIRT1和SIRT 2基因之间的合并基因型与秦川牛肉用性状的关联性。单基因效应和多基因效应方差分析模型:单基因效应方差分析:表型观察值=群体均值+标记基因型效应+年龄效应+随机误差;合并基因型方差分析:表型观察值=群体均值+SIRT 1基因型+SIRT 2基因型+SIRT 1与SIRT 2互作效应+随机误差。
2 结 果
2.1 秦川牛SIRT1和SIRT2基因PCR扩增
根据牛SIRT1(NM_001192980.1)和SIRT2的序列(NM_001113531.1),利用Primer 5.0软件设计引物,而后对SIRT 1和SIRT 2基因进行PCR扩增,结果见图1。
2.2 SIRT1和SIRT2不同位点基因型分析
牛SIRT1基因位于第28号染色体上,基因全长26 517 bp,含9个外显子。通过测序分析发现,SIRT1基因3′侧翼区存在2处SNPs,分别是G25764A(图2A)和A25846G(图2B)。G25764A位点存在GG和GA两种基因型,命名为AA和AB;A25846G位点存在AA、AG和GG 3种基因型,分别命名为AA、AB和BB。牛SIRT2基因位于第18号染色体上,基因全长19 838 bp,含16个外显子。本试验同样在SIRT 2基因3′侧翼区检测到2个SNPs,分别是C19501T(图3A)和C19518T(图3B),C19501T位点上发现CC和CT两种基因型,命名为AA和AB;C19518T位点存在CC、CT和TT 3种基因型,分别命名为AA、AB和BB。具体酶切效果见图4和图5。
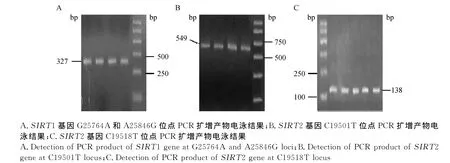
图1 SIRT1和SIRT2的PCR扩增产物电泳结果Fig.1 Detection of PCR products of SIRT1 and SIRT2

图2 SIRT1基因G25764A(A)和A25846G(B)基因型测序图Fig.2 Sequencing diagram at site G25764A(A)and A25846G(B)of SIRT1 gene
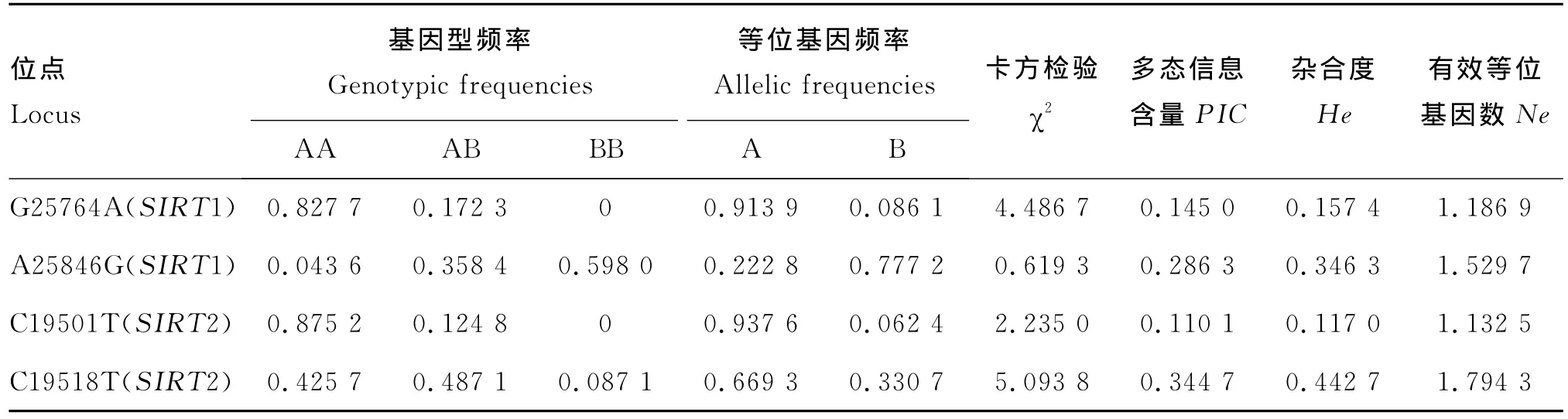
表2 秦川牛SIRT1和SIRT2不同位点的基因型和等位基因分布频率Table 2 Genotype and allele frequency of SIRT1 and SIRT2 at different sites
