
新疆荒漠绿洲生态区稻水象甲主要生物学特性及发生规律研究
2015-02-14 06:15吐尔逊郭文超付文君关志坚魏振兴
植物保护 2015年1期
王 刚, 吐尔逊, 何 江, 郭文超*, 付文君, 关志坚, 魏振兴
(1.新疆农业科学院植物保护研究所/农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091;2.石河子大学农学院,石河子 832003;3.伊犁州农业技术推广总站,伊宁 835000;4.察布查尔县农业技术推广站,察布查尔 835300)
调查研究
新疆荒漠绿洲生态区稻水象甲主要生物学特性及发生规律研究
王 刚1,2, 吐尔逊1, 何 江1, 郭文超1*, 付文君3, 关志坚4, 魏振兴4
(1.新疆农业科学院植物保护研究所/农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091;2.石河子大学农学院,石河子 832003;3.伊犁州农业技术推广总站,伊宁 835000;4.察布查尔县农业技术推广站,察布查尔 835300)
为明确稻水象甲主要生物学特性及发生规律,探索稻水象甲暴发的原因,为新疆荒漠绿洲生态区稻水象甲的防控提供依据,调查其越冬场所、寄主植物、不同生境中的种群消长动态、年生活史、发生与环境的关系等。稻水象甲越冬成虫于4月上旬末开始活动,4月中旬在杂草、小麦等旱地作物上取食,5月中、下旬随水稻移栽迁入稻田,之后进入产卵期;一代成虫羽化始期为7月中旬,自8月上旬起,一代成虫又开始向其越冬场所附近的杂草丛转移,逐渐入土越冬。稻水象甲在新疆荒漠绿洲生态区一年发生一代,以滞育成虫在土表和浅土层中越冬,越冬的主要场所为稻田附近的林带和田埂。
新疆; 荒漠绿洲生态区; 稻水象甲; 生物学特性; 发生规律
稻水象甲(LissorhoptrusoryzophilusKuschel)又名稻水象、稻根象等,属鞘翅目(Coleoptera),象虫科(Curculionidae),沼泽象亚科(Erirhininae),稻水象属(Lissorhoptrus)[1],是一种重要的国际水稻检疫性害虫。发源于美国密西西比河流域,19世纪后半叶随着农作物种植结构的变化迅速蔓延,成为主要害虫[2]。1959年,在美国加利福尼亚州的萨克拉门托流域发现孤雌生殖型的稻水象甲,而10年后稻水象甲便已分布于整个美国水稻种植区,成为稻水象甲向世界各地扩散的发源地[3]。1988年我国河北省唐海县首次发现稻水象甲[4],随后该虫在我国水稻种植区域内迅速蔓延,严重危害水稻生产,是重要的外来入侵物种[4-5]。该虫主要通过成虫啃食稻叶,幼虫蛀食稻根对水稻等禾本科作物造成危害,一般造成水稻减产15%~20%,严重的减产50%以上,甚至绝收[5]。新疆于2010年6月首次发现该虫[6],截至目前,稻水象甲在新疆仅分布于伊犁河谷地区的察布查尔锡伯自治县、乌鲁木齐市米东区、乌鲁木齐县、昌吉市、五家渠市5个县市,以及上述区域兵团所属团场水稻种植区,约135 km2的区域。
外来物种入侵已经成为影响全球农业可持续发展的重要问题[7],国际上已开展了大量相关研究,在原产地美国,稻水象甲在南部的阿肯色、密苏里和密西西比的部分地区有2个完全的世代;在加利福尼亚则有1个完全世代和1个不完全的世代;在路易斯安那和得克萨斯有时有2个完全世代和1个不完全世代,有时则发生3个完全世代[3]。在日本,按有效积温推测稻水象甲的发育世代数为北海道南部、东北、上信越山区每年1代;关东以西可能发生2代;冲绳地区可能发生3代[8]。朝鲜半岛1年发生1代,有时可发生1个不完全2代[9]。在我国河北、辽宁、吉林、北京和山东每年1代;在浙江温岭、玉环每年1代和1个不完全2代;温州等沿海地区和台湾的双季稻稻区每年2代[10]。但在新疆还未见相关报道,因此,本文在田间调查的基础上,研究了新疆荒漠绿洲生态区稻水象甲主要生物学特性及田间自然消长动态,为该区域有效控制稻水象甲的发生,延缓和阻止其传播,以及科学防治提供依据。
1 材料与方法
1.1 越冬生境调查
