
三峡水库奉节以东秭归和巫山段消落带植物群落动态特征
2015-02-11 02:06:28朱妮妮郭泉水秦爱丽裴顺祥马凡强简尊吉
生态学报 2015年23期
朱妮妮,郭泉水,* ,秦爱丽,裴顺祥,马凡强,朱 莉,3,简尊吉
1 中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室, 北京 100091;2 中国林业科学研究院华北林业实验中心,北京 102300 3 河南科技大学农学院,洛阳 471003
三峡水库奉节以东秭归和巫山段消落带植物群落动态特征
朱妮妮1,郭泉水1,*,秦爱丽1,裴顺祥2,马凡强1,朱 莉1,3,简尊吉1
1 中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室, 北京 100091;2 中国林业科学研究院华北林业实验中心,北京 102300 3 河南科技大学农学院,洛阳 471003
基于2008—2012 年对三峡水库奉节以东秭归和巫山段消落带固定样地不同海拔区段植物群落的5a定位监测,研究消落带植物群落的物种组成、优势植物、植物生活型和物种多样性的动态变化,结果表明:1)截止2012年,消落带海拔156—172 m区段共经历了4 次水库水位涨落。经历首次后(2009 年),消落带原生植物由55 科147 种减少到18 科33 种,经历4 次后(2012 年),减少到14 科39 种。与经历水库水位涨落前(2008 年)比较,经历首次后的科数减少了67.3%,种数减少了77.6%;经历4 次后的科数减少了74.5%,种数减少了73.5%。在消落带原生植物减少的同时,出现了许多“新”植物。经历首次后出现了49 种,经历4 次后出现了23 种,分别占调查当年样地植物种类总数的59.8%和32.9%。海拔172—175 m区段共经历了2 次水库水位涨落,消落带原生植物由40科91 种(2008 年)减少到了13 科20种。与经历水库水位涨落前比较,科数减少了67.5%,种数减少了78.0%。出现“新”植物21 种,约占调查当年样地植物种类总数的44.7%。通过对历次调查中消落带植物“消失”和“出现”的数量比较表明,消落带植物对经历首次水库水位涨落的反应最为敏感,此后,虽又经历过几次水库水位涨落,但其变化速率趋于减小。2)不同海拔区段、不同生态适应型植物的“消长”动态和优势种组成不完全相同。