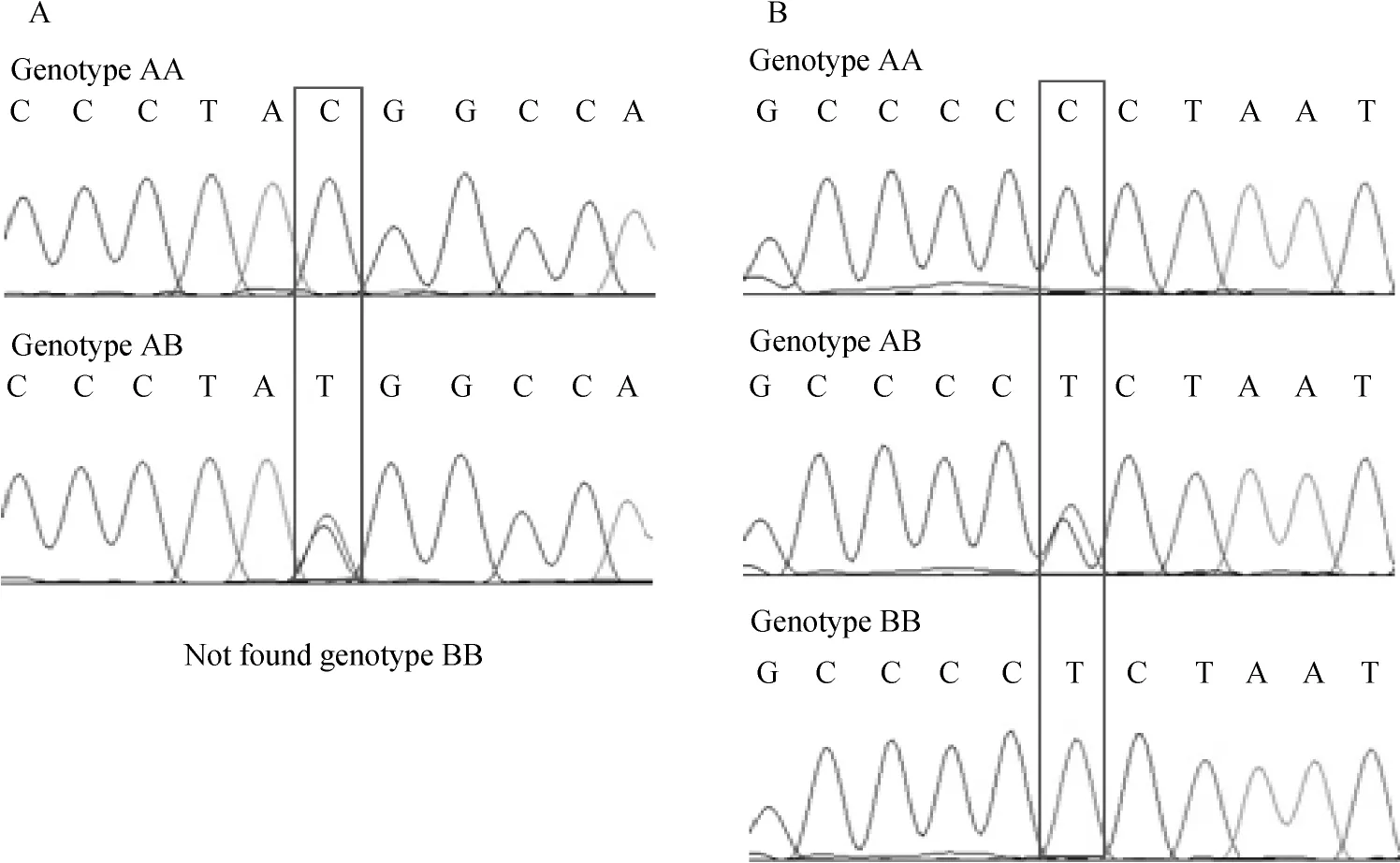
图3 SIRT2基因C19501T(A)和C19518T(B)基因型测序Fig.3 Sequencing diagram at site C19501T(A)and C19518T(B)in SIRT2 gene

图4 SIRT1基因G25764A(A)和A25846G(B)酶切Fig.4 The restriction map of G25764A(A)and A25846G(B)in SIRT1 gene
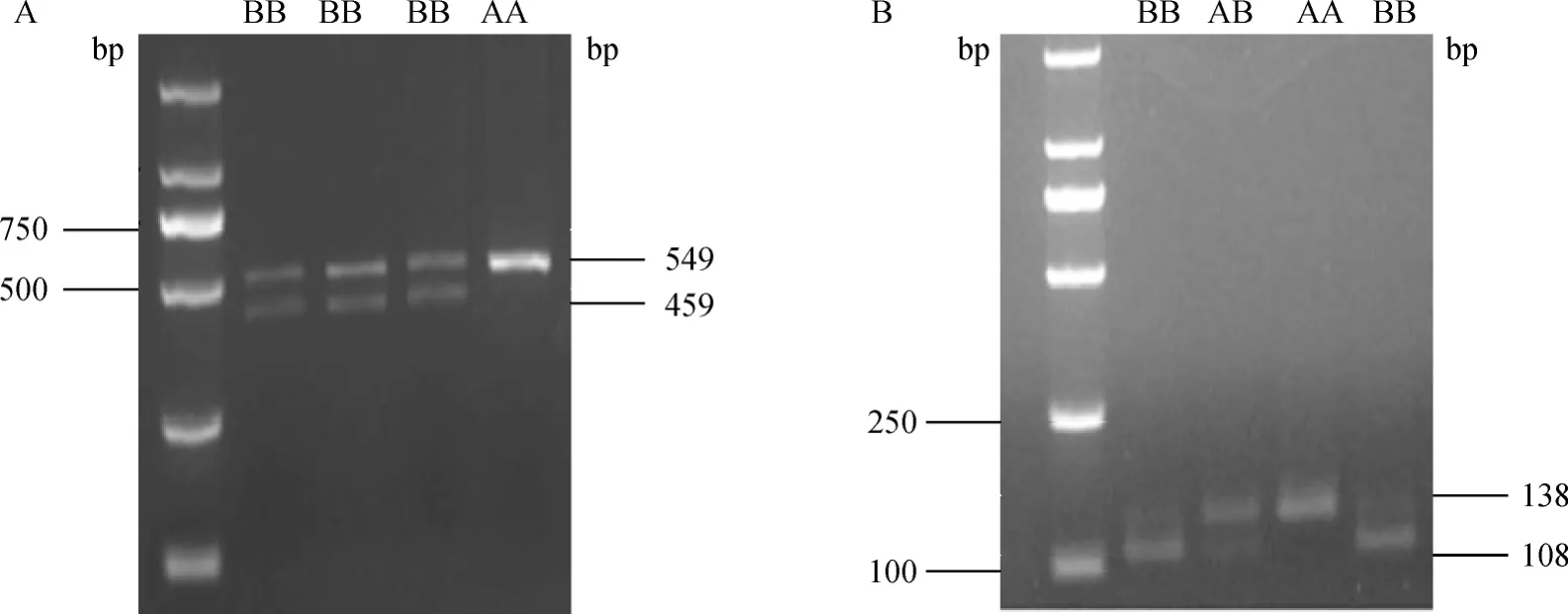
图5 SIRT2基因C19501T(A)和C19518T(B)酶切Fig.5 The restriction map of C19501T(A)and C19518T(B)in SIRT2 gene
本研究从群体遗传学角度分别分析了这4个突变位点的基因型频率、等位基因频率、杂合度、有效等位基因数和多态信息含量的遗传指标(表2),并进行卡方检验检测了Hardy-Weinberg平衡状态。结果显示,SIRT1基因G25764A和A25846G处,AA和BB分别为优势基因型,A是G25764A的优势等位基因,而B则是A25846G的优势等位基因;SIRT2基因C19501T和C19518T处,AA和AB分别为优势基因型,A同为这两个位点的优势等位基因。卡方检验表明,4个位点均处于Hardy-Weinberg平衡状态(P>0.05)。
2.3 SIRT1和SIRT2不同基因型对秦川牛肉用性状的影响
对505头健康秦川牛的3个肉用性状分别与SIRT1基因各突变位点的基因型进行关联分析。结果显示(表3),G25764A位点不同基因型在眼肌面积方面差异极显著(P<0.01),AA基因型个体均值显著高于AB型。A25846G位点上,AA基因型眼肌面积的个体均值极显著大于AB基因型均值(P<0.01),同时显著高于BB基因型(P<0.05)。对SIRT 2基因而言,C19501T位点上的AA基因型肉用性状表现最优,眼肌面积和肌间脂肪均值分别比AB基因型均值高4.09 cm2和0.31%,差异显著(P<0.05)。C19518T位点上的BB基因型为优良基因型,在背膘厚方面,BB基因型显著高于AA基因型(P<0.05);AB基因型个体的肌间脂肪含量显著大于AA基因型(P<0.05),但与BB基因型差异不显著(P>0.05)。