在新疆伊犁州察布查尔自治县选择稻水象甲发生密度大的区域,分别调查荒坡、沟渠、田埂、林带等场所0~6 cm深度土样中的虫数,随机取样,每类型取5点,每点取土面积0.25 m2,将土过筛去掉草根、树叶,在沙网中过水,去掉泥土,虫与渣用水漂拣或晒干、拨拣。
1.2 寄主调查
在稻水象甲发生期内,定期在田间和野外进行稻水象甲取食各种植物的情况调查。对有可能是寄主的植物在室内进行生物测定,以成虫为对象进行饲养,每种植物饲喂15~45 d,环境温度保持相对恒定(22~27℃),观察成虫的取食、存活和繁殖情况。根据其取食后生长发育状况,将寄主划分为:1)完全寄主:稻水象甲取食并能正常生长发育完成其世代的植物;2)不完全寄主:取食后不能完成其世代而只能完成其世代中的某个发育阶段的植物;3)偶食寄主:取食后,只能暂时维持某个虫态存活的植物。
1.3 不同生境稻水象甲种群时空动态研究
自4月1日开始,对稻水象甲越冬场所、小麦地及稻田的成虫在杂草、小麦及稻株上的虫量进行定点、定田调查观察,每类型取5点,每点调查0.25 m2,折算出每平方米的成虫数量,每隔3 d调查1次。系统观察田在水稻全生育期不施药防治,自5月上旬至9月中旬每隔3 d调查1次。成虫密度调查:采用五点取样法,每点查1 m2,记录每平方米的虫量;卵粒调查:采用五点取样法,每点查0.25 m2,再按五点取样法取其中5丛,取样后回室内用水洗净泥土,先检查根系,然后由外向内,依次剥取可见叶第1至第3叶鞘,用解剖针仔细拨取叶鞘内的卵粒。幼虫和茧蛹调查:采用五点取样法,每点查0.25 m2,再按五点取样法取其中5丛,每丛连根带泥拔出,放入60~80目的纱网袋和塑料桶内,带回室内在40目的网筛或钢筛内用水慢慢冲洗,在水稻根部细心查找无足型的白色幼虫和土黄色的茧蛹,分别折算每平方米幼虫和茧蛹的数量。
1.4 稻水象甲种群发生与环境的关系研究
在成虫发生高峰期,调查不同水稻品种、连作与轮作、不同播期、不同作物布局、不同水层管理以及不同栽植方式等的稻田中稻水象甲成虫的发生量,采用五点取样法,每点查20丛,折算出百丛虫量。
1.5 数据分析
调查数据采用统计软件IBM SPSSStatistics 20进行方差分析(ANOVA),并采用Duncan氏测验法进行多重比较。
2 结果与分析
2.1 越冬场所
调查结果表明,稻水象甲以滞育成虫在土表和浅土层中越冬,越冬的生境(场所)主要包括稻田附近的林带、田埂,其次是沟渠,再次是荒坡;越冬成虫具有一定的群集性,通常某一越冬地点有多个成虫。越冬生境(场所)的特点:一是背风向阴;二是杂草繁茂;三是土质疏松、较湿润;四是有树叶或杂草覆盖。如图1所示,越冬成虫在林带处的虫口密度最大,与田埂处的虫口密度无显著性差异(P>0.05),与沟渠、荒坡处的虫口密度有显著性差异(P<0.05)。稻水象甲越冬场所的虫口密度由大到小依次为:林带>田埂>沟渠>荒坡。

图1 不同生境中稻水象甲越冬成虫种群密度(新疆伊犁河谷,2013)Fig.1 Adult density ofLissorhoptrus oryzophilusin different overwintering habitats(Ili-Valley Xinjiang,2013)
2.2 寄主植物
在新疆伊犁河谷区域荒漠绿洲稻区,经过系统研究已确认的寄主有禾本科24种,莎草科6种,香蒲科1种,木贼科1种,灯芯草科1种,泽泻科1种,鸢尾科1种,鸭跖草科1种,车前科1种,合计9科37种。其中,稻水象甲独立寄主有:水稻(Oryza sativaLinn.)、杂草稻(Oryza sativaf.spontanea)、稗草[Echinochloa crusgalli(L.)Beauv.]、香附(莎草)(Cyperus rotundusL.)、异型莎草(Cyperus difformisL.)。次要寄主有:小麦(Triticum aestivumLinn.)、玉米(Zea maysL.)、高粱(Sorghum vulgarePers.)、李氏禾(Leersia hexandraSwartz)、马唐[Digitaria sanguinalis(L.)Scop.]、白茅[Imperata cylindrica(Linn.)Beauv.]、狗牙根[Cynodon dactylon(Linn.)Pers]、狗尾草[Setaria viridis(L.)Beauv.]