
青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征
2015-02-11 02:06:25雒明伟李倩倩贾婷婷郭正刚
生态学报 2015年23期
雒明伟, 毛 亮, 李倩倩, 赵 旭, 肖 玉, 贾婷婷, 郭正刚
兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020
青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征
雒明伟, 毛 亮, 李倩倩, 赵 旭, 肖 玉, 贾婷婷, 郭正刚*
兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 兰州 730020
恢复筑路取土迹地植物群落是青藏高原退化高寒草地恢复的重要组分,而生态化学计量是揭示退化草地自然恢复过程中土壤和植物间养分互作的重要方法。通过调查青藏公路筑路取土迹地自然恢复群落和天然群落内植物和土壤C、N、P的含量及其比值,研究了恢复群落和天然群落的C、N、P生态化学计量特征。结果表明:经历18a自然恢复后,不同地点筑路取土迹地均已逐步实现植物群落的定居,但其恢复程度存在明显差异。恢复群落植物叶片N含量高于天然群落,导致其叶片C∶N较低,表明恢复群落植物的N利用效率较低,N含量较高的模式主要原因可能是豆科植物比例较高和土壤有效N的供应较充足所致。恢复样地在0—10cm和10—20cm的土层内的有机碳(SOC)都显著低于天然样地,恢复样地土壤全氮(STN)仍一定程度上低于天然样地但含量较接近,恢复样地在10—20cm土层中土壤全磷(STP)含量较高,这说明恢复群落尽管在土壤恢复方面并未达到天然群落的水平,但已得到了明显改善,土壤肥力正发生着正向的演替。叶片N含量与土壤STN相关性不显著,叶片P含量与土壤STP含量显著地正相关,表明植物叶片P含量在一定程度上受到土壤环境中全磷的影响。综合N∶P判定阈值和叶片土壤养分相关分析结果表明研究地区草地植被主要受到P元素的限制,且工程迹地草地恢复群落比天然群落容易受到P元素的限制。
工程迹地; 碳、氮、磷; 化学计量; 高寒草地; 恢复群落
修建青藏公路和青藏铁路时,因直接移除路基两侧植被和表层土壤而在高寒草地区而形成了诸多分布广泛的取土迹地[1-2]。其干扰草地景观的整体性,迫使天然草地发生带状退化,这种退化不同于过度放牧、鼠害、冻融等因素引起的退化[3],这是因为筑路取土具有突然性,不仅瞬间移走了富含养分的表层土壤和种子等植物潜在繁殖体[2-4],而且迅速成为水土流失的源地[1]。恢复筑路取土迹地的植物群落既是保证路基安全和控制水土流失的关键,又是青藏高原退化高寒草地恢复的重要组成部分[5]。
取土迹地植被恢复过程实质是植物与土壤相互适应和反馈的过程。土壤为植被恢复提供养分,而恢复的植被通过物种间竞争和替代作用,以及植物与土壤间的适应过程而实现改良土壤肥力[6]。取土迹地恢复可分为人为重建和自然恢复两种策略,其中自然恢复策略具有经济性高和避免草皮移植对客地原生草地破坏等优点,已经被广泛的应用于工矿用地迹地的恢复[7],其本质是通过土壤内残存的植物繁殖体在适宜环境条件下发芽、发育和生殖,或者是取土迹地周围植物的繁殖体通过入侵和定居的方式再次占据取土迹地[8]。已有研究表明,青藏公路取土迹地植被恢复的周期约20a以上[9],且其恢复程度受取土迹地面积大小的影响[2]。目前关于青藏公路取土迹地植被恢复的研究主要集中于已恢复群落的初级生产力、植物多样性特征、草地生态系统结构和生物生境条件等方面[5,10-12],然而取土迹地植被恢复过程中植物和土壤间的相互作用关系尚不清楚。
生态化学计量主要通过分析植物和土壤内的C、N、P含量及其计量关系[13],解析土壤C、N、P间的组成和配置关系调控植物生长和发育的机理,因此C、N、P的组成和分配一定程度上决定着植物群落内的种群密度和种间竞争格局,从而最终决定群落演替趋向[14]。因此研究青藏公路取土迹地植物和土壤C、N、P含量及其计量比关系,不仅能够揭示取土迹地草地群落恢复过程中植物物种更替和土壤养分供给间的相互关系,而且有利于从养分供给角度剖析植物对环境的适应性以及反馈效果,诊断恢复群落正向演替时的限制养分。
本研究通过调查青藏公路取土工程迹地经过18a自然恢复的恢复群落和天然群落内植物和土壤C、N、P元素的计量特征,以期确定草地植被正向恢复的限制元素,为青藏高原取土迹地植被恢复提供科学决策依据。
1 研究区概况
研究区域位于青藏公路沿线西大滩(94°16.830′ E, 35°44.578′ N)至沱沱河(92°29.756′E, 34°17.192′N)段,全长300km,海拔约为4300—5100m。气候属高原亚寒带半干旱气候,分冷暖两季,冷季寒冷漫长,冻结期长达7—8 个月,暖季凉爽短促,植被生长期为3—4 个月[15]。年均降水量在200—400mm,84%的年降雨发生在7月到9月的生长季节,年均气温在-1.3—-4.1 ℃[1,16]。植被类型主要是以紫花针茅(Stipapurpurea)为优势种的高寒草原和以苔草属(Carexspp.)为主的高寒草甸,高寒草原常见主要伴生种是异针茅(Stipaaliena),沙生凤毛菊(Saussureaarenaria),甘青兔耳草(Saxifragatangutica),扇穗茅(Littledaleatibetica),早熟禾(Poaannua)等;高寒草甸常见主要伴生种是高山嵩草(Kobresiapygmaea),矮火绒草(Leontopodiumnanum),垫状点地梅(Androsacetapete),小叶棘豆(Oxytropismicrophylla)等。高寒草原土壤为高山高原土,高寒草甸土壤为高山草甸土,两者土层均较薄[8]。