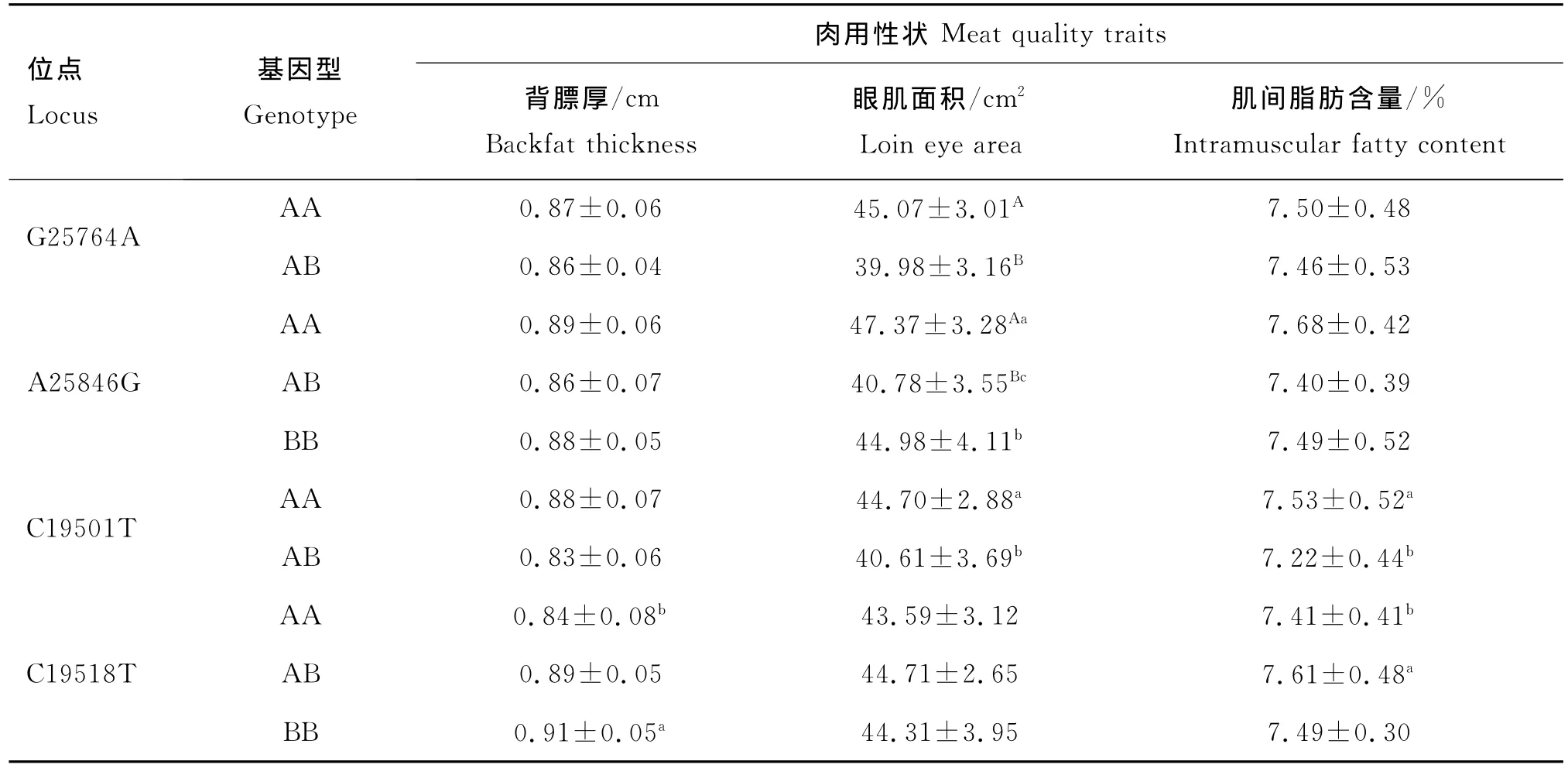
表3 SIRT1和SIRT2不同基因型对秦川牛肉用性状的影响Table 3 Effect of different genotypes at SIRT1 and SIRT2 sites on meat quality traits in Qinchuan cattle
2.4 SIRT1和SIRT2基因合并基因型对秦川牛肉用性状的影响
经分析得出,两个基因中4个位点共有22种合并基因型,将发生频率低于5.00%的13种合并基因型定义为其他类型。表4显示,AA-BB-AA-AB发生频率最高,达到25.34%,其次为AA-BB-AAAA和AA-BB-AA-BB,频率分别为21.98%和16.63%。利用合并基因型方差分析模型分析不同合并基因型对秦川牛肉用性状的影响,结果显示,合并基因型对背膘厚、眼肌面积以及肌间脂肪影响显著。由表5可见,合并基因型AB-BB-AA-BB个体的背膘厚极显著高于AA-BB-AA-AA、AA-BB-AABB和AB-AB-AA-AB3个合并基因型(P<0.01),并且显著高于AA-BB-AA-AB合并基因型个体(P<0.05)。尽管AA-BB-AA-AB合并基因型个体的眼肌面积大于AB-BB-AA-BB合并基因型个体,但差异不显著(P>0.05),这两组合并基因型均极显著高于AB-AB-AA-AB合并基因型(P<0.01)。肌间脂肪含量方面,合并基因型AB-BB-AA-BB个体均值极显著高于AA-BB-AA-AA和AB-AB-AA-AB两组合并基因型(P<0.01),并与合并基因型AA-BB-AA-BB之间存在显著性差异(P<0.05)。因此,在本试验群体中,AB-BB-AA-BB可被视为一种最为优秀的合并基因型。
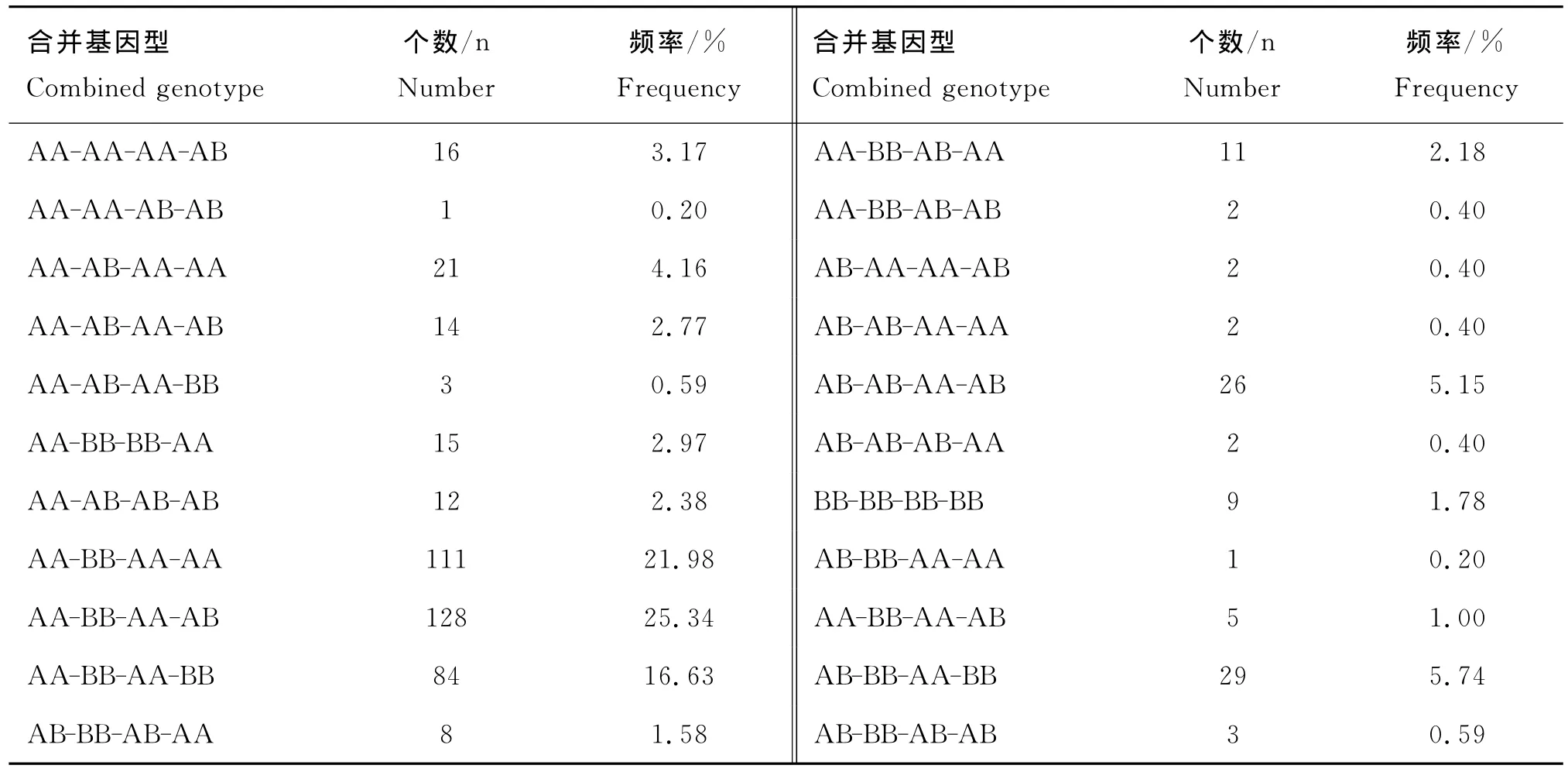
表4 秦川牛SIRT1和SIRT2合并基因型分布情况Table 4 The distribution of combined genotypes at SIRT1 and SIRT2 loci in Qinchuan cattle