海拔156—172 m区段,经历4 次水库水位涨落后,在消落带植物群落中占优势的草本植物种为菊科(Compositae)的鬼针草(Bidenspilosa)、禾本科(Gramineae)的狗牙根(Cynodondactylon)、毛马唐(Digitariachrysoblephara)、狗尾草(Setariaviridis)、莎草科(Cyperaceae)的碎米莎草(Cyperusiria),占优势的灌木树种为漆树科(Anacardiaceae)的盐肤木(Rhuschinensis)和大戟科(Euphorbiaceae)的算盘子(Glochidionpuberum);在海拔172—175 m区段,除鬼针草、毛马唐仍为优势种外,还增加了大戟科的湖北算盘子(Glochidionwilsonii),马鞭草科(Verbenaceae)的黄荆(Vitexnegundo),葡萄科(Vitaceae)的五叶地锦(Parthenocissusquniquefolia)等树种。3)消落带植物群落的优势生活型为一年生和多年生草本;物种多样性随着水库水位涨落次数的增加总体变化呈减少趋势。4)三峡水库水位周期性涨落导致消落带发生水陆环境交替变化,不同生态适应型植物对变化生境的适应能力有所不同,是消落带植物群落发生变化的主要驱动因素。
群落动态;植物生活型;物种多样性;三峡水库;消落带
三峡库区消落带相对高差30m,总面积348.93 km2,是目前我国相对高差和面积最大的水库消落带。受水库水位周期性涨落的影响,现已成为库区生态系统中物质转化、能量流动、信息传输与转换等最为活跃且最不稳定的生态脆弱带[1]。消落带植物群落是消落带生态系统的重要组成部分,在维护消落带生态系统稳定和生态功能发挥等方面起着重要作用,也是水库管理的重要对象[2-3]。及时准确掌握消落带植物群落动态,对于深入了解三峡水库消落带生态环境的变化以及消落带植被恢复和重建都具有重要的理论和实践意义。
植物群落的物种组成和结构变化是植物群落演变的具体表现特征[4]。近年来,许多学者围绕三峡库区消落带植物群落结构(如种类组成、物种丰富度或多样性、优势植物种群结构等)的变化和群落演替开展了广泛的调查和研究。熊平生等[5]、王勇等[6]和白宝伟[7]等基于植物群落演替理论和三峡自然消落带的植被分析,对消落带植物群落动态进行了推测;卢志军等[8]通过固定样方调查,对被水淹过的海拔156 m以下区段和当年未被水淹区段的植物种类组成、物种多样性和草本层生物量进行过比较;胡波等[1-2, 9-10]采用“空间推时间”的方法,对消落带植物群落的演替趋势进行了预测;郭泉水等[3]通过固定样地调查,研究了三峡库区消落带陆生植被首次经历水陆生境变化的响应。