
选择性注意的神经机制和眼动注意模型的研究综述
2015-02-17 10:27张家健
重庆理工大学学报(自然科学) 2015年11期
徐 鹏,张家健,赵 冰
(河海大学商学院,南京 211100)
选择性注意的神经机制和眼动注意模型的研究综述
徐 鹏,张家健,赵 冰
(河海大学商学院,南京 211100)
基于不同标准将眼动模型进行分类,分析不同模型的研究重点,剖析了各模型的优势和存在的问题。总结以人类为被试对象的研究中有关注意的神经机制以及注意在觉知中的作用,并在已有模型的基础上设计了外显眼动-内隐注意的眼动注意模型,用以描述刺激显著性与自上而下的主动注意控制如何通过互相作用产生不同程度的大脑活动,从而产生不同的眼动和觉知状态。
选择性注意;注意的神经机制;眼动注意模型
本文主要研究选择性注意的机制。选择性注意是这样一类认知过程:它们让有机体能够加工与当前情境相关的输入、思想或行为,而忽视与当前情境无关或引人分心的刺激。本文不讨论注意的非选择性过程,如唤醒;也不讨论注意与其他认知模块的相互作用,如记忆。所以,本文的术语“注意”将指代选择性注意这一特指的概念[1]。本文立足于容量有限加工假设,即人类的信息加工系统不能同时加工高信息负荷的多重输入。在多重输入的重压下,人类的行为绩效会下降。由资源有限性造成的加工瓶颈使这个系统不得不艰难地“决策”到底选择哪些信息接受广泛加工或获得进入觉知的机会。
1 选择性注意的内涵
注意一词可能有许多内涵,它涉及意识、觉知、注意力以及相关的功能障碍。注意是指将觉知集中于一个刺激、思想或行为上,同时忽略其他不相关的刺激、思想或行为的能力[2]。注意研究是与认知心理学同步出现的。注意研究至少部分等同于对信息加工局限性的研究[3]。注意的存在表明我们没有能力加工施加在感觉器官上的一切,并对其做出反应,而且我们也不需要这种覆盖一切的加工,特别是当我们身边充斥着大量信息的时候。内隐注意(hermannvon helmholtz)指注意所指向的位置可以与注视的位置不同。实验证明:利用一种有意的意图,即使不产生眼动和眼球调节上的变化,人仍然可以将注意集中在外周神经系统的一个特定部分上,并同时将来自其他部分的注意排斥在外[4-5]。
2 注意的神经机制
脑电和脑成像结果为早期选择模型提供了数据证据,不同的实验和模型从不同的角度解释了注意的神经机制。高度特异化的系统参与到了注意控制之中。注意控制系统包括顶叶的一部分、颞叶皮质、额叶皮质和一些下皮质结构,这些区域组成了注意控制的根源[6]。注意可以在多个加工阶段中起作用,自下而上的刺激显著性与自上而下的注意作用之间的相互作用决定了哪些信息输入可以到达觉知而哪些不能[6]。
2.1 有意注意
在使用提示任务对空间有意注意的线索化研究中,当目标出现在被试所预期的位置上,被试会更快地对其做出反应;而如果目标出现在未被提示的位置上,被试的反应会变得较慢。因此可知,当线索的预测能力促使被试将内隐注意指向被提示的视野位置时,这种内隐注意可能通过影响感觉和知觉加工来影响反应时间;当被试注意某一个刺激位置的时候,知觉加工就可能发生变化。初级视皮质对神经活动进行颅内记录的结果表明,最早一批上行输入信号进入纹状皮质V1是在刺激呈现至少35 ms后,因此,P1注意效应反映的是皮质活动[7]。更多研究结果表明:最早的空间注意效应产生于纹外皮质,视觉选择性注意在视觉感觉加工的早期就在视皮质引发了稳定的变化。
2.2 反射性注意
在反射性线索化范式中,注意是被外在刺激的低水平特征控制的,而不是被内源性地有意控制。这类效应趋向于表现出空间特异性,会影响到对被提示位置及其周边区域的加工,但只存在于外源性线索之后很短的一段时间里(50~200 ms)。当任务无关的外源性刺激与目标刺激的时间间隔变大时(>300 ms),随着时间流逝,最近被反射性地注意过的位置会被抑制,被试对被提示位置上的刺激的反应会变得比较慢,产生返回抑制现象[8]。
