
新疆托木尔峰国家级自然保护区树附生地衣群落数量分类1)
2015-03-06 09:16艾尼瓦尔吐米尔阿不都拉阿巴斯
东北林业大学学报 2015年5期
艾尼瓦尔·吐米尔 阿不都拉·阿巴斯
(新疆大学,乌鲁木齐,830046)
责任编辑:王广建。
地衣作为生态系统的重要组成成分之一,在植物群落演替过程中,地衣对微环境的改变,为随后定居的植物提供栖息地,给食草动物提供食物等方面具有重要的作用。同时,地衣体内的共生光合蓝藻促进生态系统的氮循环,加速生态系统的物质和养分的循环[1-2]。附生地衣是森林生态系统生物多样性的重要成员,广泛的分布在各类森林生态系统。附生地衣具有提高森林结构复杂性、改变林冠层水分、影响营养物质的循环以及为动物提供栖息地、食物和筑巢材料等功能[3-5]。附生地衣的生态学研究是目前国外比较受关注的研究领域之一,在森林生态系统中,附生地衣对生态系统中物质、能量的平衡以及生物多样性的维系等方面发挥着重要的作用。
在国外,附生地衣生态学研究起步较早,他们采用了数值分类等生态学定量研究方法,系统的研究了不同类型森林生态系统中的附生地衣群落结构特征、地衣群落物种分布格局以及影响地衣物种分布的环境因素等[6]。Cobanoglu等[7]研究了土耳其安塔利亚Elmali雪松林附生地衣物种组成、种类分布与树木直径大小、海拔高度、方位、方向等之间的关系。Ozturk等[2]采用除趋势对应分析法(DCA)和双向指示种分析法(TWINSPAN)等定量方法,对土耳其Uludag山脉附生地衣在森林生态系统中的地带性分布特征进行了研究。Nascimbene等[8]研究了意大利东北高山银杉林森林生态系统的动态演替与附生地衣群落间的关系,发现该地区附生地衣种类及群落物种组成随着森林结构和动态而发生变化。
在国内,虽然地衣群落生态学方面研究起步较晚,但近年来研究人员应用定量研究方法研究了不同地区森林生态系统附生地衣群落及物种多样性。陈健斌等[9]对北京东灵山地区主要树生地衣种类的组成进行了报道;Li等[10-11]在我国西南地区的哀牢山原生林和次生林中对附生大型地衣多样性、物种组成和生物量进行研究。郗鹏等[12]对北京东灵山小龙门林场6种树木树皮上生长的地衣的多样性和生态特点进行了调查研究;王玉良等[13]对鹞落坪国家级自然保护区森林树生地衣群落进行了定量研究,并分析了群落中地衣种间关联关系;艾尼瓦尔·吐米尔等[14-16]对新疆阿勒泰东部山脉、乌鲁木齐南部山区森林生态系统和阿尔泰山两河源自然保护区森林生态系统附生地衣群落进行了定量研究。
王先业[17]首次报道天山托木尔峰地区的地衣的文献,文中报道天山托木尔峰地区的地衣有67种10变种和4变型,隶属于28属,15科,其中23种3变种3变型为我国新记录,28种5变种1变型为新疆新分布,并有两个新组合。然后,有关学者根据实地调查和文献资料对托木尔峰自然保护区地衣的地理区系进行了补充报道[18-19]。但对托木尔峰国家级自然保护区地衣群落生态学方面尚未见报道。因此,系统地研究该地区树附生地衣物种多样性及地衣群落特征,不仅为该地区生物多样性的有效管理提供基础数据,同时也对科学评价托木尔峰国家级自然保护区森林生态系统的稳定性和环境质量提供科学依据。