研究结论一致表明:受三峡水库水位涨落的影响,消落带植物的生存条件发生了巨大改变;消落带植物群落的正常发育和演替过程已受到严重干扰;不同植物因适应水陆交替变化生境的能力不同,其“消长”动态表现各异。这些研究提供了许多关于消落带植物群落动态变化的信息,对于深入揭示消落带动态变化规律具有一定的理论参考价值,不足之处在于:(1)以往研究大多集中在水库蓄水位达到海拔175 m之前,而对达到海拔175 m以后植物群落的调查研究较少。三峡水库正常运行的水位在海拔145—175 m,因此,其研究结果大多反映的是消落带局部地段植物群落的变化。(2)大多数研究采用的是“一次性调查”或“空间推时间”的方法,对于经历多次水库水位涨落持续干扰下消落带植物群落动态变化的定位研究还很少涉足,特别是缺乏对三峡库区奉节以东地区消落带植物群落动态的持续监测和研究。经历水库水位涨落后,在消落带上出现的大多是森林群落次生演替早期的先锋植物[3],这些植物种的“消长”波动性很大。“一次性调查”或以此调查为基础采用“空间推时间”的方法,对消落带植物群落动态变化趋势预测的结果必然会存在较大的不确定性。三峡库区的地势东部高,西南部低。奉节位于库区东部和西南部的分界处。奉节以东库岸以中山山地为主[11],为峡谷地貌。消落带的坡度大,水土流失严重,为三峡库区滑坡、崩塌等地质灾害多发区。地形是区域生态环境的一个重要因子。地形不同,气候、土壤、植被等资源及其分布以及人类对资源的利用方式和程度都会有所不同[11]。生物多样性是环境的产物。受不同生态环境因子及其生态过程的综合影响,消落带植物群落的组成、结构及其变化过程都会有所不同[12-14]。而目前尚未见针对三峡水库奉节以东峡谷地貌区消落带植物群落动态的研究报道。
本文以2008 年在三峡水库奉节以东巫山和秭归段典型消落带上设置的4 块固定监测样地为依托,在获得经历水库水位涨落前消落带植物群落物种组成和结构等初始资料的基础上,区分海拔156—172 m和172—175 m区段,对消落带植物群落的种类组成及不同植物的“消长”动态、植物生活型和物种多样性的变化进行了为期5a的定位监测,旨在通过对多年定位观测结果的比较和分析,揭示三峡水库奉节以东地区消落带植物群落在水库水位涨落持续干扰下的动态变化规律和发展趋势,为全面评估三峡水库运行对消落带植物群落的影响以及消落带植被恢复和重建中的物种选择提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于三峡水库奉节以东干流的秭归段和巫山段。