2 研究方法
2.1 试验设计和取样
野外调查与取样时间为2012年7月中下旬,是植物生长的旺盛期。沿1994年扩建和修缮青藏公路时遗留的取土迹地布设样地,将取土迹地自然恢复的植物群落定为恢复群落,以毗邻取土迹地的天然草地(未受工程干扰)为天然群落(对照)。以海拔、植被类型和地貌特征为参考依据,选择7 个研究调查和取样点,分别为西大滩、不冻泉、五道梁、乌丽、风火山、可可西里和沱沱河(表1)。
每个调查点分别设置4对配对成组的 10m×10m的恢复群落样地和天然群落样地,4组样地平行布置且总体分布方向都是垂直于公路,同组恢复样地与天然样地距离100m以上,每个样地内采用对角线法设置4个1 m×1 m的样方,调查记录样方内的植物种、盖度,收获地上生物量,然后带回实验室烘干称重。样方调查完后,利用内径为3.5 cm的土钻在每个样地内沿对角线打钻5次,按0—10cm和10—20cm两层收集土样,去除植物根及残体、石头等杂质后,将每个样地内同一土层土样合并为一个样品,室温下自然风干,充分研磨,经过0.25 mm土筛过筛后储藏,以备分析。为测定群落水平植物叶片的养分含量,样地中随机再选取了3个1 m×1 m的样方,齐地面刈割草地植物,摘下植物叶片后,将3个样方内植物叶片混合均匀后,用四分法获取成熟、健康植物叶片样品。叶片带回实验室,105 ℃杀青15 min,65 ℃烘干至恒重,粉碎后密封保存,以备分析。
2.2 样品分析
土壤和植物叶片C、N、P含量均采用常规分析方法测定,其中植物和土壤有机碳采用外加热、重铬酸钾容量法测定,全氮采用凯氏定氮法测定,全磷采用钼锑抗比色法测定[17]。
2.3 数据处理
应用Excel2010和SPSS18.0统计分析软件对数据进行整理、分析和制图。利用独立样本t检验比较恢复群落与天然群落植物养分特征的差异;由于土壤具有高度的空间异质性,将每个研究位点恢复样地与天然样地配比成对,运用配对样本t检验比较两者土壤养分的差异;运用Pearson相关分析植物与土壤C、N、P含量及计量特征的相关性,其中植物叶片C、N、P含量为恢复群落和天然群落的数据,土壤C、N、P含量为0—10cm和10—20cm土层的平均值。
3 结果与分析
3.1 恢复群落与天然群落的群落特征
筑路取土迹地上植被群落筑路过程中遭受严重破坏,生物部分几乎为零,但经历18a的自然恢复后,虽然筑路迹地上均生长和分布着植物群落,但较天然草地而言,不同地点植物恢复程度存在明显差异(表1)。不冻泉恢复群落盖度和天然群落盖度基本接近,而其余地点恢复群落盖度表现为不同程度地低于天然草地。不冻泉恢复群落生物量与天然群落生物量接近,五道梁恢复群落生物量大于天然群落生物量,其余地点恢复群落生物量均低于天然草地,这是因为不冻泉取土迹地较好的土壤水热条件为以小叶棘豆(Oxytropismicrophylla)(生物量占50%以上)为主的先锋物种定居和茁壮生长创造了良好的条件,而五道梁恢复群落生物量较大的主要原因是恢复群落内阔叶独一味占据优势,其较高的生物量提升了恢复群落的生物量。植物群落组成方面,沱沱河、乌丽、西大滩的恢复群落均已出现与天然草地一致的优势种,而不冻泉、可可西里和乌丽的恢复群落与天然群落仅存在1—2种相同的主要伴生种。说明青藏高原草地区筑路取土迹地恢复群落演变方向有些地点趋向于毗邻的天然草地,而有些地点的恢复群落仍然处于分化的未稳定状态。
表1 青藏高原高寒草地区筑路取土迹地恢复群落和天然群落的特征
Table 1 Features of restorable plant communities at land used for highway construction and natural plant communities in the alpine grassland regions of Qinghai-Tibet Plateau
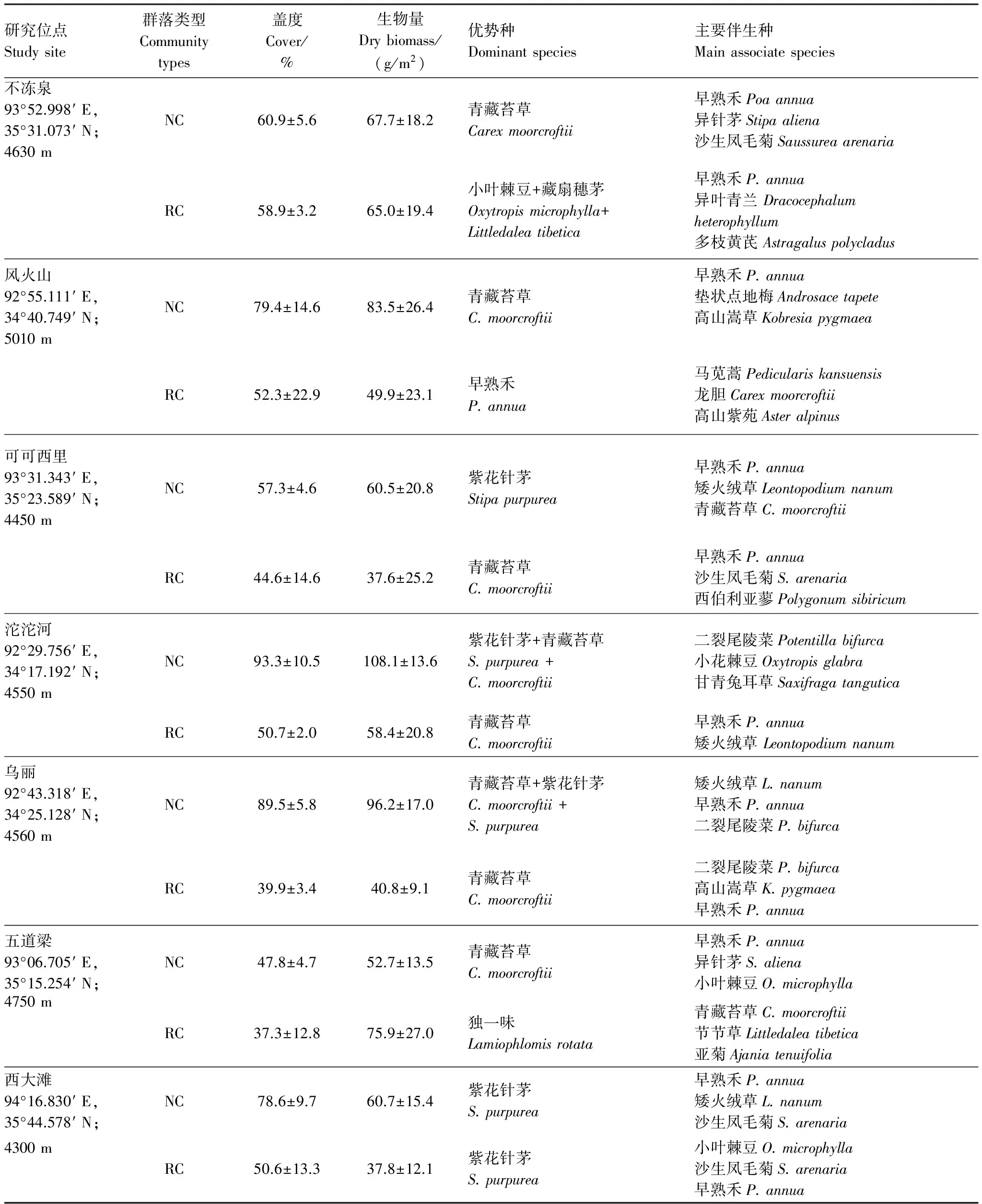
研究位点Studysite群落类型Communitytypes盖度Cover/%生物量Drybiomass/(g/m2)优势种Dominantspecies主要伴生种Mainassociatespecies不冻泉93°52.998′E,35°31.073′N;4630mNC60.9±5.667.7±18.2青藏苔草Carexmoorcroftii早熟禾Poaannua异针茅Stipaaliena沙生凤毛菊SaussureaarenariaRC58.9±3.265.0±19.4小叶棘豆+藏扇穗茅Oxytropismicrophylla+Littledaleatibetica早熟禾P.annua异叶青兰Dracocephalumheterophyllum多枝黄芪Astragaluspolycladus风火山92°55.111′E,34°40.749′N;5010mNC79.4±14.683.5±26.4青藏苔草C.moorcroftii早熟禾P.annua垫状点地梅Androsacetapete高山嵩草KobresiapygmaeaRC52.3±22.949.9±23.1早熟禾P.annua马苋蒿Pediculariskansuensis龙胆Carexmoorcroftii高山紫苑Asteralpinus可可西里93°31.343′E,35°23.589′N;4450mNC57.3±4.660.5±20.8紫花针茅Stipapurpurea早熟禾P.annua矮火绒草Leontopodiumnanum青藏苔草C.moorcroftiiRC44.6±14.637.6±25.2青藏苔草C.moorcroftii早熟禾P.annua沙生凤毛菊S.arenaria西伯利亚蓼Polygonumsibiricum沱沱河92°29.756′E,34°17.192′N;4550mNC93.3±10.5108.1±13.6紫花针茅+青藏苔草S.purpurea+C.moorcroftii二裂尾陵菜Potentillabifurca小花棘豆Oxytropisglabra甘青兔耳草SaxifragatanguticaRC50.7±2.058.4±20.8青藏苔草C.moorcroftii早熟禾P.annua矮火绒草Leontopodiumnanum乌丽92°43.318′E,34°25.128′N;4560mNC89.5±5.896.2±17.0青藏苔草+紫花针茅C.moorcroftii+S.purpurea矮火绒草L.nanum早熟禾P.annua二裂尾陵菜P.bifurcaRC39.9±3.440.8±9.1青藏苔草C.moorcroftii二裂尾陵菜P.bifurca高山嵩草K.pygmaea早熟禾P.annua五道梁93°06.705′E,35°15.254′N;NC47.8±4.752.7±13.5青藏苔草C.moorcroftii早熟禾P.annua异针茅S.aliena小叶棘豆O.microphylla4750mRC37.3±12.875.9±27.0独一味Lamiophlomisrotata青藏苔草C.moorcroftii节节草Littledaleatibetica亚菊Ajaniatenuifolia西大滩94°16.830′E,35°44.578′N;NC78.6±9.760.7±15.4紫花针茅S.purpurea早熟禾P.annua矮火绒草L.nanum沙生凤毛菊S.arenaria4300mRC50.6±13.337.8±12.1紫花针茅S.purpurea小叶棘豆O.microphylla沙生凤毛菊S.arenaria早熟禾P.annua
NC:Natural community,天然群落;RC:Restorable community,恢复群落
3.2 恢复群落与天然群落的植物叶片养分及计量特征
高寒草地群落植物总体水平上叶片C、N、P含量分别为(408.64±61.21)mg/g、(18.60±1.81)mg/g、(1.42±0.50)mg/g;C∶N为22.10±3.55,C∶P为320.70±118.04,N∶P为14.53±4.84(表2)。恢复群落叶片N含量显著地高于天然群落(P<0.05),而叶片C、P含量在恢复群落和天然群落间差异不显著。恢复群落叶片C∶N显著地低于天然群落(P<0.05),而C∶P和N∶P并没有表现出显著差异,说明筑路时所产生的生境变化明显影响了恢复群落植物叶片的N含量。
表2 青藏高原高寒草地区筑路取土迹地恢复群落与天然群落植物叶片养分含量及化学计量比(平均值±标准偏差)
Table 2 The leaf C、N、P concentration and stoichiometric ratio(Mean±SD)of restorable plant communities at land used for highway construction and natural plant communities at community level in the alpine grassland regions of Qinghai-Tibet Plateau
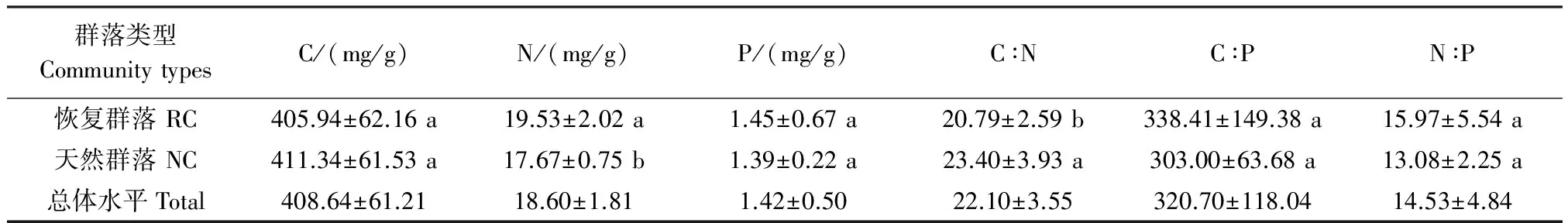
群落类型CommunitytypesC/(mg/g)N/(mg/g)P/(mg/g)C∶NC∶PN∶P恢复群落RC405.94±62.16a19.53±2.02a1.45±0.67a20.79±2.59b338.41±149.38a15.97±5.54a天然群落NC411.34±61.53a17.67±0.75b1.39±0.22a23.40±3.93a303.00±63.68a13.08±2.25a总体水平Total408.64±61.2118.60±1.811.42±0.5022.10±3.55320.70±118.0414.53±4.84
独立样本t检验,不同字母表示P<0.05水平上差异显著
3.3 恢复样地与天然样地土壤养分含量和计量特征
配对t检验结果表明,恢复群落0—10cm土层内有机碳(SOC)和全氮含量(STN)显著低于天然群落相应土层内SOC和STN含量(P<0.05),而土壤全磷(STP),SOC∶STN,SOC∶STP,STN∶STP在恢复群落和天然群落间均未表现出显著差异(表3);恢复群落10—20cm土层内SOC含量显著低于天然群落相应土层内的SOC含量(P<0.05),而STP含量显著高于天然群落相应土层内的STP含量,恢复群落10—20cm土层内的SOC∶STP显著地低于天然样地对应土层的SOC∶STP,恢复群落STN含量也低于天然草地但未表现出显著的差异。