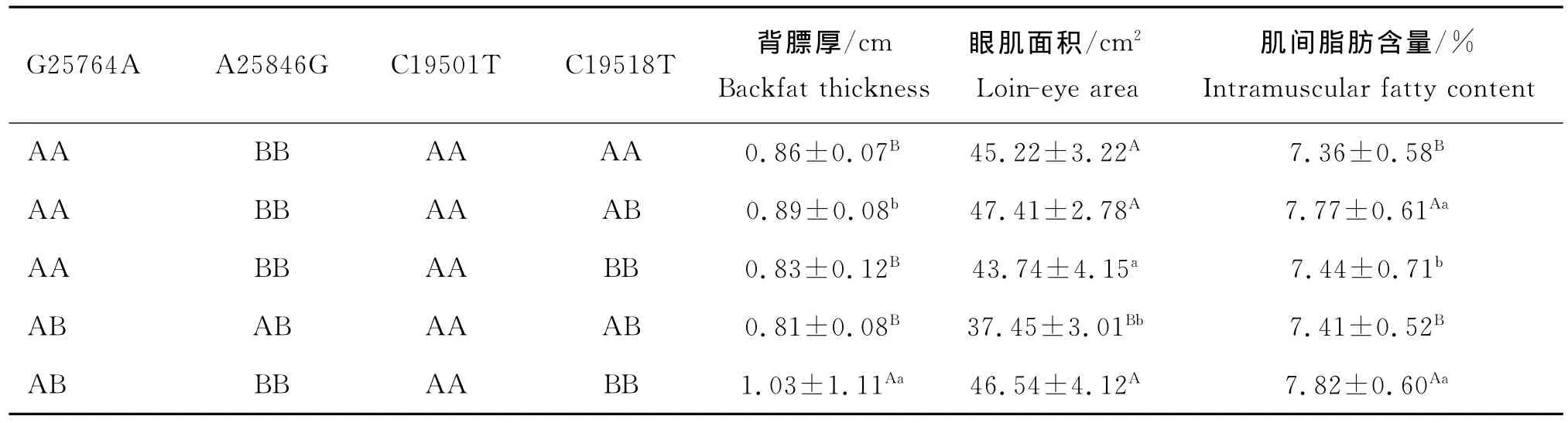
表5 SIRT1-SIRT2合并基因型对秦川牛肉用性状的影响Table 5 Effect of different combined genotypes of SIRT1-SIRT2 on meat quality traits in Qinchuan cattle
3 讨 论
目前,动物基因聚合育种技术逐步在畜禽生产中得到应用。李国辉等在白耳鸡群体中对GH、POU1F1和PRL基因的单基因及聚合基因型对产蛋率影响的研究,结果表明2个或3个优势基因聚合的遗传效应高于优势单基因型的遗传效应[15],类似的结果在张学余的研究中得到了证实[16]。郭云雁等在猪上发现,MyoG和c-fos基因的有利合并基因型和不利合并基因型个体的各性状(背膘厚除外)间差异均达到了显著或极显著水平[17]。在反刍动物方面,合并MC4R和PROP 1基因型对中国美利奴羊生长性状有一定的影响[18]。以上研究我们可以发现,在动物上实现多基因聚合育种是切实可行的,将更有利于利用标记辅助选择对畜禽进行遗传改良。
秦川牛是我国著名的五大地方黄牛之一,具有肉役性能突出、遗传稳定和适应性强等优良特性,然而其生长发育较慢,后躯肌肉欠充实的缺陷也比较明显[19]。目前,针对秦川牛的遗传改良多集中于单一基因的多态性研究[20-22],鉴于基因聚合育种技术在畜禽上的逐步应用,本试验挑选两个与机体脂肪代谢相关联的基因,研究单基因多态性及合并基因型与秦川牛肉用性状的关联分析。
SIRT 1基因在机体内可以通过对几种调控代谢及内分泌信号的转录因子去乙酰化作用来调节其活性,在调控糖脂代谢、胰岛素分泌和基因转录方面发挥重要的作用[23]。目前,在人和小鼠上研究的比较全面。M.X.Li等研究表明,SIRT1基因-274C>G突变与24月龄南阳牛的体尺性状显著相关[24-25]。本试验通过PCR-PFLP对秦川牛SIRT 1基因全长进行了多态性分析,结果检测到了在3′侧翼区发现2个多态位点,分别是G25764A和A25846G。通过不同基因型与秦川牛肉用性状关联分析发现,G25764A位点不同基因型在眼肌面积方面差异极显著(P<0.01),AA基因型个体均值显著高于AB型。A25846G位点上不同基因型同样在眼肌面积方面存在显著性差异,AA基因型个体均值极显著高于AB基因型个体均值(P<0.01),并显著高于BB基因型个体均值(P<0.05)。本试验发现的SNP位点与M.X.Li等的结果不相一致,这可能是由于样本选择上的差异所导致的。