其典型的地理特征是:水库干流两侧山体的海拔多在1200m左右。山势陡峭,长江“三峡”中著名的西陵峡和巫峡两个最大的峡即位于此。消落带的坡度一般在25°以上;2/3区段的基岩为石灰岩,只有秭归段部分地段分布着花岗岩。研究区的地形和地质条件在三峡水库奉节以东峡谷地貌区具有一定的代表性。
固定样地设在秭归县茅坪镇和巫山县巫峡镇消落带内。两地气候类型一致,均属中亚热带湿润季风气候。其中,秭归县茅坪镇的年均温18 ℃,年均降雨量1 049.3 mm,≥10℃年活动积温 5 723.6 ℃,全年无霜期305 d;巫山县巫峡镇年均温18.4 ℃,年均降雨量1 100mm,≥10℃ 年活动积温 5 857.3 ℃,全年无霜期306 d。地带性植被类型也一致,均为亚热带常绿阔叶林。但由于人类的长期干扰,在水库蓄水前消落带的地带性植被已被全部破坏,取而代之的是人类干预下的各种次生植被和人工植被。为了较全面反映研究区的植被特征,固定样地从在研究区有一定代表性的次生植被和人工植被中选择。各样地经历水库水位涨落前的自然概况见表1。
消落带水陆交替变化与水库水位涨落同步。不同年份水库水位涨落变化有所不同。2008 年冬季水库最高蓄水位首次达到海拔172 m,此前,最高蓄水位为海拔156 m。2009 年与2008 年相同。2010年冬季水库最高蓄水位上升至海拔175 m,此后,其最高水位一直维持在该海拔高度上运行。截止2012年8月,海拔156—172 m区段共经历了4 次水库水位涨落,海拔172—175 m区段共经历了2次。研究时段内不同年份的水库水位变化见图1[15]。
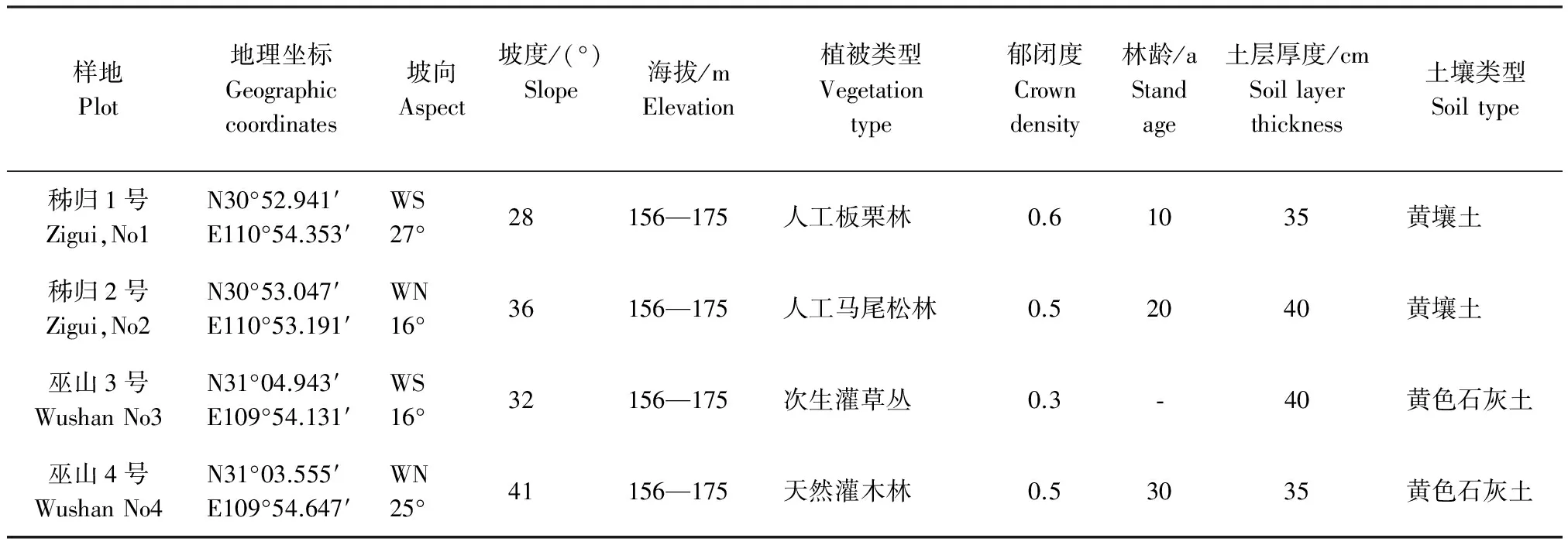
表1 样地自然概况Table 1 Natural conditions of the sample plot
2008年秭归1号和2号样地的板栗和马尾松被砍伐
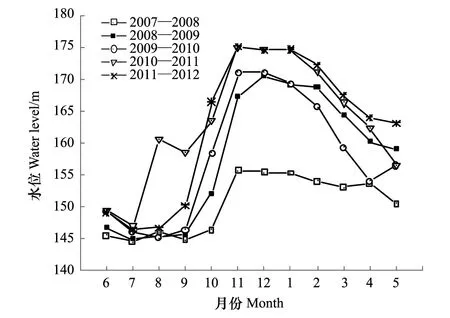
图1 三峡水库水位变化(2007—2012年) Fig.1 Water-level fluctuation of the Three Gorges Reservoir(2007—2012)
1.2 样地设置和调查