无论恢复群落还是天然群落,土壤SOC含量均表现为0—10cm土层显著大于10—20cm土层(表4)(P<0.05)。STN含量在天然草地内表现为0—10cm显著大于10—20cm(P<0.05),但恢复群落内0—10cm和10—20cm间STN含量差异不显著。天然群落和恢复群落内STP含量,SOC∶STN,SOC∶STP和STN∶STP在土层间没有明显差异。
表3 青藏高原高寒草地区筑路取土迹地恢复群落与天然群落相同土层土壤养分含量及计量比(平均值±标准偏差)
Table 3 The soil C、N、P concentration and stoichiometric ratio of restorable plant communities at land used for highway construction and natural plant communities in the alpine grassland regions of Qinghai-Tibet Plateau(Mean±SD)
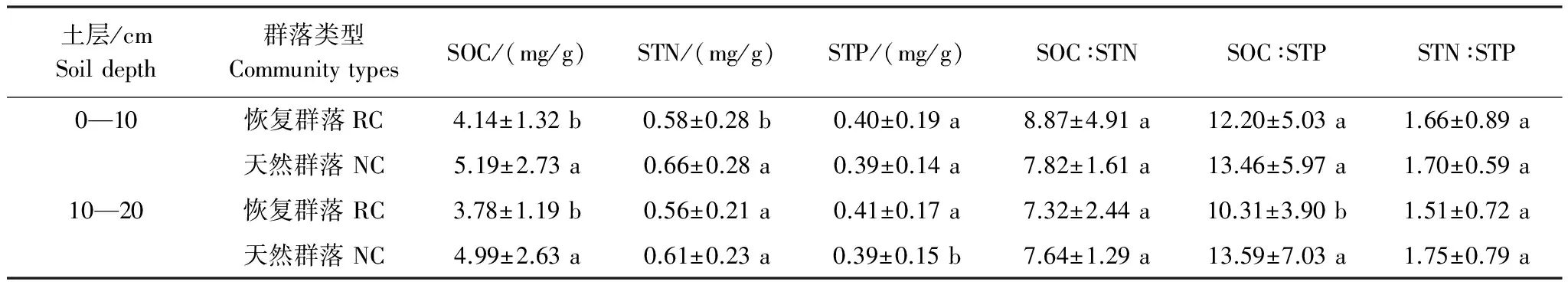
土层/cmSoildepth群落类型CommunitytypesSOC/(mg/g)STN/(mg/g)STP/(mg/g)SOC∶STNSOC∶STPSTN∶STP0—10恢复群落RC4.14±1.32b0.58±0.28b0.40±0.19a8.87±4.91a12.20±5.03a1.66±0.89a天然群落NC5.19±2.73a0.66±0.28a0.39±0.14a7.82±1.61a13.46±5.97a1.70±0.59a10—20恢复群落RC3.78±1.19b0.56±0.21a0.41±0.17a7.32±2.44a10.31±3.90b1.51±0.72a天然群落NC4.99±2.63a0.61±0.23a0.39±0.15b7.64±1.29a13.59±7.03a1.75±0.79a
SOC:土壤有机碳含量Soil organic carbon;STN:土壤全氮含量Soil total nitrogen;STP:土壤全磷含量 Soil total phosphorus;配对样本t检验,不同字母表示P<0.05水平上差异显著
表4 青藏高原高寒草地区恢复群落与天然群落土壤养分含量及化学计量比的垂直分布特征(平均值±标准偏差)
Table 4 Vertical distribution of soil C、N、P concentration and stoichiometric ratio of restorable plant communities at land used for construction highway and natural plant communities in the alpine grassland regions of Qinghai-Tibet Plateau(Mean ± SD)
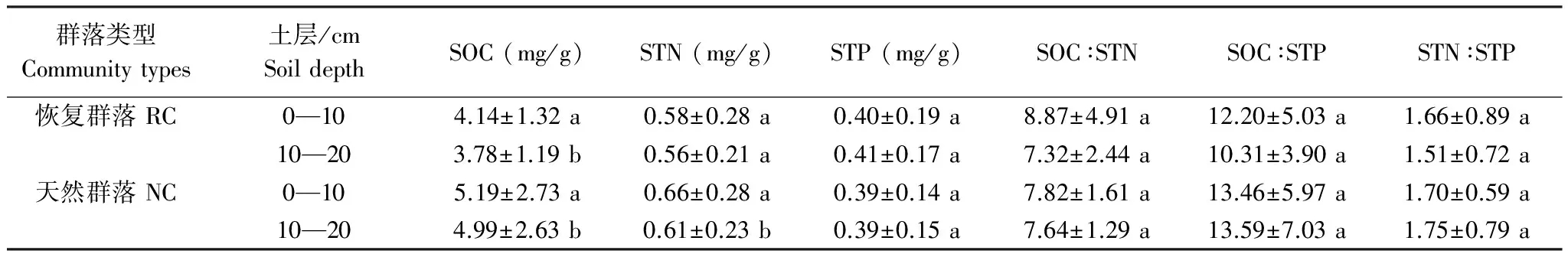
群落类型Communitytypes土层/cmSoildepthSOC(mg/g)STN(mg/g)STP(mg/g)SOC∶STNSOC∶STPSTN∶STP恢复群落RC0—104.14±1.32a0.58±0.28a0.40±0.19a8.87±4.91a12.20±5.03a1.66±0.89a10—203.78±1.19b0.56±0.21a0.41±0.17a7.32±2.44a10.31±3.90a1.51±0.72a天然群落NC0—105.19±2.73a0.66±0.28a0.39±0.14a7.82±1.61a13.46±5.97a1.70±0.59a10—204.99±2.63b0.61±0.23b0.39±0.15a7.64±1.29a13.59±7.03a1.75±0.79a
3.4 群落叶片和土壤C、N、P含量及计量比间的关系
Pearson相关性分析结果表明,植物群落叶片C含量与叶片C∶N、STN呈显著地正相关(表5);叶片N含量与叶片P含量呈显著正相关;叶片P含量与C∶P和N∶P呈极显著负相关,而与STP显著呈正相关;叶片C∶N与STN呈显著正相关;叶片C∶P与叶片N∶P呈显著正相关;叶片N∶P与STP呈显著负相关;土壤SOC与STN、SOC∶STP、STN∶STP呈显著正相关;土壤STN与SOC∶STP、STN∶STP呈极显著正相关;土壤STP与SOC:STP、STN:STP呈显著负相关。
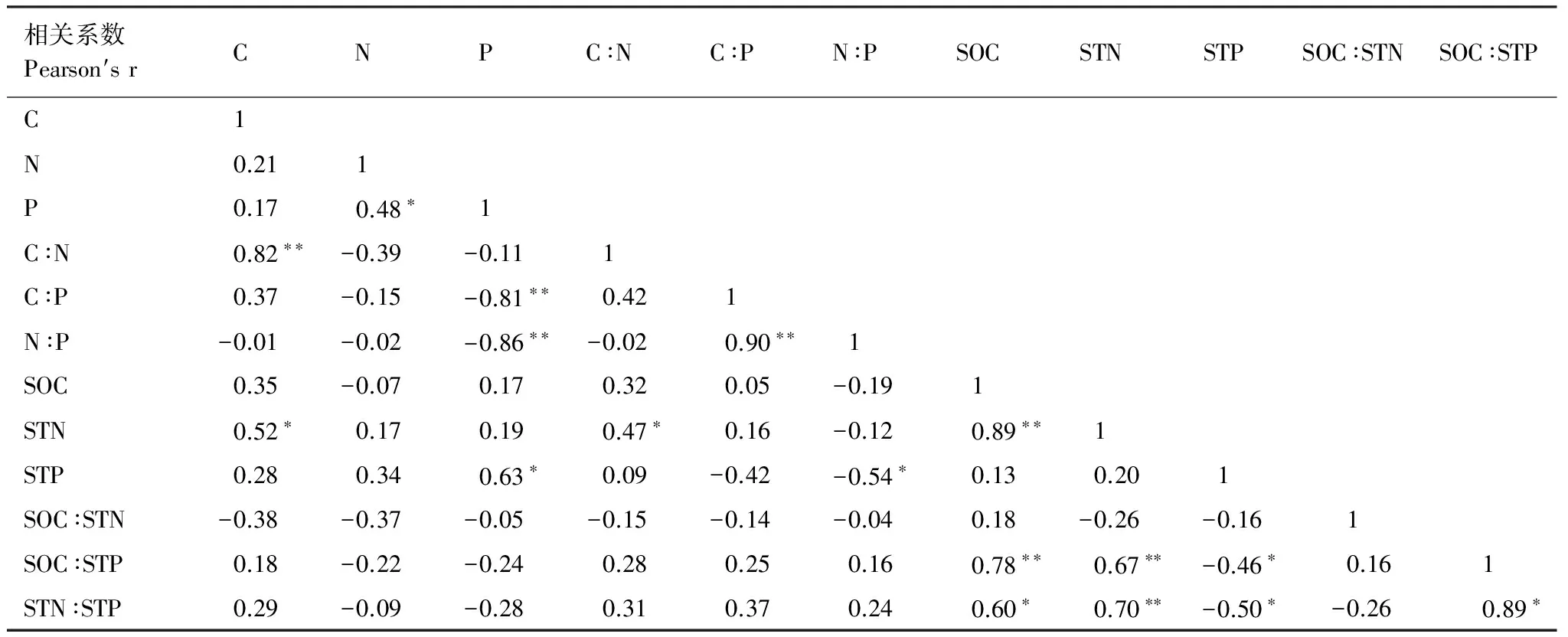
表5 群落植物叶片C、N、P含量及计量比和土壤养分含量及计量比之间的相关性Table 5 The correlations between community plant leaf stoichiometric variables and soil stoichiometry variables
Pearson相关性检验,数据为相关系数,*P<0.05;**P<0.01
4 讨论
4.1 筑路取土迹地对植物群落特征及化学计量学特征的影响