1 研究地区概况
托木尔峰自然保护区位于我国新疆天山山脉西端温宿县境内的托木尔峰地区。托木尔峰是天山山脉最高峰,海拔7 435.3 m。它位于天山山脉的中段,面积约1.5×104km2。气候属于温带南缘,由于本地区的大气环流和天山的屏障作用,致使山地南、北坡的气候迥然不同,托木尔峰北坡属于温带大陆性半湿润型气候,南坡为温带大陆性半干旱型气候。年降水量在两坡差别较大(南坡约300 mm,北坡600~700 mm)。由于地质、地形条件复杂,南、北坡气候差异显著,水热条件的垂直变化明显,对植被类型多样性的形成创造了极有利的条件,温带荒漠、山地草原和森林的广泛分布是本地区自然景观的主要特征[20]。
2 研究方法
2.1 野外调查
在托木尔峰国家级自然保护区的北坡、海拔2 200~3 000 m随机设立20 m×20 m的样地25个。每个样地选择2个样点,每个样点选择胸径(DBH)大于30 cm的树木。在每棵树的胸径高度,用面积为20 cm×20 cm的铁筛调查附生地衣种类、数量、频度、盖度等;同时记录每个样点的树种、森林郁闭度等。
2.2 数据分析
以样点为对象,以树附生地衣的盖度为指标,以地衣种数和样点数构成矩阵,采用双向指示种分析方法和除趋势对应分析法,对附生地衣群落分类及不同海拔的梯度分布分析[2,10-11]。数据分析采用Win TWINS 2.3和CANOCO for Windows 4.5软件。
根据树附生地衣的种类和覆盖度,计算样点组的Shannon-Weiner物种多样性指数和Pielou均匀度指数,比较各样点组的物种多样性特点。
Patrick丰富度指数:D=S。D表示丰富度指数;S表示样地内的种数。
采用Shannon-Weiner公式计算多样性指数[21]。公式为
均匀度用以下公式来计算:J=H/Hmax。
式中:Pi=ni/N,N为样方内地衣覆盖度,ni为第i种的覆盖度。J为均匀度指数;H为实测多样性值指数;Hmax为理论上群落多样性指数的最大值(Hmax=ln s,s为总种数)。
群落Whittaker相似性指数计算公式[21]:
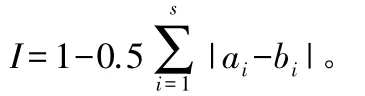
式中:I为相似性指数;ai、bi分别为物种i个体在群落a和群落b中的比例;s为a和b群落中相应的种数。
3 结果与分析
3.1 树附生地衣种类组成
根据已有的文献资料[17-18,22]并结合实地调查数据,本研究共鉴定出分布在托木尔峰国家级自然保护区北坡的树附生地衣有31种隶属于14科、20属(见表1)。从表1可知,茶渍目在研究地区占优势,分别占该地区附生地衣科、属和种数的85.71%、85%和87.09%。
3.2 树附生地衣群落数值分类
本研究中,分布在50个样点树的31个树附生地衣种类,根据地衣的盖度进行了定量分析。对树附生地衣种类和样点的DCA排序(见图1)。