固定样地设置于2008 年8 月。在巫山和秭归消落带内各设置2 块。巫山的2 块样地设在水库干流左右两岸,秭归的2 块样地均设在水库干流的右侧。各样地的宽度和长度的投影距离均为15 m和19 m。宽边设在海拔156 m(2008 年水库最高蓄水位),上线设在海拔175 m。样方设置采用相邻格子法。样方大小因调查对象而异。乔木幼苗幼树和灌木样方为2 m × 2 m,草本为1 m × 1 m。调查内容包括:植物种类,不同植物的植株数量、平均高度、盖度和植物生活型。其中,丛生草本和灌木植株数量调查的方法是:以地面以上是否为单株存在为判别标准,至于其地面以下植株与植株之间或不同植株的根系之间是否连接不予考虑。
固定样地设置后,沿样地4边,每隔投影距离5 m,埋1 根水泥桩,并在顶端涂上红漆,以做长期观测的永久标记。调查时间分别为2008年8月、2009年8月、2012年8月。2008年8月首次调查,获取海拔156—175 m区段经历水库水位涨落前消落带植物群落的初始资料,2009年8月和2012年8月对经历首次和4 次水库水位涨落后植物群落的动态进行了两次监测调查。
1.3 数据处理与分析
由于消落带海拔156—172 m区段和海拔172—175 m区段经历的水库水位涨落次数不同,为此,在资料整理时,将两个海拔区段作为两个独立的统计单元进行处理。
统计指标包括:不同植物在不同调查年份的生命表征、植物生活型组成、重要值、群落的物种丰富度、Shannon-Weiner指数、Simpson指数、Pielou均匀度指数。其中,消落带植物生活型划分参考《中国植被》[16]的生活型分类系统,共区分出乔木、灌木、藤本、多年生草本、一年生草本等5种生活型。植物重要值、物种丰富度及多样性指数参考有关文献[17-18]提供的计算公式计算。
数据整理、计算和制图均采用Excel 7.0和Origin 9软件完成。
2 结果与分析
2.1 消落带植物群落种类组成及其动态变化
2.1.1 消落带植物群落不同植物的“消长”变化
区分海拔156—172 m和172—175 m两个区段,按不同调查年份,对4 块固定样地中的不同植物在样地中出现和“消失”的调查记录分别统计,结果见表2。
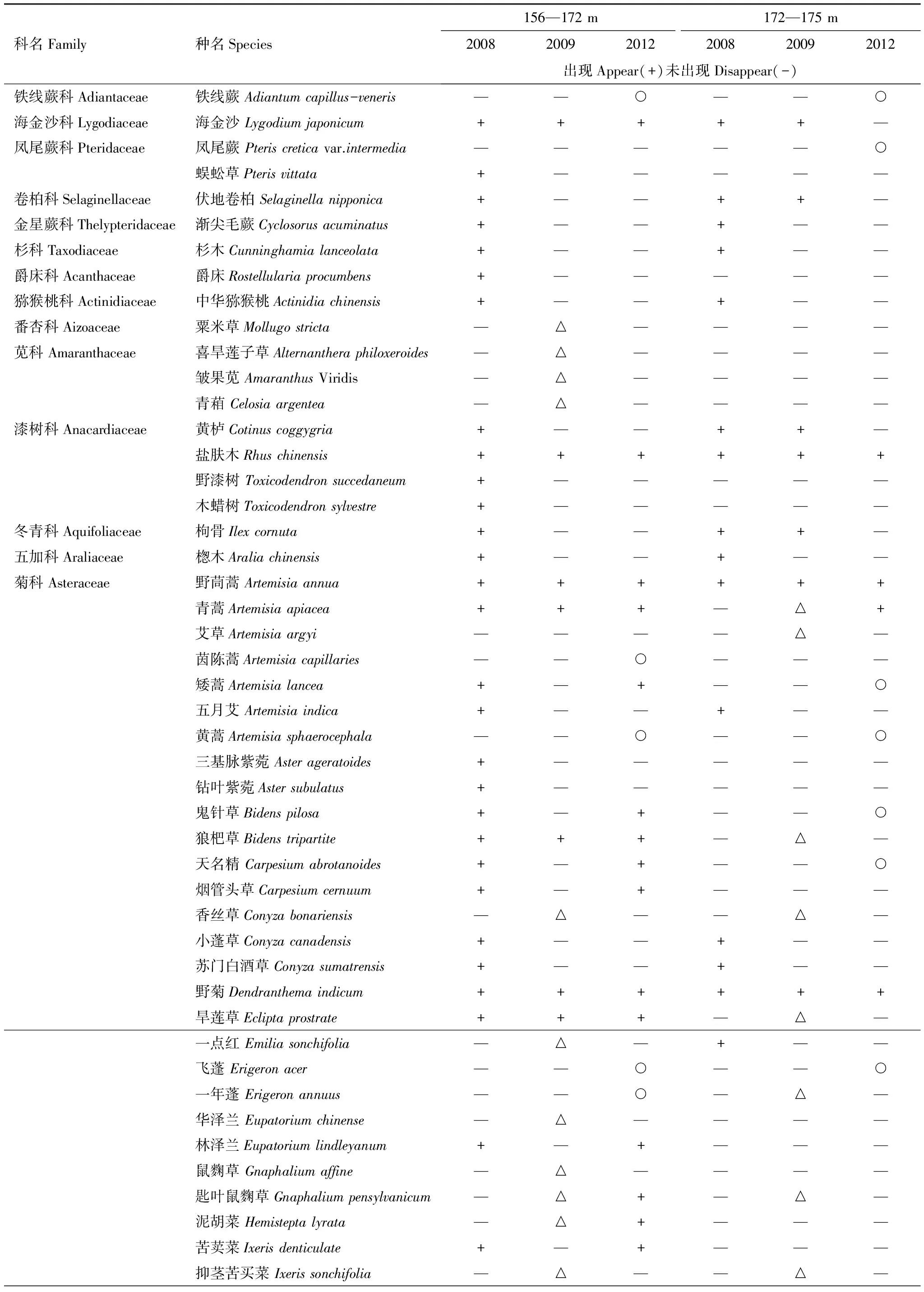
表2 2008—2012年三峡水库消落带植物的消长动态Table 2 Plant community "growth" dynamics in Hydro-fluctuation Belt of the Three Gorges Reservoir(2008—2012)