筑路取土迹地经历了18a无人工辅助的自然恢复过程,已经逐步实现植物群落的定居,植物群落的盖度、生物量和组分均显示出一定的自然恢复能力,但不同取样点恢复程度存在差异,有些调查点的恢复群落已经和天然群落享有相同的优群种,说明这些地区的筑路取土迹地恢复群落已经向毗邻天然草地群落的方向演变,如沱沱河、乌丽、西大滩等地,而有些调查点筑路取土迹地恢复群落和毗邻天然群落仅共享一定数量的主要伴生种,而优势种不同,如不冻泉、可可西里和乌丽,这说明这些地点的筑路取土迹地要么群落恢复缓慢,要么当前恢复群落就是稳定群落,其最终演变趋向经过近18a后仍然不明确。
恢复群落的演变趋向一定程度上决定于群落内植物组分的更替过程,而植物物种的更替受其生存环境中养分和水分等因素的限制,这是因为不同系统发育的植物对养分具有选择吸收的特性,因此筑路取土迹地植被与土壤C、N、P含量及其计量比很大程度上影响着取土迹地土壤和植被协同演变进程。恢复群落与天然群落叶片C含量没有显著差异,主要由于C是构成生物体的基本元素,其在植物体养分含量比例很大且较为稳定。恢复群落植物叶片N含量较高,主要原因可能是,一方面恢复群落内豆科植物种类及比例大于天然群落(主要是棘豆和黄芪),豆科植物具有固氮能力,从而提高了群落水平叶片的N含量;另一方面恢复群落盖度较天然群落小,植被夏季隔热能力相对减弱,土壤内温度相对高,有助于微生物将植物残体分解为无机物质,加速植物残体N向土壤归还的过程。本研究中恢复群落叶片特征存在着N含量升高C∶N比降低的模式,这与添加N肥(NH4NO3)能够提高受N限制的内蒙古草原植物叶片的N含量的结果本质上趋同[18],差异在于本研究是自然N素含量增加的过程,而内蒙古草原是人为添加N素含量的过程,然而当植物生长受P限制时,添加氮素并没有显著影响植物叶片N∶P[19],这说明不同N和P元素限制的环境内,N素增加对植物叶片的N含量及N∶P的影响不一致。P含量在天然和恢复群落间并未表现出显著差异,一方面主要原因是土壤速效磷供给受植被盖度影响较小,另外一方面是不同群落的植物对P素的利用能力不同,可能掩盖了工程迹地对植物叶片P含量的影响。
叶片C∶N和C∶P反映了植物吸收营养元素时所能同化C的能力,同时也反映了植物营养元素的利用效率[20]。恢复群落植物叶片N含量高于天然群落而C含量差异不大,由此导致了恢复群落较低的叶片C∶N,表明在光合能力和C储量能力差别不大的情况下,恢复群落的N养分利用效率较低,主要是因为恢复群落内豆科植物多,增强了植物群落有效态N的累积能力,这可能加速群落恢复到稳定态的速率,而恢复群落与天然群落C∶P差异不大。
4.2 恢复样地与天然样地的养分特征的比较
土壤养分的高低不仅反映了之前群落与土壤协同恢复的作用结果,也是草原植物养分需求的现实供给能力的体现,因此一定程度上预示着目前草原植物的生长状况和演替方向。植被生产力和有机质的矿化强度决定了土壤SOC含量,而它们又强烈的受到水热状况等影响[21],恢复样地在0—10cm和10—20cm的土层内的SOC各占对照的天然样地的80%和76%,说明恢复群落在土壤恢复方面并未达到天然群落的水平,但已经趋向正向演化。土壤N素主要以有机氮的形式存在于有机质中,恢复样地的STN含量只在0—10cm土层中表现出显著小于天然样地,而在10—20cm土层内没有差别,可能由于STN不仅与有机质的量有关,而且与有机质的分解强度有关。STP含量在恢复样地10—20cm土层中高于对应的天然样地,表明在深层土壤中天然样地比恢复样地更容易受到P缺乏的限制。工程迹地在深层土壤中拥有较高的STP含量,可能原因是工程迹地的地势一般低于周边未干扰草地,因此容易汇集来自天然草地土壤淋溶与地表径流而损失的P元素,另外筑路施工时铲去上层土壤导致土壤的风化速率发生不明确的影响也是潜在的原因。土壤SOC∶STN是土壤氮素矿化能力的标志,低的SOC∶STN可以加快微生物的分解和氮的矿化速率[22],本研究发现恢复群落和天然群落样地SOC∶STN比差别不大。STN∶STP是反映土壤内部氮磷循环和氮磷平衡的主要指标,有助于确定生态过程对筑路工程迹地的响应[23],本研究中恢复群落和天然群落土壤STN∶STP并未表现出显著差异。土壤SOC∶STP比是表征P有效性的一个指标,其中较低的比值对应着较高 P有效性,本研究表明恢复群落SOC∶STP一定程度上低于天然群落,工程迹地土壤P有效性较高。
工程迹地与天然样地的SOC在0—10cm土层的含量大于10—20cm土层的含量,STN有类似的趋势,而P含量在土层间差异不大,这种土层间的养分垂直分布格局主要与土壤养分来源的不同有关:土壤SOC和STN主要来源分别是凋落物的归还和有机质分解,因而土壤浅层较土壤深层具有较多的有机质和氮素来源,并且地表较高的温度加速了微生物分解有机质的能力[24],从而导致表层STN含量较深层STN含量增加;而P主要来源于土壤母质,是一个相对稳定的指标,因此两个土层间的差异不大。
当前的筑路工程迹地的草地-土壤系统经历了18a的自然协同恢复演替过程,其相比工程迹地形成初期,地表植被随着上层土壤的铲挖而移除并伴随着土壤养分的急剧下降的情况而言,尽管有机质和氮素养分未达到天然群落的水平,但已得到了明显改善,土壤肥力正发生着正向的演替。
4.3 叶片与土壤养分的含量和计量特征的关系