尽管SIRT 2基因与SIRT 1基因的细胞定位不同,但作用机制大致相似,均可以通过结合并去乙酰化多种转录因子,进而调控糖脂代谢[26]。目前,尚未见其他有关SIRT2基因在动物上的多态性研究。本试验在SIRT 2基因的3′侧翼区发现2个多态位点,发现这两个突变位点均对秦川牛肉用性状有一定的影响。其中C19501T位点上的AA基因型和C19518T位点上的BB基因型为优势基因型。眼肌面积和肌间脂肪方面,C19501T的AA基因型个体均值均显著高于AB基因型(P<0.05);C19518T的BB基因型个体背膘厚均值显著高于AA基因型(P<0.05);AB基因型个体的肌间脂肪显著大于AA基因型(P<0.05),但与BB基因型差异不显著(P>0.05)。
通过合并SIRT 1和SIRT 2基因发现,合并基因型各组之间的背膘厚,眼肌面积和肌间脂肪之间差异显著(P<0.05或P<0.01)。其中合并基因型AB-BB-AA-BB个体的背膘厚和肌间脂肪含量均值最高,尽管在眼肌面积方面略小于AA-BB-AA-AB合并基因型个体的均值,但差异不显著(P>0.05)。通过比较发现,两个基因的联合效应要大于单基因对秦川牛肉用性状的效应,合并基因型AB-BB-AABB为最优秀的合并基因型。如果单纯的叠加各位点的优势基因,合并基因型AA-BB-AA-BB所表现出来的肉用性状应优于合并基因型AB-BB-AABB,这与本试验结果并不相符,说明了多基因聚合效应绝不仅仅是各基因效应的简单相加,联合效应可能优于单个基因型效应。
4 结 论
本研究发现,SIRT1(G25764A和A25846G)和SIRT2基因(C19501T和C19518T)在秦川牛中存在多态性,与部分肉用性状显著关联。SIRT1和SIRT2基因的优势合并基因型为AB-BB-AA-BB,其个体的背膘厚、眼肌面积和肌间脂肪含量极显著或显著高于其他合并基因型,可以尝试将SIRT 1和SIRT2基因作为影响生长性状的候选基因用于标记辅助选择,为秦川牛选育工作提供科学依据。
(
):
[1] LIANG F,KUME S,KOYA D.SIRT1 and insulin resistance[J].Nat Rev Endocrinol,2009,5(7):367-373.