当一个感觉事件与之后的一个目标刺激紧随出现在同一位置时,反射性线索效应就可能出现。在这种条件下,对被提示位置的反应时间短于对非提示位置的反应,但这种情况只在线索和目标刺激间的时间间隔较短(<200 ms)的时候出现;在时间间隔较长的情况下,这种效应会发生反转,产生返回抑制。Joseph Hopfinger反射性的和有意的空间注意转移都会对早期视觉加工施加相似的生理调制[1]。
2.3 视觉搜索的空间注意
在视觉搜索过程中,当一个视觉搜索任务要求有意地进行注意移动时,被试的反应要慢于视觉搜索任务不要求有意地进行注意移动的情况;当被试让自己注意的焦点被搜索集中的视觉感觉信息驱动时,视觉搜索快于执行缓慢、自主控制的项目搜索[9]。为了探索视觉空间注意的机制,Hans-jochen Heinze&G.R.M,Mangun将ERP和基于正电子发射断层扫描PET的功能性成像技术结合,实验发现空间注意会导致被注意视野的对侧半球纹外皮质激活,这种激活在腹侧皮质表面上的后部梭状回上尤为明显[1]。由此可知,P1注意效应和后部梭状回的PET注意效应是相互联系的。在于空间注意的 fMRI研究中,Roger Tootell&Anders Dale[5]考察了视皮质中注意调控的组织细节,研究发现:空间注意表现为对多重纹外视觉区中神经活动的较强调节,同时也包括较小幅度的对纹外皮质的调节。
2.4 特征注意
特征注意起到了提供信号让空间注意能够被指向含有相关特征的刺激所在位置上的作用,在这个位置上空间注意聚焦并执行更细节化的分析。Marissa Carrasco指出:提前提供指向刺激的某个视觉特征的注意线索会导致任务表现提升。注意既可以被指向某个空间位置,又可以被指向目标刺激的某个非空间的特征。
Steve Luck&Steve Hillyard发现:N2pc是内隐视觉注意聚焦的一个标志,它反映了加工过程中完成客体识别前的一个阶段。Max Hopf,Hans-Jochen Heinze&Steve Luck在使用ERP和MEG同步记录的技术时发现:在搜索阵列呈现140 ms后,在腹侧枕颞皮质出现了一个与特征选择相关的ERP,紧随这种特征注意效应,在枕颞皮质更前端的位置产生了N2pc。实验证明:在类似视觉搜索范式这种没有提前获知目标刺激所在位置的情况下,特征选择性注意早于视觉空间注意。特征选择会将随后的注意转移引导到这些被选择的特征所在的位置上,以便在此进行高分辨率的分析,从而识别联合目标刺激[9]。
在特征注意的成像研究中,Maurizio Corbetta &Steven Petersen利用PET技术考察了在选择性地注意某些特征(如颜色、形状、运动)时,纹外皮质中所发生的变化。研究发现:当被试在某个特定的刺激维度时,指向颜色、形状或运动的选择性注意在纹外皮质的神经被不同激活而且基本在各自不相重叠的脑区,并因刺激特征维度不同而受到调制。该实验也间接证明了选择性注意在特征分析完成之前便在通道特异的皮质区域内影响对输入的知觉加工[10]。Ariel Schoenfeld,Hans-Jochen Heinze&Steven Hillyard利用fMRI技术定位出了对指向颜色或运动的选择性注意敏感的脑区,并发现这些区域中与注意相关的神经活动在刺激中的运动或颜色变化呈现1 200 ms后出现。
2.5 客体注意
John Duncan将空间注意与客体注意相对比,发现对同一客体可以同时进行两种知觉判断,而且判断正确率不会下降:但如果这2个知觉判断同时对2个不同的物体进行其正确率就会受到影响。Egly指出:在有效线索与无效线索分别指向不同客体时,与指向同一客体相比,空间线索有效性带来的行为上的反应时代价和收益变大。