本研究中,恢复群落和天然群落植物叶片C含量与土壤STN表现出显著的正相关关系。叶片N含量与土壤STN相关性不显著,表明叶片N含量可能更倾向于是一种物种性状,而与土壤养分无关[25]。土壤STN与群落叶片N含量没有显著相关性,也解释了恢复群落土壤的STN比天然群落较低,但恢复群落叶片拥有较高的叶片N含量的看似矛盾的情况,说明STN并不是叶片N含量的决定因素。另外植物吸收的是移动性很强的可利用氮,并且研究区84%的年降雨发生在7月到9月的生长季节[1],导致这些移动性很强的硝态氮和氨态氮易随降水、淋溶径流迁移至因取土而高差较低的取土迹地地段,因此分布于取土迹地的恢复群落的生境拥有较为丰富的可吸收有效氮,可能也是造成恢复群落叶片较高的N含量的现实情况的原因之一,该过程与广东鼎湖山自然保护区,地势低处土壤硝态氮含量较高,是雨水冲刷携带增加所致的结果本质趋同[26]。另外,恢复群落植物叶片N含量较高和土壤STN含量较低,说明了筑路工程迹地提高了养分富集系数(植物营养元素与土壤元素含量的比值),加速了草地系统的N循环。通过植物的生物量的归还过程,恢复群落较高的叶片N含量对工程迹地土壤养分的恢复有积极意义,并且对加速群落演替和恢复到稳定态具有促进作用。群落叶片P含量与土壤STP含量显著地正相关,表明植物叶片P含量在一定程度上受到土壤环境中全磷的影响。
群落植物叶片N∶P与土壤STN没有显著的相关关系,原因可能是植物叶片N∶P的变化受到植物系统发育、气候因子、土壤养分有效性等因子共同驱动作用[27-28],土壤STN对群落叶片N∶P的影响被其它因子的作用抵消。而群落叶片N∶P与土壤STP含量表现出显著的负相关,与Güsewell和Koerselman[29]、丁凡等[25]的研究结果一致。
4.4 恢复群落与天然群落的氮磷养分限制格局
一般认为,植物叶片N∶P是植物对当地土壤环境的长期适应结果,反映了植物对生境养分供给的一种适应性,因此N∶P也通常作为判断植物养分受限状况的指示剂[30-32]。Koerselman and Meuleman[33]通过对欧洲湿地生态系统研究提出氮磷受限情况的判断阈值,即N∶P>16意味着植物生长受P的限制,N∶P<14反映植物生产力受N限制,比值在14与16之间则受N、P的共同制约限制,并且这对阈值已广泛地应用到养分受限状况的判定研究中[28,34],然而群落的养分限制性状况受控于物种的元素利用策略等多种因素的共同制约,植物生长营养受限的判定阈值应随地区不同或物种组成不同而变化[14,35]。青藏高原北部群落水平植物叶片N∶P比值为14.5,按照上述阈值判断,本地区植物生长受到N、P的共同限制。
Garnier[36]指出当植物受到某元素限制时,该元素在叶片中的含量会与土壤中对应的元素的供给能力呈正相关关系。本研究结果得出,群落叶片N含量与土壤STN没有显著相关关系,而群落叶片P含量与土壤STP含量显著地正相关,说明本地区植物的生长不受STN的限制而受到STP的限制。这与前面通过养分受限的N∶P判断阈值得出的本地区植物生长受到N、P共同限制的结论不一致,其原因可能是本研究区域潜在更低的N、P受限状况的判定阈值。Han等[37]指出中国陆地植物生长普遍受到P元素的限制的原因是,中国土壤P浓度为561 μg/kg低于全球平均水平可能导致了中国植物叶片低的P含量,而本研究地区土壤STP更低,因此植物受P元素的限制更强烈。工程迹地恢复群落叶片N∶P为15.97而天然群落的N∶P为13.08,而较高的N∶P比值反映了植被受到P限制更强烈,所以筑路工程迹地草地恢复群落比天然群落更容易受到P元素的限制。虽然本研究结果表明,研究地区草地植被主要受到P元素的限制,但青藏高原地区植物究竟是受N元素限制,还是P元素限制,目前仍然没有共识。Liu等[38]在青藏高原纳木措地区通过两年氮肥(NH4NO3)添加实验,发现非豆科植被主要受N的限制。杨晓霞等[39]在青海海北高寒草地生态系统研究站采用N、P添加实验,发现青藏高原高寒草甸植物生长可能受土壤可利用P的限制。而刘旻霞等[40]的研究结果表明,青藏高原东北缘高寒草甸地区杂草在阴坡倾向于N限制而在阳坡倾向于P限制。Liu等和杨晓霞等的研究通过人为添加元素,改变了研究地区自然状态下植物与土壤养分间的关系,而本研究主要关注天然群落和取土迹地恢复群落的差异,因此可能是由于不同研究所处的地点、研究对象和采用的手段存在差异,导致青藏高原地区草地植物N、P计量研究的结果出现分异。
5 结论
实地调查结果表明,青藏高原北部地区筑路取土迹地经历18a自然恢复后,筑路取土迹地均已逐步实现植物群落的定居,但其恢复程度存在明显差异。恢复群落植物叶片N含量较高,C∶N较低,表明恢复群落植物的N利用效率较低,主要是恢复群落内豆科植物比例较高和土壤有效N供应充足所致。虽然恢复群落的土壤恢复度尚未达到天然群落水平,但土壤肥力正发生正向演替。研究地区草地植被主要受到P元素的限制,且工程迹地草地恢复群落比天然群落容易受到P元素的限制。
[1] Guo Z G, Long R J, Niu F J, Wu Q B, Hu Y K. Effect of highway construction on plant diversity of grassland communities in the permafrost regions of the Qinghai-Tibet plateau. The Rangeland Journal, 2007, 29(2):161- 167.