[2] NORTH B J,VERDIN E.SIRTuins:Sir2-related NAD-dependent protein deacetylases[J].Genome Biol,2004,5(5):224.
[3] TONTONOZ P,SPIEGELMAN B M.Fat and beyond:the diverse biology of PPARγ[J].Annu Rev Biochem,2008,77:289-312.
[4] PICARD F,KURTEV M,CHUNG N,et al.Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma[J].Nature,2004,429(6993):771-776.
[5] BAUR J A,PEARSON K J,PRICE N L,et al.Resveratrol improves health and survival of mice on a high-calorie diet[J].Nature,2006,444(7117):337-342.
[6] LAGOUGE M,ARGMANN C,GERHART-HINES Z,et al.Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α[J].Cell,2006,127(6):1109-1122.
[7] MILNE J C,LAMBERT P D,SCHENK S,et al.Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes[J].Nature,2007,450(7170):712-716.
[8] GAIMAN S,ARMSTRONG C M,KAEBERLEIN M,et al.Transcriptional silencing and longevity pro-tein Sir2 is an NADdependent histone deacetylase[J].Nature,2000,403(6771):795-800.
[9] LANDRY J,SUTTON A,TAFROV S T,et al.The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases[J].Proc Natl Acad Sci USA,2000,97(11):5807-5811.
[10] JING E,GESTA S,KAHN C R.SIRT2 regulates adipocyte differentiation through FoxO1 acetylation/deacetylation[J].Cell Metab,2007,6(2):105-114.
[11] WANG F,TONG Q.SIRT2 suppresses adipocyte differentiation by deacetylating FOXO1 and enhancing FOXO1’s repressive interaction with PPARgamma[J].Mol Biol Cell,2009,20(3):801-808.
[12] 刘炳婷,白 亮,刘 飞,等.过表达SIRT2抑制猪前体脂肪细胞的分化[J].中国生物化学与分子与分子生物学报,2010,26(6):575-580.LIU B T,BAI L,LIU F,et al.Overexpression of Sirt2 suppresses porcine preadipocyte differentiation[J].Chinese Journal of Biochemistry and Molecular Biology,2010,26(6):575-580.(in Chinese)
[13] J萨姆布鲁克,DW拉塞尔.分子克隆实验指南(上、下册)[M].黄培堂等译.北京:科学出版社,2002.SAMBROOK J,RUSSELL D W.Molecular cloning a laboratory manual[M].HUANG P T.Beijing:Science Press,2002.(in Chinese)
[14] WEAVER B,DUBOIS S.SPSS macros to compare any two fitted values from a regression model[J].Behav Res Methods,2012,44(4):1175-1190.