* 表中△表示在2008年未出现,2009年新出现的植物; ○表示2008年和2009年均未出现,仅在2012年出现的植物
从表2可概括出2008—2012年消落带植物群落种类组成变化的基本特征如下:
(1)原来生长在消落带的大多数植物经历水库水位涨落后消失,同时也有许多“新”植物出现。不同植物的消长过程不同,有些种经历首次水位涨落后即“永久消失”,如棕榈(Trachycarpusfortunei)、毛葡萄(Vitisheyneana)以及多种乔灌木;有些种在经历首次水库水位涨落后 “消失”,但经历多次水位涨落后又有出现,如矮蒿(Artemisialancea),苎麻(Boehmerianivea)等; 也有一些种在样地上始终存在,如野茼蒿(Artemisiaannua)、野菊(Dendranthemaindicum)等。不同植物的消长动态,反映了不同生态适应型植物对消落带水陆变化生境响应程度上的差异。
(2)不同调查年份、不同海拔区段消落带植物群落的数量特征随着经历水库水位涨落次数、淹水深度和持续水淹时间长短而变化。经历水库水位涨落前(2008年8月),在海拔156—172 m区段共调查到原生植物55 科147 种。经历首次水库水位涨落后(2009年8月),减少到18 科33 种。经历4 次水库水位涨落后(2012年8月),减少到14 科39 种。与2008年比较,2009年和2012年科数分别减少了67.3%和74.5%,种数分别减少了77.6%和73.5%。对不同调查年份科、种的变化速率的比较发现,消落带植物对首次经历水库水位涨落的反应最为敏感,而保存下来的植物虽又经历过几次水库水位涨落,但其变化速率趋于减小。如2009—2012年这3年间,原生植物的科数仅减少了22.2%,与经历首次水库水位涨落后的变化(减少了67.3%)明显不同。
在海拔172—175 m区段,2008年8月调查到原生植物40科91 种,2009年8月减少到24科37 种,2012年8月减少到13科20种。与2008年比较,2009年和2012年的科数分别减少了40.0%和67.5%,种数减少了59.3%和78.0%。该区段从2010年冬季开始受水库水位涨落影响,截止2012年8月,仅经历了2 次水库水位涨落的影响。2008年8月到2009年8月植物科、种的变化,属自然环境下植物种群的消长过程。将此期间的变化率(40.0%)与经历2次水库水位涨落影响下的变化率(67.5%)比较,凸显了水库水位涨落对消落带植物种群的强烈干扰作用。将该区段科数和种数减少的幅度与海拔156—172 m区段比较可见,海拔172—175 m区段的科数减少的幅度小,而种数减少的幅度相差无几。反映出经历水库水位涨落次数、淹水深度、水淹持续时间等对消落带植物群落不同的影响效应。