Kathleen O’Craven,Paul Downing&Nancy Kanwisher通过实验证明:注意对于客体表征存在直接影响,注意促进对被注意客体的所有特征的加工。Notger Mueller&Andreas Kleinschmidt利用fMRI技术证明了空间注意中存在的客体效应,即Egly提出的反应时效应。同时发现,在视觉皮质的V1区到V4区,目标与线索刺激同属同一客体与分属不同客体时相比,未被提示的位置表现出了更强的神经活动,且客体表征可以调制空间注意。
2.6 注意控制网络
Joseph Hopfinger&Maurizio Corbetta利用fMRI技术对注意控制机制进行了研究。研究发现:当线索出现时,如果呈现相同的线索,但被试并不需要注意它们并对其做出反应,但要持续对它们进行被动注视,则相关的自上而下的注意控制网络脑区域(包括上部额叶、下部顶叶、上部颞叶、扣带后皮质的一部分和岛叶)并不会出现激活,虽然视皮质仍然会参与加工线索的视觉特征。除了有意空间注意的额顶叶注意网络,Maurizio Corbetta发现了其他参与注意加工的神经网络——颞顶联合区TPJ和腹外侧前额叶区域。2个系统共同起作用,以使注意可以被引导到相关的位置和潜在的目标,同时也可以在一个新的目标在他处出现时中断这一注意状态,以便注意中心得以转移。
2.7 注意的神经机制总结
综上所述,脑成像研究为注意的早期选择说提供了确切证据,虽然不排除晚期选择的存在,但是后者尚缺乏脑机制方面的证据,有待于未来探讨。尽管大量的认知神经科学研究表明注意对视皮层加工的调节始于外纹状皮层,但是越来越多的证据表明初级视皮层的加工也能受到注意的调节。
1)如果传入的感觉信号含有将被注意的相关的物理特征,这些信号在感觉皮质中就会发生变化。注意控制系统的投射影响着编码这些刺激特征的皮质神经元的兴奋性。
2)实验数据指出了注意效应在知觉加工过程中产生作用的位置,在多个视皮质区区域,被注意的刺激与被忽视的刺激相比能激发更大的神经反应。
3)高度集中的空间注意可以调节视觉系统中皮质下水平的丘脑中继核团的活动,这为早期选择模型提供了证据支持。
4)基于特征的选择性注意对视觉皮质水平加工存在影响,且注意在刺激呈现后经过很短的潜伏期便已对视觉皮质水平加工的较早阶段产生作用。这与早期选择模型的观点一致。
5)与选择性空间注意效应相反,特征注意的潜伏期较长,并且这种注意效应发生在视觉层级性加工系统的较晚阶段。
6)基于空间的注意系统之外,还存在一个基于客体的注意系统。客体的出现会影响空间注意在空间中的分配方式。在一个客体内部,注意扩散会受到促进。当没有空间注意参与时,自上而下的注意控制可以在客体表征这一水平上影响知觉分析。
7)从执行注意控制系统发出的神经投射与感觉特异性皮质区域中的神经元相联系,并经由这种联系改变后者的可兴奋性。如果某个刺激被赋予较高的优先性,它在感觉区引发的神经反应可能会增大;如果它与当前目标无关,对它的反应可能会减小。
3 眼动注意模型
3.1 基于不同控制网络(注意源)的眼动注意模型
以眼动所受的不同注意控制网络(注意源)研究视角来看,眼动的注意模型可分为刺激驱动的眼动注意模型和概念驱动的眼动注意模型。
3.1.1 刺激驱动的眼动注意模型
What模型是一个刺激驱动的眼动注意模型,强调视觉对识别物体的注意,揭示物体内在含义与维持固视的作用[11]。在外显眼动行为上,这种视觉注意表现为固视与其维持时间,其中,固视的维持时间体现了个体对该物体注意的程度。