DCA排序的前两个排序轴的特征值分别为0.87(第一排序轴)和0.643(第二排序轴),包含了48.42%的信息量,符合进行DCA排序分析。DCA排序结果表明,附生地衣种类在不同海拔分布形成4个组(见图1)。采集海拔2 200~2 600 m的树附生地衣种类分布在DCA排序图的左侧,其中海拔2 200 m的10个样点组成一个组;海拔2 400和2 600 m的20个样点组成第二组。海拔2 800和3 000 m的样点在DCA排序图的右边形成两个组。根据树附生地衣的覆盖度作为指标将50个样点进行TWINSPAN数量分类,结果见图2。

表1 托木尔峰自然保护区树附生地衣种类组成
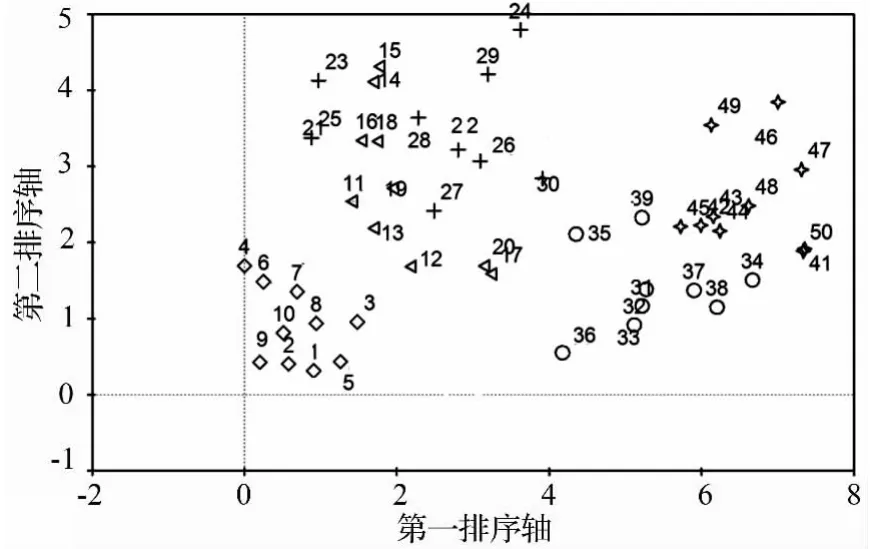
图1 树附生地衣盖度在不同海拔的DCA排序
依据DCA和TWINSPAN分析结果,并结合研究地区的自然环境特征,将研究地区的树附生地衣分类为4个群落,并根据群落生境特征的指示种和优势种命名群落[2]。
群落A:包括样点1~10,分布在海拔2 200 m左右,降水量适中,光照条件较好。主要的树种分布在水分较好的河谷两侧,以绒毛杨(Populus pilosa Rehd.)、银柳(Salix argyracea E.Wolf)和细穗柳(Salix tenuijulis Ledeb.)等组成,郁闭度小于0.4。主要地衣种类有茎口果粉衣(Ch.stemonea(Ach.)Muell)、枪石蕊(Cl.coniocraea(Floerke)Spreng.)、小疣巨孢衣(M.verrucosa(Ach.)Haf.)、黑蜈蚣衣(Ph.nigricans(Flk.)Moberg)、类锈橙衣(C.ferrugineoides(H.Magn))、柳茶渍(L.saligna(Schrad.)Zahlbr.)等6种。命名为:茎口果粉衣+黑蜈蚣衣+枪石蕊群落。
群落B:包括海拔2 400~2 600 m的样点11~30。海拔较低处的树种有帕米尔杨(Populus pamirica Kom.)、天山桦木(Betula tianschanica Rupr.),林下灌木有刚毛忍冬(Lonicera hispida Pall.ex Roem.et Schult.)、天山花楸(Sorbus tianschanica Rupr.)等,在海拔较高处还分布着雪岭云杉(Picea schrendina Fisch.et Mey),郁闭度0.5左右。分布在该植被带的主要树附生地衣种类有柯贝尔副茶渍(L.koerberiana Lahm.)、黄烛衣(C.concolor(Dicks.)Stein.)、包氏饼干衣(R.bohlinii H.Magn.)、茸刺梅衣(P.subargentifera Nyl.)、蜈蚣衣(Ph.stellaris(L.)Nyl.)、粉粒树花(R.pollinaria(Westr.)Ach.)、中国树花(R.sinensis Jatta.)、白蜡树花(R.fraxinea(L.)Ach.)等15种。命名为:黄烛衣+粉粒树花+中国树花群落。