[2] 毛亮, 周杰, 郭正刚. 青藏高原高寒草原区工程迹地面积对其恢复植物群落特征的影响. 生态学报, 2013, 33(11):3547- 3554.
[3] Cui X F, Graf H F. Recent land cover changes on the Tibetan Plateau:a review. Climatic Change, 2009, 94(1- 2):47- 61.
[4] 李东明, 郭正刚, 安黎哲. 青藏高原多年冻土区不同草地生态系统恢复能力评价. 应用生态学报, 2008, 19(10):2182- 2188.
[5] Jin H J, Yu Q H, Wang S L, Lü L Z. Changes in permafrost environments along the Qinghai-Tibet engineering corridor induced by anthropogenic activities and climate warming. Cold Regions Science and Technology, 2008, 53(3):317- 333.
[6] Korb J E, Johnson N C, Covington W W. Slash pile burning effects on soil biotic and chemical properties and plant establishment:recommendations for amelioration. Restoration Ecology, 2004, 12(1):52- 62.
[7] Bradshaw A. Restoration of mined lands-using natural processes. Ecological Engineering, 1997, 8(4):255- 269.
[8] Wang G X, Yao J Z, Guo Z G, Wu Q B, Wang Y B. Changes in permafrost ecosystem under the influences of human engineering activities and its enlightenment to railway construction. Chinese Science Bulletin, 2004, 49(16):1741- 1750.
[9] 马世震, 陈桂琛, 彭敏, 周国英, 赵以莲. 青藏公路取土场高寒草原植被的恢复进程. 中国环境科学, 2004, 24(2):188- 191.
[10] 郭正刚, 刘慧霞, 王根绪, 程国栋. 人类工程对青藏高原北部草地群落β多样性的影响. 生态学报, 2004, 24(2):384- 388.
[11] 郭正刚, 牛富俊, 湛虎, 吴青柏. 青藏高原北部多年冻土退化过程中生态系统的变化特征. 生态学报, 2007, 27(8):3294- 3301.
[12] 孔亚平, 陈济丁, 辛有俊. 青藏公路多年冻土路段沿线植被及其变化. 公路, 2008, (3):179- 184.
[13] Sterner R W, Elser J J. Ecological Stoichiometry:the Biology of Elements from Molecules to the Biosphere. Princeton, NJ:Princeton University Press, 2002.
[14] Zhang L X, Bai Y F, Han X G. Differential responses of N∶P stoichiometry ofLeymuschinensisandCarexkorshinskyito N additions in a steppe ecosystem in Nei Mongol. Acta Botanica Sinica, 2004, 46(3):259- 270.
[15] 孙海松. 青海高寒草地资源及其永续利用. 草业科学, 1998, 15(3):1- 3.
[16] 王一博, 王根绪, 程玉菲, 李元寿. 青藏高原典型寒冻土壤对高寒生态系统变化的响应. 冰川冻土, 2006, 28(5):633- 641.
[17] 郑淑霞, 上官周平. 黄土高原地区植物叶片养分组成的空间分布格局. 自然科学进展, 2006, 16(8):965- 973.
[18] Cui Q, Lü X T, Wang Q B, Han X G. Nitrogen fertilization and fire act independently on foliar stoichiometry in a temperate steppe. Plant and Soil, 2010, 334(1- 2):209- 219.
[19] Kozovits A R, Bustamante M M C, Garofalo C R, Bucci S, Franco A C, Goldstein G, Meinzer F C. Nutrient resorption and patterns of litter production and decomposition in a Neotropical Savanna. Functional Ecology, 2007, 21(6):1034- 1043.
[20] Herbert D A, Williams M, Rastetter E B. A model analysis of N and P limitation on carbon accumulation in Amazonian secondary forest after alternate land-use abandonment. Biogeochemistry, 2003, 65(1):121- 150.
[21] Jobbágy E G, Jackson R B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications, 2000, 10(2):423- 436.
[22] Springob G, Kirchmann H. Bulk soil C to N ratio as a simple measure of net N mineralization from stabilized soil organic matter in sandy arable soils. Soil Biology and Biochemistry, 2003, 35(4):629- 632.
[23] 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8):3937- 3947.
[24] Schuerings J, Beierkuhnlein C, Grant K, Jentsch A, Malyshev A, Peuelas J, Sardans J, Kreyling J. Absence of soil frost affects plant-soil interactions in temperate grasslands. Plant and Soil, 2013, 371(1- 2):559- 572.
[25] 丁凡, 廉培勇, 曾德慧. 松嫩平原草甸三种植物叶片N、P化学计量特征及其与土壤N、P浓度的关系. 生态学杂志, 2011, 30(1):77- 81.
[26] 黄文娟, 刘菊秀, 唐旭利, 黄钰辉, 刘世忠, 褚国伟, 周国逸. 鼎湖山5种森林土壤的无机氮和有效磷含量. 应用与环境生物学报, 2009, 15(4):441- 447.