Where模型也是一个刺激驱动的眼动注意模型,强调视觉注意对确定物体的空间位置和触发眼跳的作用。如果呈现在旁中央凹或者外周视野中的物体,尤其是新颖物体,那么个体视觉的注意可以有意识或无意识地确定其空间方位,并由此触发指向个体感兴趣的物体的眼跳,其中眼跳体现了个体对兴趣物体注意的意图[11]。
3.1.2 概念驱动的眼动注意模型
How模型是一个概念驱动的眼动注意模型,强调主动的视觉注意以及由此而触发的固视和眼跳[11]。在外显眼动行为上,这种视觉注意表现为对于同一刺激、不同的固视对象和不同的眼跳方向。
Spotlight模型也是一个概念驱动的眼动注意的模型,认为视觉注意类似于一个“聚光灯”,强调视觉注意与眼动是分离的,且视觉注意脱离中央凹注视的方向。视觉注意在场景中的转移机制具有持续性,且视觉注意的每次“聚焦”的空间范围是有限的,其中心任务就是定位[11]。当位置确定后,就会触发指向该位置的眼跳。
Morrison模型保留了“聚光灯”理论中将内部注意机制比喻成一个聚光灯的思想,并作了一定修改。该模型认为眼动是由于成功地加工引起的,而不是由于遇到“困难”引起的。当注意机制移动到下一个词的时候,就会有一个信号传到眼动控制系统,进而发生眼跳,使眼睛注视下一个区域[11]。
3.2 基于不同知觉阶段的眼动注意模型
以眼动注意发生在知觉不同阶段的研究视角,眼动的注意模型可分为早选择的眼动注意模型和晚选择的眼动注意模型。
3.2.1 早选择的眼动注意模型
信息加工系统模型包含了感觉输入在内的脑对数据的这个加工过程。这个过程被认为是一个容量有限的阶段,强调只有一定的信息能够通过这一阶段并被眼动注意[1]。在该模型中,大量有可能进入更高层次加工的感觉输入不得不经过筛选,以保证仅允许最重要的事件通过,获得个体的眼动注意。被选择进入更高层次加工或作为无关信息拒绝前,刺激不需要经过完全的知觉加工。Anne Treisman提出,可能非注意通道的信息并不是彻底被阻挡在更高水平的加工之外,而是仅仅退化或被削弱了。
战略战术模型认为,被试在阅读中通常会采用两种策略,即整体性策略和局部策略进行阅读。采用整体性策略,被试尽量使眼睛落在中央凹视野的最佳视觉位置,然后进行眼动注意;同时被试会采用局部策略,当眼睛没有落在最佳注视位置上,被试就会使眼睛移动到最佳注视位置的其他部分,从而对注视点进行再注意。该模型认为影响眼动注意的最主要因素是在注意早期获得的非注视物水平的信息,即眼睛在一个注视点上的最初停留位置,而当注视点落在一个不合适的位置时,会有一次再注视[11]。
Glenmore模型认为,眼跳目标的选择是通过平行加工以及在二维重要性地图内的竞争抑制来完成的[11]。注视中心控制实际的眼跳触发,并调节来自不同认知加工路线的感觉输入。当眼跳被触发时,目标客体将会在重要性竞争中处于优势地位,以赢者的身份出现,从而使眼跳到达目标位置。其中,重要性地图为当前知觉广度内的某一位置赋予一个重要性向量值。
3.2.2 晚选择的眼动注意模型
知觉系统加工模型,认为被注意和被忽视的感觉输入都同等地被知觉系统加工,直到获得语义水平的编码和分析。会存在选择过程来决定信息是否接受更进一步的加工,或是否在觉知中获得表征[1]。该模型指出:眼动注意过程并不能通过改变刺激被感知觉系统加工的方式来影响知觉并获得眼动注意。相反,选择发生在信息加工过程的更高阶段,这些阶段涉及对特定刺激是否应该获得进入觉知的机会,是否应该被编码入记忆,是否在信息加工后获得眼动注意,是否应该触发一个反应的决策[11]。
SWIFT-II模型认为眼睛的运动和信息的加工状态是联系在一起的,而且眼睛向哪里运动和什么时候运动是随着时间的变化而变化的,并主要依赖于之前的注视位置,强调信息的识别按空间分布的形式加工,并且信息加工并不是一个严格的序列过程[11]。根据中央凹到副中央凹视敏度的衰减,该模型认为信息的加工率在中央凹处最高而在外围衰减。眼跳计划包含可变阶段的加工阶段和不可变阶段的加工阶段。眼跳目标即使在眼跳计划启动后仍可改变,眼跳发起时间是一个随机过程,且被中央凹激活量调节。