(3)在海拔156—172 m区段,经历首次水库水位涨落后(2009年8月),共记录到49 种“新”植物,约占调查当年样地植物种类总数的59.8%;经历4 次水库水位涨落后(2012年8月),为 23 种,约占调查当年样地上植物种类总数的32.9%。该结果显示,消落带上“新”植物也是以经历首次水位涨落后出现最多,而随着经历水库水位涨落次数的增加,“新”出现的植物也在逐渐减少。在海拔172—175 m区段,出现了21 种“新”植物,约占调查当年样地上植物种总数的44.7%。明显小于海拔156—172 m区段经历首次水库水位涨落后“新”植物在群落中所占的比例(59.8%)。
2.1.2 消落带的优势植物及其变化
对2008—2012年不同调查年份、不同植物的重要重要值分别计算,并提取在此期间某个调查年份植物重要值在10以上的植物种类列如表3。
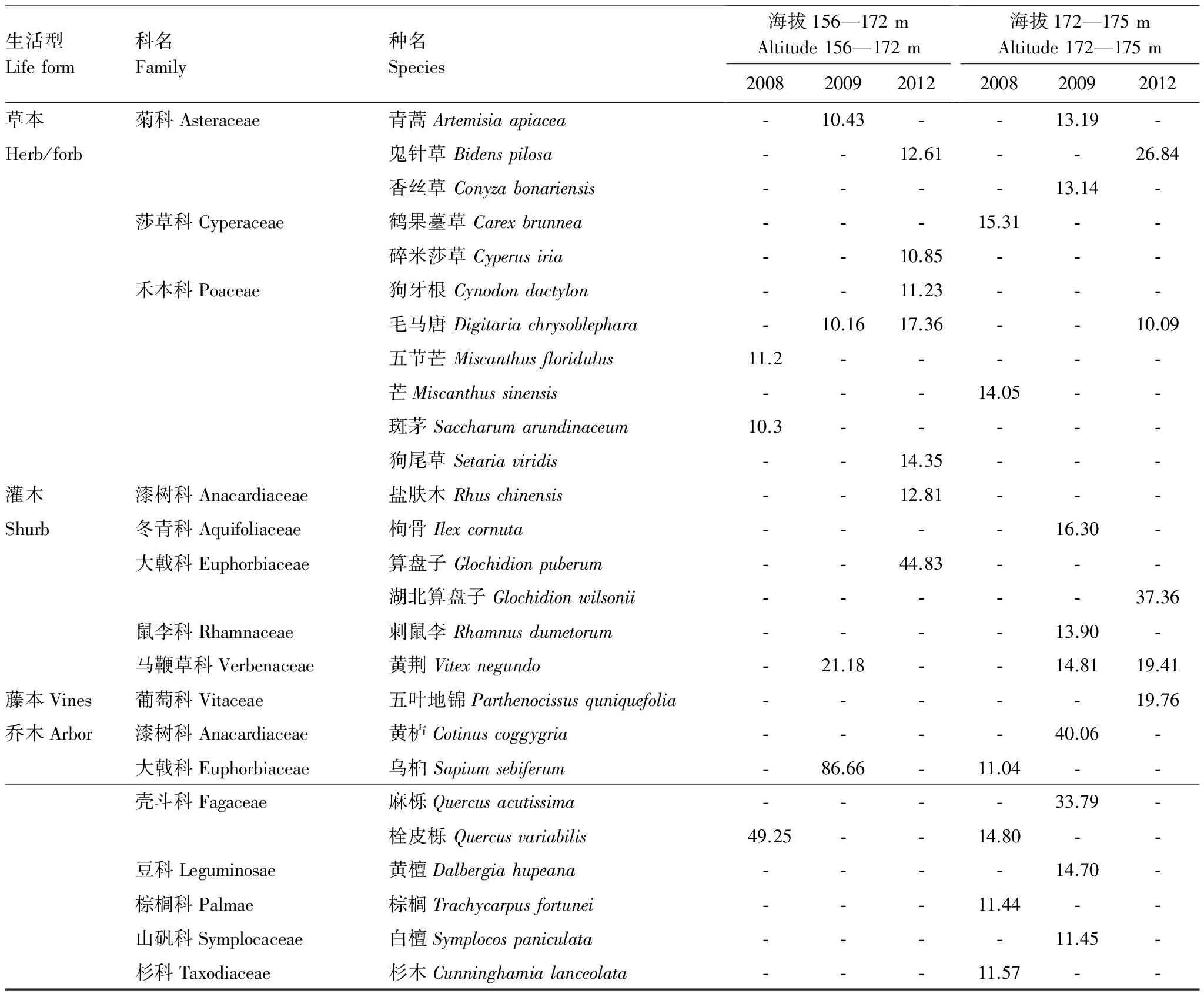
表3 2008—2012年三峡水库消落带优势植物的重要值Table 3 Important values of dominant plants in Hydro-fluctuation Belt of the Three Gorges Reservoir(Year 2008—2012)
从表3可以看出,在海拔156—172 m区段,经历4 次水库水位涨落后(2012年),草本植物的优势种为菊科(Asteraceae)的鬼针草(Bidenspilosa)、莎草科(Cyperaceae)的碎米莎草(Cyperusiria),禾本科(Poaceae)的狗牙根(Cynodondactylon)、毛马唐(Digitariachrysoblephara)、狗尾草(Setariaviridis)。灌木优势种为漆树科(Anacardiaceae)的盐肤木(Rhuschinensis)、大戟科(Euphorbiaceae)的算盘子(Glochidionpuberum),未见乔木优势种。在海拔172—175 m区段,除鬼针草和毛马唐仍为优势种外,还增加了大戟科的湖北算盘子(Glochidionwilsonii),马鞭草科(Verbenaceae)的黄荆(Vitexnegundo),葡萄科(Vitaceae)的五叶地锦 (Parthenocissusquniquefolia)。与海拔156—172 m区段比较,海拔172—175 m区段灌木和藤本的优势种有所增加。