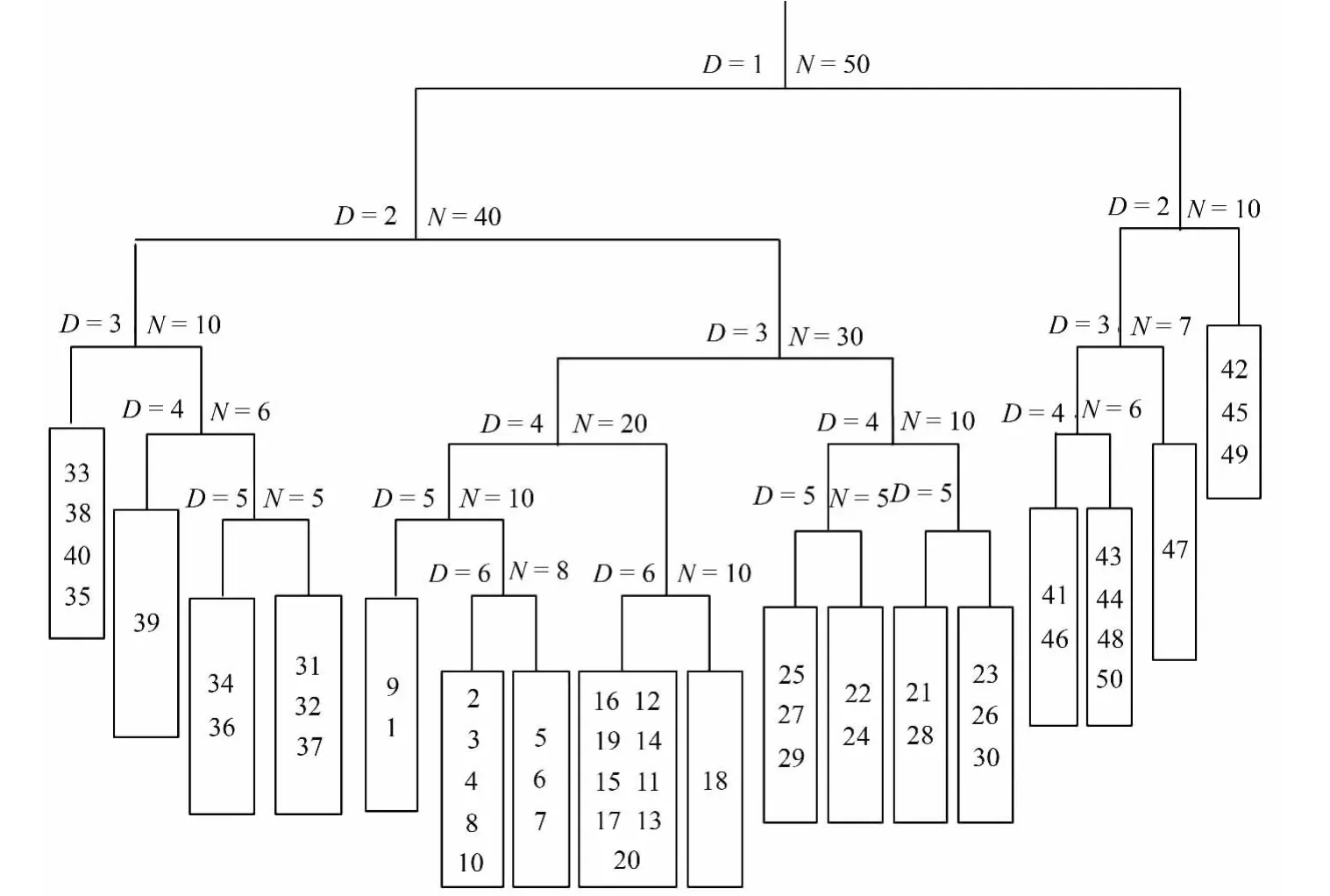
图2 树附生地衣群落TWINSPAN分类树状图
群落C:包括样点31至样点40,分布在海拔2 800 m。该地带的树种有雪岭云杉(Picea schrendina),郁闭度达到0.8左右,雨量充足,地表凋落物较丰富。主要地衣种类有柯贝尔副茶渍(L.koerberiana Lahm.)、金黄茶渍(C.aurella(Hoffm.)Zahlbr.)、粗糙猫耳衣(L.brebisonii Mont.apud Webb.)、包氏饼干衣(R.bohlinii H.Magn.)等10种。命名为:柯贝尔副茶渍+金黄茶渍+粗糙猫耳衣群落。
群落D:包括样点41至50,分布海拔2 850 m至3 000 m左右。树种有雪岭云杉(Picea schrendina),郁闭度0.5左右。主要地衣种类有短黄枝衣(T.brevior(Nyl.)Vain.)、类锈橙衣(C.ferrugineoides(H.Magn))、斑面蜈蚣衣(Ph.aipolia(Humb.)Furnr.)、小疣巨孢衣(M.verrucosa(Ach.)Haf.)、金黄茶渍(C.aurella(Hoffm.)Zahlbr.)等8种。命名为:斑面蜈蚣衣+类锈橙衣+污白雪花衣群落。
3.3 树附生地衣群落参数
为了进一步找出各群落之间的差异,我们计算群落多样性指数、均匀度指数和相似性系数,比较了4个不同的群落之间的相关性(见表2、表3)。
群落的多样性通常与其组成种的丰富度或多度及种间个体数分布的均匀度两个参数有关。用其指数可以定量比较不同地区或同一地区不同群落的结构特征。由表2可见,群落B的多样性指数最大(2.251);其次为群落C(1.725),群落A的多样性指数最低(1.017)。
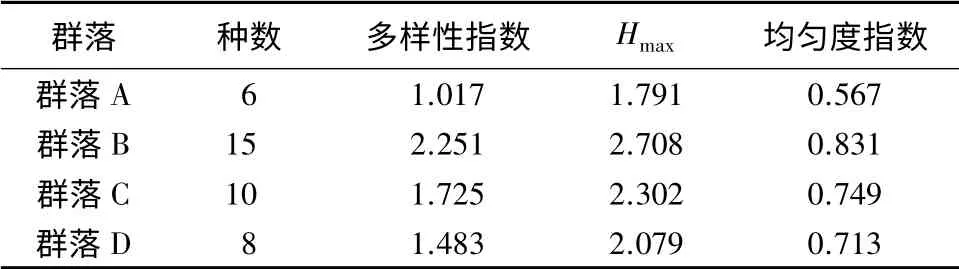
表2 托木尔峰自然保护区森林生态系统树生地衣群落多样性及均匀度

表3 托木尔峰自然保护区森林生态系统树生地衣群落相似性
群落相似性是反映不同群落相互关系程度的主要参数,用该参数可以比较不同地区或同一个地区不同群落之间的差异程度,为群落的分类提供依据。由表3可知,群落B和群落C之间的相似性最高,相似性系数为0.918,这与群落B和群落C中分布的树生地衣种类比较相似,主要树种基本一致等因素有关系;群落C和群落D相似性次之,相似性系数为0.649;群落A和群落D之间的相似性最低,相似性系数为0.264,其主要原因是组成群落的地衣种类不同,森林郁闭度、光照强度等因素有一定的关系。
4 结论与讨论