、扁秆藨草(Scirpus planiculmisFr. Schmidt)、碎米莎草(Cyperus iriaL.)、球穗扁莎草[Pycreus globosus(All.)Reichb.]、节节草(Equisetum ramosissimumDesf.)。偶食寄主有:冰草[Agropyron cristatum(Linn.)Gaertn.]、看麦娘(Alopecurus aequalisSobol.)、牛筋草[Eleusine indica(L.)Gaertn.]、黑麦草(Lolium perenneL.)、早熟禾(Poa annuaL.)、荩草[Arthraxon hispidus(Thunb.)Makino]、画眉草[Eragrostis pilosa(L.)Beauv.]、白草(Pennisetum centrasiaticumTzvel.)、千金子[Leptochloa chinensis(L.)Nees]、燕麦草[Arrhenatherum elatius(Linn.)Presl]、耿氏硬草[Sclerochloa kengiana(Ohwi)Tzvel.]、野燕麦(Avena fatuaL.)、芦苇(Phragmites communisTrin.)、鸭跖草(Commelina communisLinn.)、车前草(Plantago asiaticaL.)、鸢尾草(Iris tectorumMaxim.)、欧泽泻(Alisma plantago-aquaticaL.)、灯心草(Juncus effususLinn.)、牛毛毡[Heleocharis yokoscensis(FranchetSav.)TangetWang]、小香蒲(Typha minimaFunk.)。
2.3 不同生境稻水象甲种群时空动态
研究表明,在新疆伊犁河谷水稻种植区稻水象甲越冬成虫于4月上旬末开始出土,先在越冬场所附近的杂草丛活动并取食;到4月中旬末,则逐渐扩散开,在杂草、小麦等旱地作物上活动、取食;5月上旬,自秧田揭膜起,秧田即可见成虫;插秧后,本田即有成虫。5月中、下旬的晴天黄昏,可见大量成虫自旱地向水田迁飞,5月下旬,水稻本田中越冬成虫达到高峰期。越冬成虫于5月下旬末开始产卵,6月上旬,是其产卵高峰期,产卵期一直持续到7月上旬,产卵结束后越冬成虫逐渐死亡。稻水象甲幼虫从6月上旬开始出现,数量逐渐增多,到6月下旬末达到最大,之后幼虫数量逐渐下降。稻水象甲蛹自6月下旬开始出现,数量逐渐增多,到7月中旬达到最大,之后蛹数量逐渐下降。成虫羽化始期为7月中旬,盛期出现在7月下旬至8月上旬。自8月上旬起,稻水象甲又开始向其越冬场所附近的杂草丛转移。成虫于8月中旬开始入土越冬,到9月下旬才见不到活动的成虫。不同环境中稻水象甲种群时空动态如图2所示。

图2 不同生境中稻水象甲种群时空动态(新疆,伊犁河谷,2013)Fig.2 Population dynamics ofLissorhoptrus oryzophilusin different habitats(Ili-Valley Xinjiang,2013)
2.4 稻水象甲生活史
稻水象甲在新疆荒漠绿洲生态区一年发生一代。该虫以滞育成虫在稻田附近的林带、田埂、沟渠、荒坡等有覆盖场所0~6 cm的土壤中越冬。越冬成虫于4月上旬末开始出土。4月中旬为出土高峰期,出土期一直延续至4月下旬。成虫出土后爬行或迁飞寻找食物,在栽培寄主出土前,首先取食马唐、莎草等野生寄主叶片。5月上旬,自秧田揭膜起,秧田即可见成虫;插秧后,本田即有成虫。5月中、下旬的晴天黄昏,可见大量成虫自旱地向水田迁飞。越冬后成虫产卵时期为5月下旬,盛期出现在6月上、中旬。新一代幼虫发生期为6月上旬,盛期出现在6月中旬至7月上旬。新一代幼虫化蛹始期为6月下旬,盛期出现在7月上、中旬。新一代成虫羽化始期为7月中旬,盛期出现在7月下旬至8月上旬。新一代成虫入土越冬始期为8月中旬,盛期出现在8月下旬至9月上旬。越冬休眠期长达6~8个月(稻水象甲年生活史如表1所示)。
2.5 稻水象甲种群发生与环境的关系