E-Z读者模型对认知加工与阅读中眼动的关系进行了更为精细的定性说明和定量描述,该模型是序列加工模型。E-Z模型的注意指认知上的而非空间上的注意,强调注意的有序转移[11]。该模型提出眼动计划可分为2步完成:眼跳计划的不稳定阶段M1和眼跳计划的稳定阶段M2。
3.3 眼动模型的评价
眼动的注意模型是在大量实验数据和已观测到的眼动现象基础上提出的,其目的是解释和预测眼动现象。不同的眼动注意模型不断进行改进,并验证模型的相关假设及精确性。各种眼动注意模型对于开展新的实验研究有一定的价值,各种模型发展至今已更具兼容性和完善性,不再像以前对不同的假设具有很强的排他性。不同的模型具有不同的优势,但同时也存在自身的局限:
1)what模型和where模型是刺激驱动的眼动注意模型,强调眼动这一外显的行为可以通过视觉注意间接研究人类大脑中枢是如何加工视觉信息的。视觉注意where模型的加工是平行的,而视觉注意what模型的加工是系列的,两者共同解释了视觉注意的 what与 where方面的完整加工模式。
2)视觉注意与眼动分离,这是眼动研究必须重视的问题。因为如果视觉注意的方向和内容与眼跳方向和眼动的注视内容不能对应起来,那么就不能通过眼动的结果推测个体的视觉注意情况,也不能用眼动的结果推测个体的心理活动。
3)how模型突出个体之间对视觉刺激加工的眼动模式的差异性,以及期望、目的等心理活动对视觉注意和眼动的影响。
4)Morrison模型是一个比较系统的模型,能够解释眼动的基本事实。但此模型不能解释为什么会出现回视,也不能解释注视停留位置是如何选择的。
5)战略战术模型存在的问题是所提出的最佳注视位置的结论是在独立呈现的条件下获得的,而正常的视觉识别要复杂得多,所提出的局部策略的预测并没有得到实验的证实。
6)Glenmore模型的独特之处在于运用重要性地图,为自下而上的视觉加工提供了竞争舞台,将决定何时发生眼跳与视觉识别过程相互分离。虽然指出了语言加工对眼动决定的实质性影响,但没有在信息水平上讨论视觉和语言加工以及眼动控制。
7)SWIFT-II模型采用眼跳计划和眼跳目标选择分离的方法,推迟了目标选择的时间,在很大程度上克服了E-Z读者模型所存在的问题。该模型根据眼跳目标选择激活的动态变化的原则,将初级眼球运动和认知因素结合,认为眼跳计划和眼跳目标选择分离,视觉注意梯度分配,存在一定的自主性,注意分布观点更加清楚。
8)E-Z读者模型是在眼动追踪的实验数据基础上建立的,不但对认知加工与阅读中眼动关系进行了定性的说明,同时进行了定量的描述。该模型存在的缺点是不能深入解释所有阅读中引导眼动的加工,而只是描述了这些加工与眼动结果之间的关系,忽视了更高级水平的语言加工在眼动中的作用。
4 外显眼动-内隐注意的眼动注意模型构建与分析
4.1 模型的构建
Marois指出觉知中在自下而上的刺激影响与自上而下的注意制约间存在着相互作用。本文将What模型、Where模型、How模型和三分模型相结合,设计出了外显眼动-内隐注意的模型,用以描述刺激显著性与自上而下的主动注意控制如何通过互相作用产生不同程度的大脑活动,从而产生不同的眼动和觉知状态。外显眼动-内隐注意的眼动模型见图1。
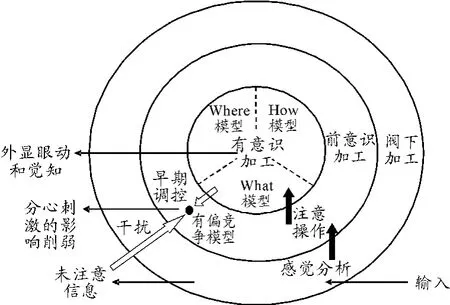
图1 外显眼动-内隐注意的眼动模型
4.2 模型分析
1)阀下加工,即由觉知阀限以下的刺激所产生的大脑活动。当一个加工过程是阀下加工时,其信息无法到达觉知。
2)前意识加工,在这一状态下,大脑中刺激驱动的活动强到足以产生明显的感觉加工,但是在没有自上而下的注意对其信号进行放大的情况下,它不能达到觉知阀限。