树附生地衣是森林生态系统的主要成分之一,保护树附生地衣物种多样性,在有效评价森林生态系统稳定性和环境质量方面具有重要意义。已有的研究表明,树附生地衣群落物种组成及分布受多种环境因素的影响[1,6,23]。Hauck[5]根据文献资料,对影响北方针叶林树附生地衣丰富的因素进行了系统的分析,并认为针叶林生态系统树附生地衣的分布,主要受光照、温度、水分、树皮酸碱度、养分、空气污染、林龄、树木连续性,以及附载植物胸径、树高、树种因素的控制。Ozturk等[2]采用DCA和TWINSPAN研究方法,对土耳其Uludag山脉附生地衣在森林生态系统中的地带性分布研究时发现,温度、降水量和湿度等气候因素与海拔高度的关系密切,地衣生物量和湿度具有正相关关系。因此,树附生地衣群落的物种组成随着海拔梯度的变化,充分的反映了生态条件(除了森林性质和年龄外)对地衣群落组成及物种分布的影响[1-2,6]。Mc Mullin等[4]采用CCA排序法,研究了加拿大西南部诺瓦斯克舍省针叶林森林的结构复杂性和附生地衣多样性间的关系,发现森林结构越复杂,不同年龄的树种越多,可以有效地提高森林生态系统内部环境的异质性,从而提高了附生地衣的多样性。因此,提出在森林砍伐时,应该充分考虑森林生态系统中不同年龄阶段的树木的保护,适当留下年龄大的树木,通过调整林冠层的郁闭度、增加不同年龄树种的数量,能够提高森林生态系统树附生地衣植物的多样性。艾尼瓦尔·吐米尔等[16]采用CCA排序方法,对阿尔泰山两河源自然保护区树附生地衣种类的分布与环境因素之间的关系进行了定量研究,发现研究地区树附生地衣群落的分布主要受到海拔、人为干扰、郁闭度、空气湿度和光照强度等因素的影响。其中,对于地衣分布影响最大的是海拔高度,其次为郁闭度和湿度。
采用双向指示种分析方法和除趋势对应分析法,初步研究了托木尔峰自然保护区不同海拔高度的树附生地衣物种多样性及群落结构。结果发现:研究地区森林生态系统中随着海拔高度的升高,分布在树干和树枝上的地衣种类出现差异,充分说明了各种生态因子随着海拔的变化,影响树附生地衣种类的分布。同时,采用多元数量分析,结合传统的群落分类方法对地衣植物群落进行分类,能够有效地提高研究数据的科学性、准确性,减少研究者的偏见和避免数据处理当中的一些误差。
因此,系统的研究托木尔峰国家级自然保护区森林生态系统树生地衣群落结构及物种多样性,不仅为掌握该地区森林生态系统结构特征、植物群落结构、植物资源分布规律等提供基础资料,而且为该区制定有效的可持续森林经营决策提供科学依据。
[1]Romagni J G,Gries C.Post fire recolonization of dominant epiphytic lichen species on Quercus hypoleucoides(Fagaceae)[J].American Journal of Botany,2000,87(12):1815-1820.
[2]Ozturk S,Oran S,Guvenc S,et al.Analysis of the distribution of epiphytic lichens in the oriental beech(Lipsky)forests along an altitudinal gradients in Uludag mountain,Bursa-Turkey[J].Pakistan Journal of Botany,2010,42(4):2661-2670.