本研究只对新疆伊犁河谷下游荒漠绿洲生态区不同环境条件下稻水象甲(成虫)发生高峰期种群数量进行比较。稻水象甲在新疆荒漠绿洲生态区一年发生一代,水稻本田中一年有两个成虫发生高峰期,即5月下旬出现的越冬成虫高峰期和7月下旬出现的一代成虫高峰期。研究结果表明,在其他栽培措施相同的条件下,不同水稻品种之间,稻水象甲成虫发生量无明显差别。在管理水平相同的条件下,连作田与轮作田之间,稻水象甲成虫发生量也无明显差别。不同的播期、作物布局、水层管理及栽植方式环境条件下,稻水象甲成虫高峰期存在显著差异。
稻水象甲成虫取食具有一定的趋嫩性,5月下旬时,播种早且移栽早的水稻已处于分蘖末期,叶片已变硬,稻水象甲则逐渐向移栽晚的稻田中寻找较嫩的水稻叶片。如图3所示,移栽早的稻田中稻水象甲成虫高峰期发生量显著小于移栽晚的稻田中稻水象甲成虫高峰期发生量(P<0.05),尤其是一代稻水象甲成虫高峰期发生量。因此,早播、早移栽可适当减轻稻水象甲发生量。
在新疆伊犁河谷区域稻水象甲越冬成虫是由旱地逐渐向水田转移的,水旱相邻的稻田中越冬成虫基数相对较大,稻水象甲发生量较大。由图4可知,在不同作物布局条件下,管理水平相同时,水旱相邻的稻田中稻水象甲成虫高峰期发生量显著高于水稻田中央田块中稻水象甲成虫高峰期发生量(P<0.05)。

图4 不同作物布局稻田稻水象甲(成虫)发生高峰期种群数量对比(新疆伊犁河谷,2013)Fig.4 The peak number ofL.oryzophilusadults in different crops layout paddy(Ili-Valley Xinjiang,2013)
在新疆伊犁河谷区域稻水象甲的发生与水分条件密切相关。产卵和幼虫孵化离不开水,在水稻分蘖末期进行浅水管理或适时排水晒田,使成虫产卵和孵化处于不利环境中(因成虫一般在水下水稻叶鞘中产卵),并增加卵和幼虫的死亡率(因幼虫孵化后在地表活动一段时间再钻到水稻根部取食)。而浅水管理或干湿交替管理可有效抑制水稻无效分蘖,对水稻生长更为有利。5月末、6月初正是荒漠绿洲区水稻分蘖末期,适时晒田对稻水象甲一代成虫高峰期发生量有明显的抑制作用。如图5所示,长期深水灌溉的稻田中稻水象甲越冬成虫高峰期发生量与适时晒田的稻田中稻水象甲越冬成虫高峰期发生量无显著性差异(P>0.05),但两种不同水层管理方式下,适时晒田的稻田中稻水象甲一代成虫高峰期发生量显著小于长期深水灌溉的稻田中稻水象甲一代成虫高峰期发生量(P<0.05)。因此,在实际生产中,适时晒田作为农艺措施可有效抑制控制稻水象甲种群的发展。
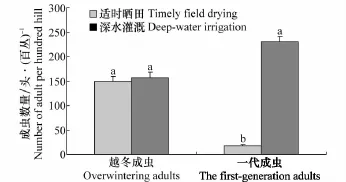
图5 不同水层管理稻田稻水象甲(成虫)发生高峰期种群数量对比(新疆伊犁河谷,2013)Fig.5 The peak number ofL.oryzophilusadults in different water management paddy(Ili-Valley Xinjiang,2013)
实际生产中,直播稻田长期处于干湿交替的状态,不利于稻水象甲的发生。由图6可知,在不同栽植方式条件下,管理水平相同时,插播的稻田中稻水象甲成虫高峰期发生量显著大于直播的稻田中稻水象甲成虫高峰期发生量(P<0.05)。因此,采用直播这种栽植方式可适当减轻稻水象甲的发生。

图6 不同栽植方式稻田稻水象甲(成虫)发生高峰期种群数量对比(新疆伊犁河谷,2013)Fig.6 The peak number ofL.oryzophilusadults in different ways of planting paddy(Ili-Valley Xinjiang,2013)
3 结论与讨论
本文通过调查发现,稻水象甲以滞育成虫在土表和浅土层中越冬,越冬的主要场所为稻田附近的林带和田埂,与武清彪[11]的研究结果基本一致。柴一秋[12]发现稻水象甲越冬场所背阴向阳,而本研究中却发现稻水象甲越冬场所背风向阴,关于两者存在明显差异的原因,有待进一步探究。