3)有意识加工,指的是当刺激足够显著,并且这些信号被自上而下的注意过程所扩大,它们可以越过觉知的阀限[1-2]。
4)What模型强调视觉对识别物体的注意,揭示物体内在含义与维持固视的作用。What模型的视觉神经通路是“P型神经节细胞→外侧膝状体的小细胞胞层→V1区的4Cβ、斑点、斑点区间→V2区的亮带、窄带→V3区→V4区→IT区等”。人眼视网膜接受视觉信息后,经视神经传递至外侧膝状体,在该处换元后传入沈叶的初级视皮层,信息得到初步加工后入颞叶的What神经通路负责加工颜色、形状、空间结构,同时识别静态物体。如果个体对处于中央凹视野中的物体感兴趣,并由此对其进行深度加工,那么个体视觉的注意中心将会始终维持在该物体上。在外显眼动行为上,这种视觉注意表现为固视与其维持时间,其中,固视的维持时间体现了个体对该物体注意的程度[12]。
5)Where模型强调视觉注意对确定物体的空间位置和触发眼跳的作用。Where模型的视觉神经通路是“M型神经节细胞→外侧膝状体的大细胞胞层→V1区的4Cβ、4B层→V2区的宽带→V3区→V5区、MST区→7区等”。人眼视网膜接受视觉信息后,经视神经传递至外侧膝状体,在该处换元后传入沈叶的初级视皮层,信息得到初步加工后入顶叶的Where神经通路负责加工视觉的空间信息,识别运动中的物体。如果呈现在旁中央凹或者外周视野中的物体,尤其是新颖物体,那么个体视觉的注意可以有意识或无意识地确定其空间方位,并由此触发指向个体感兴趣的物体的眼跳,其中眼跳体现了个体对兴趣物体注意的意图[12]。
6)How模型。Gibson强调主动的视觉注意以及由此而触发的固视和眼跳。在个体对刺激做出眼动行为反应前,脑中枢会先根据事物的要求或个体的期望对刺激做出是否需要眼动反应,如果反应,那么对于不同的反应进行加工的这种中枢信息加工的结果就是个体的视觉注意的主动性与随意性。在外显眼动行为上,这种视觉注意表现为对于同一刺激,不同的固视对象和不同的眼跳方向[12]。
7)未注意的信息可以引导神经反应,尽管其强度变弱了,但可以通过产生干扰的方式影响注意信息的加工[13]。
8)有偏竞争模型认为,当一个视觉场景中的不同刺激可能落入同一个视觉神经元的感受野中,2个刺激竞争对神经元发放控制,此时注意可以有利于被注意的刺激的方式帮助解决这种竞争。Abine Kastner等利用fMRI技术发现[14]:在空间集中注意缺失的情况下,同时呈现的临近刺激会相互干扰,从而削弱每个刺激引发的神经反应。然而,当将注意引入,并将其指向刺激阵列中的一个刺激时,同时呈现竞争刺激便不再干扰反应。这种干扰削弱的现象可以被认为是注意削弱了分心刺激的影响,而如果注意不进行干扰,这些分心刺激可能会对神经活动产生干扰。
9)模型中包含一个快速、并行的前注意感觉分析阶段,其后是一个有限能力的注意阶段,这一阶段的输出导致了觉知和不同的外显行为眼动。注意操作相比无意识加工出现较晚,其影响有意识信息被知觉、编码、报告等的方式。
5 结束语
外显眼动-内隐注意的眼动模型是本文根据选择性注意的神经机制并结合已有眼动模型优缺点而设计的眼动模型,用以描述刺激显著性与自上而下的主动注意控制如何通过互相作用产生不同程度的大脑活动,从而产生不同的眼动和觉知状态。该模型的主要创新点在于重视内隐注意对眼动的影响,指出了内隐注意对视觉的重要作用,揭示了在引起大脑觉知产生前的神经机制对输入信息的加工过程和影响。
[1]Michael S G,Richard B L,George R M.Cognitive Neuroscience:The Biology of the Mind[M].W W Norton:3rd International student edition,2014:426-479.