[15] 李国辉,张学余,苏一军,等.3个基因SNPs及基因聚合对白耳鸡产蛋数的遗传效应[J].江西农业大学学报,2010,32(4):661-667.LI G H,ZHANG X Y,SU Y J,et al.Effect of single and pyramiding genotypes of three genes on egg production of Baier chicken[J].Acta Agriculturae Universitatis Jiangxiensis,2010,32(4):661-667.(in Chinese)
[16] 张学余,韩 威李国辉.AFABP和HFABP基因多态位点及聚合基因型对白耳鸡胸肌肌内脂肪含量(IMF)的效应分析[J].云南农业大学学报,2010,25(4):530-534.ZHANG X Y,HAN W,LI G H.Effect of polymorphism sites and pyramiding genotypes of A-FABP and H-FABP genes on Baier chicken muscle intramuscular fat(IMF)contents[J].Journal of Yunnan Agricultural University,2010,25(4):530-534.(in Chinese)
[17] 郭云雁,程笃学,张龙超,等.MyoG和c-fos基因多态性及其合并基因型与猪肌纤维性状的关联分析[J].畜牧兽医学报,2013,44(2):188-196.GUO Y Y,CHENG D X,ZHANG L C,et al.Effects of single and combined genotypes of MyoG and c-fos genes on muscle fibers traits in pig[J].Acta Veterinaria et Zootechnica Sinica,2013,44(2):188-196.(in Chinese)
[18] 曾献存,陈韩英,贾 斌,等.MC4R和PROP 1基因多态性及合并基因型与中国美利奴羊生长性状的关联分析[J].畜牧兽医学报,2011,42(9):1227-1232.ZENG X C,CHEN H Y,JIA B,et al.Effects of single and combined genotypes of MC4R and PROP1 genes on growth traits in Chinese Merino sheep[J].Acta Veterinaria et Zootechnica Sinica,2011,42(9):1227-1232.(in Chinese)
[19] 高建斌,昝林森,杨 宁,等.秦川牛DKK1基因SNPs检测及其与体尺、肉质性状的关联分[J].畜牧兽医学报,2013,44(3):376-386.GAO J B,ZAN L S,YANG N,et al.Polymorphisms of DKK1 gene and its association with body measurement and meat quality traits in Qinchuan cattle[J].Acta Veterinaria et Zootechnica Sinica,2013,44(3):376-386.(in Chinese)
[20] 杨彦杰,昝林森,王洪宝.秦川牛脂联素基因SNPs检测及其与胴体、肉质性状的相关性[J].遗传,2009,31(10):1006-1012.YANG Y J,ZAN L S,WANG H B.SNPs detection of adiponectin gene and its relationship with carcass and meat quality traits in Qinchuan cattle[J].Hereditas,2009,31(10):1006-1012.(in Chinese)
[21] TIAN W Q,WANG H C,SONG F B,et al.Association between a single nucleotide polymorphism in the bovine chemerin gene and carcass traits in Qinchuan cattle[J].Genet Mol Res,2011,10(4):2833-2840.
[22] XUE M,ZAN L S,GAO L,et al.A novel polymorphism of the myogenin gene is associated with body measurement traits in native Chinese breeds[J].Genet Mol Res,2011,10(4):2721-2728.
[23] ANNE B,LORA B S,FITZHUGH S,et al.Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase[J].Science,2004,303(26):2011-2015.
[24] LI M X,SUN X M,HUA L S,et al.SIRT1 gene polymorphisms are associated with growth traits in Nanyang cattle[J].Mol Cell Probes,2013,27:215-220.
[25] LI M X,SUN X M,ZHANG L Z,et al.A novel c.-274C>G polymorphism in bovine SIRT1 gene contributes to diminished promoter activity and is associated with increased body size[J].Anim Genet,2013,44(5):584-587.
[26] WANG F,NGUYEN M,QIN F X,et al.SIRT2 deacetylates Fox O3a in response to oxidative stress and caloric restriction[J].Aging Cell,2007,6(4):505-514.
猜你喜欢
畜牧兽医学报(2022年12期)2023-01-03 02:28:26
中国牛业科学(2022年2期)2022-12-08 07:51:41
现代畜牧科技(2021年5期)2021-07-20 08:07:22
养猪(2020年2期)2020-04-14 11:42:50
中国牛业科学(2020年3期)2020-01-11 16:06:25
养猪(2019年3期)2019-06-19 06:53:14
中国畜牧业(2015年17期)2015-12-07 08:30:50
当代畜禽养殖业(2014年12期)2014-02-27 08:00:07
当代畜禽养殖业(2014年7期)2014-02-27 07:59:13
中国牛业科学(2011年6期)2011-08-04 03:02:54