在寄主植物方面,日本的Asayama等[13]研究发现,稻水象甲成虫寄主有禾本科61种,莎草科15种,灯心草科4种,鸭跖草科和鸢尾科各2种,泽泻科和香蒲科各1种,合计7科86种,幼虫寄主有禾本科12种,莎草科6种,灯心科和鸭跖草科各2种,泽泻科1种,合计5科23种。孙富余[14]、商晗武[15]、李志强[16]等研究表明,稻水象甲寄主有10科64种,主要以禾本科、莎草科植物为主,水稻、玉米及高粱受害最为严重。而本研究中已确认的稻水象甲寄主有禾本科24种,莎草科6种,香蒲科1种,木贼科1种,灯芯草科1种,泽泻科1种,鸢尾科1种,鸭跖草科1种,车前科1种,合计9科37种;其中,新发现稻水象甲成虫取食木贼科的节节草和车前科的车前草。寄主植物种类差异可能与稻水象甲发生区寄主植物分布和生物多样性等因素存在一定的关系。
在生物学特性研究方面,本研究和前人研究结果基本一致,稻水象甲越冬成虫于4月上旬末出土,4月中旬逐渐扩散开,在杂草、小麦等旱地作物上活动、取食,5月中、下旬迁入水田,之后越冬成虫进入产卵期,幼虫从6月上旬末开始出现,蛹自6月下旬开始出现,一代成虫羽化始期为7月中旬,自8月上旬起,一代成虫又开始向其越冬场所附近的杂草丛转移,逐渐入土越冬。因稻水象甲个体小、幼虫期和蛹期完全在有泥土包裹的水稻根部中完成,人工饲养技术不成熟,还不能准确得出稻水象甲的发育历期和发育速率,有待进一步的观察和研究。
在发生与环境的关系方面,研究表明,不同水稻品种之间,连作与轮作方式之间,稻水象甲发生严重程度无明显差异;不同的播期、作物布局、水层管理、栽植方式之间,稻水象甲发生严重程度有明显差异。因稻水象甲的成虫取食具有一定的趋嫩性,移栽早的稻田中水稻叶片硬化程度高,稻水象甲发生量较少;越冬成虫是由旱地逐渐向水田转移的,水旱相邻的稻田中越冬成虫基数相对较大,稻水象甲发生量较大;稻水象甲的发生与水分条件密切相关,产卵和幼虫孵化离不开水,在水稻分蘖末期适时排水晒田,可使成虫产卵和幼虫孵化处于不利环境中。5月末、6月初正是荒漠绿洲区水稻分蘖末期,适时晒田对稻水象甲一代成虫高峰期发生量有明显的抑制作用;实际生产中,直播稻田长期处于干湿交替的状态,不利于稻水象甲的发生。因此,播种早且移栽早、适时晒田、直播的水稻田块,可有效控制稻水象甲种群发生和危害。
[1] 全国农业技术推广服务中心.中国稻水象甲[M].北京:中国农业出版社,1998.
[2] Grigarick A A.Study on the rice water weevil,past,present,and future in the United States with emphasis on California[C]∥Hirai K.Establishment,spread,and management of the rice water weevil and migratory rice insect pest in East Asia.NARC,Tsukuba,1993:12 31.
[3] Way M O,Wallace R G.Rice water weevil integrated pest management in the United States with emphasis on the South Immigration and distribution patterns of the rice water weevil,Lissorhoptrusoryzophilus,in a paddy field in relation to the total effective temperature[C]∥Hirai K.Establishment,spread,and management of the rice water weevil and migratory rice pests in East Asia.NARC:Tsukuba,1993:58 82.
[4] 孙汝川,毛志农.稻水象甲[M].北京:农业出版社,1996.
[5] 丁玉芝.稻水象甲的发生与防治[J].植物保护,2002,28(2):57.
[6] 郭文超,吐尔逊,魏振兴,等.新疆首次发现水稻重大外来有害生物稻水象甲[J].新疆农业科学,2011,48(1):70 74.
[7] Tabashnik B.Pest adaptation[J].Nature,1997,389:778.