[3]Dettki H,Esseen P A.Modelling long-term effects of forest management on epiphytic lichens in northern Sweden[J].Forest Ecology and Management,2003,175(1/3):223-238.
[4]McMullin R T,Duinker P N,Richardson D.Relationships between the structural complexity and lichen community in coniferous forest of southwestern Nova Scotia[J].Forest Ecology and Management,2010,260(5):744-749.
[5]Hauck M.Site factors controlling epiphytic lichen abundance in northern coniferous forests[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2011,206(2):81-90.
[6]Christopher J E.Lichen epiphyte diversity:A species,community and trait-based review[J].Perspectives in Plant Ecology,Evolution and Systematics,2012,14(2):131-152.
[7]Cobanoglu G,Sevgi O.Analysis of the distribution of epiphytic lichens on Cedrus libani in Elmali research forest(Antalya,Turkey)[J].Journal of Environmental Biology,2009,30(2):205-212.
[8]Nascimbene J,Dainese M,Sitzia T.Contrasting response of epiphytic and dead wood dwelling lichen diversity to forest management abandonment in silver fir mature woodland[J].Forest Ecology and Management,2013,289:325-332.
[9]陈健斌,刘晓娟,黄永青.北京东灵山地区主要树生地衣调查初报[J].生态学报,1999,19(1):76-79.
[10]Li S,Wen Y L,Da W L.Bole epiphytic lichens as potential indicators of environmental change in subtropical forest ecosystemin sounthern China[J].Ecological Indicators,2013,29:93-104.
[11]Li S,Wen Y L,Da W L.Epiphytic lichens in subtropical forest ecosystem in southern China:Species diversity and implications for conservation[J].Biological Conservation,2013,159:88-95.
[12]郗鹏,杨丽媛,陈赟,等.北京东灵山小龙门地区树生地衣多样性及生态研究[J].北京师范大学学报:自然科学版,2008,44(5):507-510.
[13]王玉良,郑爱琴,阿不都拉·阿巴斯.鹞落坪树生地衣群落数量分类及多样性[J].广西植物,2011,31(6):747-753.
[14]艾尼瓦尔·吐米尔,阿不都拉·阿巴斯.阿勒泰东部山区森林生态系统树生地衣群落特征的初步研究[J].云南植物研究,2006,28(4):415-420.
[15]艾尼瓦尔·吐米尔,热依木·马木提,阿不都拉·阿巴斯.乌鲁木齐南部山区森林生态系统树生地衣群落结构[J].植物学报,2009,44(5):578-586.
[16]艾尼瓦尔·吐米尔,张婷,阿不都拉·阿巴斯.新疆阿尔泰山两河源国家级自然保护区树附生地衣群落物种分布与环境关系的研究[J].林业资源管理,2013(2):56-63.
[17]王先业.天山托木尔峰的地衣[M]//中国科学院登山科学考察队.天山托木尔峰的生物.乌鲁木齐:新疆人民出版社,1985:328-353.
[18]艾尼瓦尔·吐米尔,阿不都拉·阿巴斯.托木尔峰自然保护区地衣的补充研究[J].干旱区研究,2000,17(3):20-27.
[19]热衣木·马木提,阿地里江,阿不都拉·阿巴斯.托木尔峰自然保护区地衣的生态地理成分及垂直分布[J].东北林业大学学报,2010,38(10):44-50.
[20]中国科学院登山科学考察队.天山托木尔峰的生物[M].乌鲁木齐:新疆人民出版社,1985.
[21]阳含熙,卢泽愚.植物生态学的数量分类方法[M].北京:科学出版社,1981.
[22]阿不都拉·阿巴斯,吴继农.新疆地衣[M].乌鲁木齐:新疆科技卫生出版社,1998.
[23]Roger P C.Factors influencing epiphytic lichen communities in aspen-associated forests of the bear river range,IDAHO and UTAH[D].Logan:UTAH State University,2007.
猜你喜欢
湖北植保(2022年4期)2022-08-23
党政干部论坛(2022年3期)2022-04-20
土壤(2021年1期)2021-03-23
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
土壤学报(2017年5期)2017-11-01
摄影之友(影像视觉)(2017年1期)2017-07-18
福建农业学报(2016年6期)2016-11-01
中国环境监察(2016年5期)2016-10-24
小学阅读指南·低年级版(2016年6期)2016-05-14