[2]Luck J S,Hollngworth A.Visual memory[M].Oxford: Oxford University Press,2013:21-40.
[3]孙久荣.神经解剖生理学[M].北京:北京大学出版社,2004.
[4]Desaguliers J.A Plain and easy experiment to confirm sir Isaac newton’s doctrine of the different refrangibility of the rays of light[J].Philosophical Transactions,2014,29 (338-350):430-460.
[5]Moran J,Desimone R.Selective attention gates visual processing in the extrastriate cortex[J].Science,1985,229(4715):772-787.
[6]Desimone R.Stimulus-selective properties of inferior tem-poral neurons in the macaque[J].The Journal of Neuroscience,1984,4(8):2041-2069.
[7]Mitchell J P.Activity in right temporo-parietal junction is not selective for theory-of-mind[J].Cerebral Cortex,2011,18:262-289.
[8]Horwitz G D,Albright T D.Short-latency fixational saccades induced by luminance increments[J].Journal of Neurophysiology,2008,90(2):1330-1370.
[9]Nyberg L M A,Cabeza R.General and specific brain regions involved in encoding and retrieval of events:What,where,and when[J].Proceedings of the National Academy of Science,1995,93:11280-11289.
[10]Corbetta M M,Dobmeyer S F,Shulman G.Selective and divided attention during visual discriminations of shape,color and speed:Functional anatomy by position emission tomography[J].Journal of Neuroscience,1991(11): 2383-2410.
[11]闫国利,白学军.眼动研究研究心理学导论[M].北京:科学出版社,2012:94-120.
[12]高闯.眼动实验原理——眼动的神经机制、研究方法与技术[M].武汉:华中师范大学出版社,2012:120 -124.
[13]Sams M H,Rif J R,Knuutila J.The human auditory sensory memory trace persists about 10 sec:Neuromagnetic evidence[J].Journal of Cognitive Neuroscience,1993 (5):363-370.
[14]孙林岩,崔凯,孙林辉.人因工程[M].北京:科学出版社,2011.
(责任编辑杨黎丽)
Research and Innovation in Neural Mechanisms of Selective Attention and Eye Movement Patterns of Attention
XU Peng,ZHANG Jia-jian,ZHAO Bing
(Business School,Hohai University,Nanjing 211100,China)
Based on different standards,we classified eye movement models,and analyzed different models of research emphasis and studied the advantages and problems of each model.This paper summarized the neural mechanisms related to attention and awareness of the role of attention in the research that were tested in humans.On the basis of existing models,we designed a conspicuous move outside-pay attention to the implicit model to describe stimulate significant attention and top-down initiative to control how different levels of brain activity generated by the interaction,thus result in eye movement and awareness of different states.
selective attention;note that the neural mechanisms;note that eye movement patterns
TP18
A
1674-8425(2015)11-0112-08
10.3969/j.issn.1674-8425(z).2015.11.019
2015-04-21
江苏省社科联研究基金资助项目(201035);中央高校基本科研业务费资助项目(2010B10714)
徐鹏(1991—),男,硕士研究生,主要从事技术经济及管理研究。
徐鹏,张家健,赵冰.选择性注意的神经机制和眼动注意模型的研究综述[J].重庆理工大学学报:自然科学版,2015(11):112-119.
format:XU Peng,ZHANG Jia-jian,ZHAO Bing.Research and Innovation in Neural Mechanisms of Selective Attention and Eye Movement Patterns of Attention[J].Journal of Chongqing University of Technology:Natural Science,2015(11):112-119.
猜你喜欢
汽车实用技术(2022年7期)2022-04-20
载人航天(2021年5期)2021-11-20
世界科学技术-中医药现代化(2021年10期)2021-03-02
小资CHIC!ELEGANCE(2021年46期)2021-01-11
睿士(2020年11期)2020-11-16
研究生法学(2020年6期)2020-04-13
中国生物医学工程学报(2019年5期)2019-07-16
中财法律评论(2018年0期)2018-12-06
天津体育学院学报(2016年3期)2016-12-18
专利代理(2016年1期)2016-05-17