[8] Asayama T,Nakagome T.Invasion and spread of the rice water weevil,LissorhoptrusoryzophilusKuschel,in Japan[C]∥Hirai K.Establishment,spread,and management of the rice water weevil and migratory rice insect pest in East Asia. NARC:Tsukuba,1993:83 103.
[9] Takeda M.Immigration and distribution patterns of the rice water weevil,Lissorhoptrus oryzophilusKusche1,in a paddy field in relation to the total effective temperature[C]∥Hirai K.Establishment,spread,and management of the rice water weevil and migratory rice pests in East Asia.NARC:Tsukuba,1993:58 82.
[10]吕利华,马春森.信安市稻水象甲的传入及蔓延的控制对策[J].吉林农业科学,1996(3):40 42.
[11]武清彪,郑卫锋,武宇鹏,等.稻水象甲越冬场所的调查及防治对策[J].植物检疫,2005,19(5):314 315.
[12]柴一秋,陈祝安,冯惠英,等.金龟子绿僵菌对稻水象甲的致病性[J].中国生物防治,2000,16(1):22 25.
[13]Asayama T,Iori A,Kanoo M,et al.Control of the rice water weevil in Japan[C]∥Iwata S,Kurita T,Kocha T,et al.Japan Plant Protection Association,Tokyo,1986:1 175.
[14]孙富余,赵成德,田春晖,等.稻水象甲的发生规律与防治研究Ⅷ.稻水象甲综合防治技术[J].辽宁农业科学,1997(6):7 10.
[15]商晗武,程家安,蒋明星,等.起始供食时间对稻水象甲冬后成虫繁殖、取食和存活的影响[J].中国水稻科学,2003,17(1):77 81.
[16]李志强,于凤泉,田春晖,等.辽宁省不同地域稻水象甲分子遗传差异及其意义研究[J].辽宁农业科学,2006(1):1 3.
Biological characteristics and occurrence regulation of rice water weevil(Lissorhoptrus oryzophilusKuschel)in desert oasis ecological zone of Xinjiang
Wang Gang1,2, Tuerxun1, He Jiang1, Guo Wenchao1, Fu Wenjun3, Guan Zhijian4, Wei Zhenxing4
(1.Institute of Plant Protection,Xinjiang Academy of Agricultural Sciences/Key Laboratory of Integrated Pest
Management on Crops in Northwestern Oasis,Ministry of Agriculture,P.R.China,Urumqi830091,
China;2.College of Agronomy,Shihezhi University,Shihezi832003,China;3.Station of
Agricultural Techniques Extension in Yili of Xinjiang,Yining835000,China;4.Station
of Agricultural Techniques Extension in Qapqal County of Xinjiang Yili,835300,China)
The major biological characteristics and occurrence regulation of rice water weevil,Lissorhoptrus oryzophilusKuschel,were surveyed,and reasons for the outbreak ofL.oryzophiluswere explored in order to provide evidence for controllingL.oryzophilusin desert oasis ecological zone of Xinjiang.We surveyed its overwintering sites,host plant species,population dynamics in different habitats,life history,relationship with the environment and so on.OverwinteringL.oryzophilusbegan unearthed at the end of the early April,then gradually feed on the weeds,wheat and other dryland crops in mid-April.A large number of adults migrated from dryland to paddy fields in mid to late May and began to oviposit in late May.The first generation adults emerged in mid-July.Since the early August,L.oryzophilusbegan to transfer to the weed fields,for overwintering.L.oryzophilushas one generation a year in desert oasis ecological zone of Xinjiang,and overwinter as diapause adults on the soil surface or in shallow soil.The main overwintering sites were forest and ridge nearby the rice fields.
Xinjiang; desert oasis ecological zone;Lissorhoptrus oryzophilus; biological characteristics;occurrence regulation
S 435.112
A
10.3969/j.issn.0529 1542.2015.01.028
2014 01 09
2014 04 01
新疆维吾尔自治区自然科学基金(2013211A041)
*通信作者 E-mail:gwc1966@163.com
猜你喜欢
当代水产(2022年7期)2022-09-20
农家参谋(2021年10期)2021-12-05
今日农业(2021年21期)2021-11-26
当代水产(2021年4期)2021-07-20
农民致富之友(2019年32期)2019-11-23
今日农业(2019年13期)2019-01-03
福建农业科技(2018年2期)2018-09-10
中国农业文摘-农业工程(2016年5期)2016-04-12
学生天地·小学中高年级(2015年11期)2016-01-14
环球时报(2009-09-11)2009-09-11