[28] Townsend A R, Cleveland C C, Asner G P, Bustamante M M C. Controls over foliar N∶P ratios in tropical rain forests. Ecology, 2007, 88(1):107- 118.
[29] Güsewell S, Koerselman W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology, Evolution and Systematics, 2002, 5(1):37- 61.
[30] Aerts R, Chapin F S III. The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30:1- 67.
[31] Güsewell S. N∶P ratios in terrestrial plants:variation and functional significance. New Phytologist, 2004, 164(2):243- 266.
[32] Frank D A. Ungulate and topographic control of nitrogen:phosphorus stoichiometry in a temperate grassland; soils, plants and mineralization rates. Oikos, 2008, 117(4):591- 601.
[33] Koerselman W, Meuleman A F M. The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6):1441- 1450.
[34] Tessier J T, Raynal D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. Journal of Applied Ecology, 2003, 40(3):523- 534.
[35] Chen F S, Niklas K J, Zeng D H. Important foliar traits depend on species-grouping:analysis of a remnant temperate forest at the Keerqin Sandy Lands, China. Plant and Soil, 2011, 340(1- 2):337- 345.
[36] Garnier E. Interspecic variation in plasticity of grasses in response to nitrogen supply // Population Biology of Grasses. Cambridge:Cambridge University Press, 1998:155- 155.
[37] Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2):377- 385.
[38] Liu Y W, Xu X L, Wei D, Wang Y H, Wang Y S. Plant and soil responses of an alpine steppe on the Tibetan Plateau to multi-level nitrogen addition. Plant and Soil, 2013, 373(1- 2):515- 529.
[39] 杨晓霞, 任飞, 周华坤, 贺金生. 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应. 植物生态学报, 2014, 38(2):159- 166.
[40] 刘旻霞, 朱柯嘉. 青藏高原东缘高寒草甸不同功能群植物氮磷化学计量特征研究. 中国草地学报, 2013, 35(2):52- 58.
C, N, and P stoichiometry of plant and soil in the restorable plant communities distributed on the Land Used for Qinghai-Tibet Highway Construction in the
Qinghai-Tibetan Plateau, China LUO Mingwei, MAO Liang, LI Qianqian, ZHAO Xu, XIAO Yu, JIA Tingting, GUO Zhenggang*
TheStateKeyLaboratoryofGrasslandAgro-ecosystems,CollegeofPastoralAgriculturalScienceandTechnology,LanzhouUniversity;P.O.Box61,Lanzhou730020,China
The restoration of Land Used for Qinghai-Tibet Highway Construction is an important part to restore degraded alpine grassland in the Qinghai-Tibet Plateau, China. C, N, P stoichiometry is considered an effective tool to disclose the synergistic adaptation of plant growth and element availability in the process of plant restoration on Land Used for Qinghai-Tibet Highway Construction. A field survey was carried out to investigate whether the Land Used for Qinghai-Tibet Highway Construction contributes to shifts in the C, N, and P stoichiometry of the leaves and soil of the plant community, and to identify the N- or P- limitation status of plants in the natural community and restorable communities growing for nearly two decades as part of the natural regeneration of Land Used for Qinghai-Tibet Highway Construction. Seven survey sites were selected along the Qinghai-Tibet Highway, in which four groups of restorable and natural plots were arranged. Then, four subplots for each plot were randomly used to harvest community leaves and soil samples. Organic C, N, and P concentrations of plant leaves and soil samples were analyzed by using the external heating method, the Kjeldahl acid-digestion method, and the Mo-Sb-Ascorbic acid colorimetric method, respectively. The independent-sample t test and paired t test were used to analyze differences in leaf C, N, and P stoichiometric variables and differences in soil organic C, N, and P stoichiometric variables from restorable and natural plots, respectively. The relationship between leaf and soil C, N, and P stoichiometric variables was analyzed by Pearson correlation. This study showed that the plant communities had gradually established on the Land Used for Qinghai-Tibet Highway Construction at each study site, but that the plant composition was different between restorable communities and natural communities after 18 years natural restoration. The leaf N concentration of restorable communities was higher than that of natural communities; thus, inducing lower leaf C∶N, which showed that the N nutrient-use efficiency of the restorable communities was lower than that of the natural communities. A high percentage legume plant and relatively adequate available N supplement may be the major reasons for lower leaf C∶N in restorable communities. The soil organic carbon (SOC) of the restorable plots at both 0—10cm and 10—20cm soil depths were significantly lower than those of natural plots, and the soil total nitrogen (STN)shared the similar pattern with SOC. The soil total P of the restorable plots in the 10—20cm soil layer was higher than that in natural plots. These results indicate that the soil fertility of restorable communities did not reach the level of natural communities, but that it had improved significantly. Leaf N and STN had no significant correlation; however, leaf P was significantly positively correlated with soil total phosphorus (STP), showing that leaf P is influenced by STP to some certain extent. Using the thresholds of N, P-limitation, and the results of the correlation between leaf elements and soil nutrients, this study shows that plants are probably P-limited in the study region, with plants in the restorable communities suffering from more intense P-limited conditions than those in the natural communities.
lands used for construction; C, N, P; stoichiometry; alpine grassland; restorable community
国家行业公益资助项目(201203041); 国家科技支撑资助项目(2012BAD13B05); 教育部创新团队(IRT13019);兰州大学中央高校基本科研业务费(lzujbky-2014-m01)
2014- 04- 30; < class="emphasis_bold">网络出版日期:
日期:2015- 05- 18
10.5846/stxb201404300866
*通讯作者Corresponding author.E-mail:guozhg@lzu.edu.cn
雒明伟, 毛亮, 李倩倩, 赵旭, 肖玉, 贾婷婷, 郭正刚.青藏高原筑路取土迹地恢复植物群落与土壤的碳氮磷化学计量特征.生态学报,2015,35(23):7832- 7841.
Luo M W, Mao L, Li Q Q, Zhao X, Xiao Y, Jia T T, Guo Z G.C, N, and P stoichiometry of plant and soil in the restorable plant communities distributed on the Land Used for Qinghai-Tibet Highway Construction in the Qinghai-Tibetan Plateau, China.Acta Ecologica Sinica,2015,35(23):7832- 7841.
猜你喜欢
兰台内外(2021年33期)2021-12-17 15:41:02
现代园艺(2021年23期)2021-12-01 07:47:44
云南画报(2020年12期)2021-01-18 07:19:08
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
军民两用技术与产品(2018年11期)2018-07-16 11:49:24
现代农村科技(2018年6期)2018-01-18 06:09:22
山西建筑(2017年15期)2017-07-01 22:03:22
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
筑路机械与施工机械化(2014年10期)2014-03-01 03:02:25