2.2 消落带植物群落的生活型组成及其变化
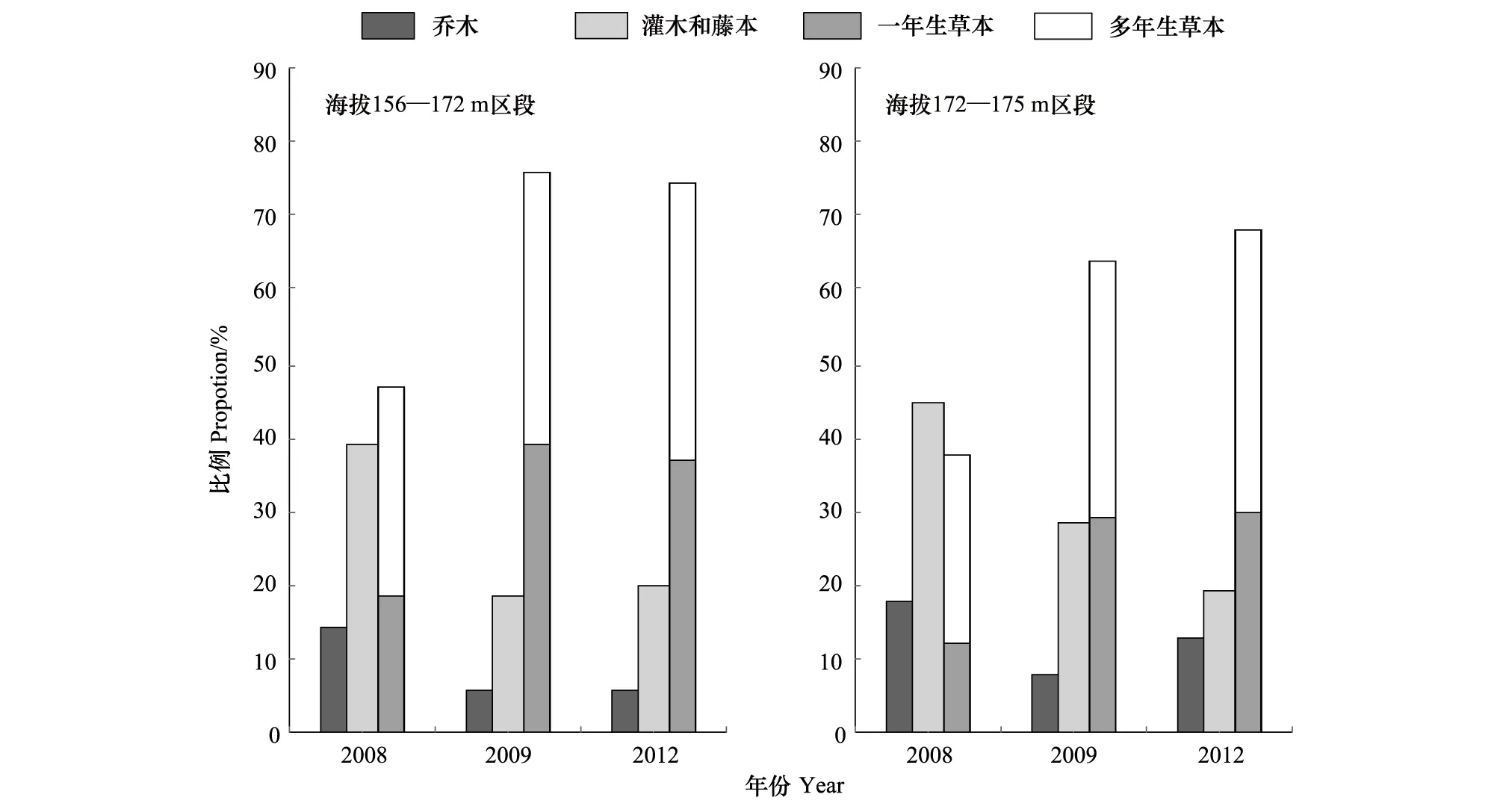
图2 2008—2012年三峡水库消落带植物生活型组成的变化Fig.2 Changes in the composition of plant life form in Hydro-fluctuation Belt of the Three Gorges Reservoir(2008—2012)
2008—2012年,消落带不同海拔区段植物生活型组成的总体变化趋势是:草本植物在群落中所占比例增加,木本植物所占比例减少(图2)。海拔156—172 m区段,经历首次水库水位涨落后,灌木和藤本在群落中所占比例由2008年的38.8%,减少到18.3%,乔木由14.3%减少到6.1%,而一年生草本则由18.4%增加到39.0%,多年生草本由28.6%增加到36.6%。经历4 次水库水位涨落影响后(2012年),灌木和藤本在群落中所占比例减少到20.0%,乔木减少到5.7%,一年生草本增加到37.1%,多年生草本增加到37.1%。海拔172—175 m区段经历2次水库水位涨落后,灌木和藤本在群落中所占比例由2008年的45.1%,减少到19.1%,乔木由17.6%,减少到12.8%,一年生草本由12.1%,增加到29.8%,多年生草本由25.3%增加到38.3%。
2.3 消落带植物群落的物种多样性及其变化
对群落的物种丰富度R、Shannon-Weiner指数H′、Simpson指数Ds、Pielou均匀度指数J的计算结果见图3。
由图3可以看出,在2008—2012年,海拔156—172 m区段草本的R、H′、Ds指数均呈先升后降的变化趋势,均匀度J指数为单调下降;灌木和藤本的H′、Ds、R和J指数均为单调下降。海拔172—175 m区段,草本的H′、Ds指数为单调下降,R指数为先升后降,J指数为先降后升。灌木和藤本的H′、Ds、R指数为单调下降,J指数为单调上升;乔木各项测定指标的变化趋势与灌木和藤本相同。
从总体变化趋势分析,灌木和藤本与乔木的物种多样性指数下降最为明显。海拔156—172 m区段和海拔172—175 m区段的均匀度指数变化趋势相反;草本植物的物种多样性H′、Ds、J指数变化总体上呈下降趋势;经历首次水库水位涨落后,物种丰富度增加,而经历多次水库水位涨落后又有所下降。
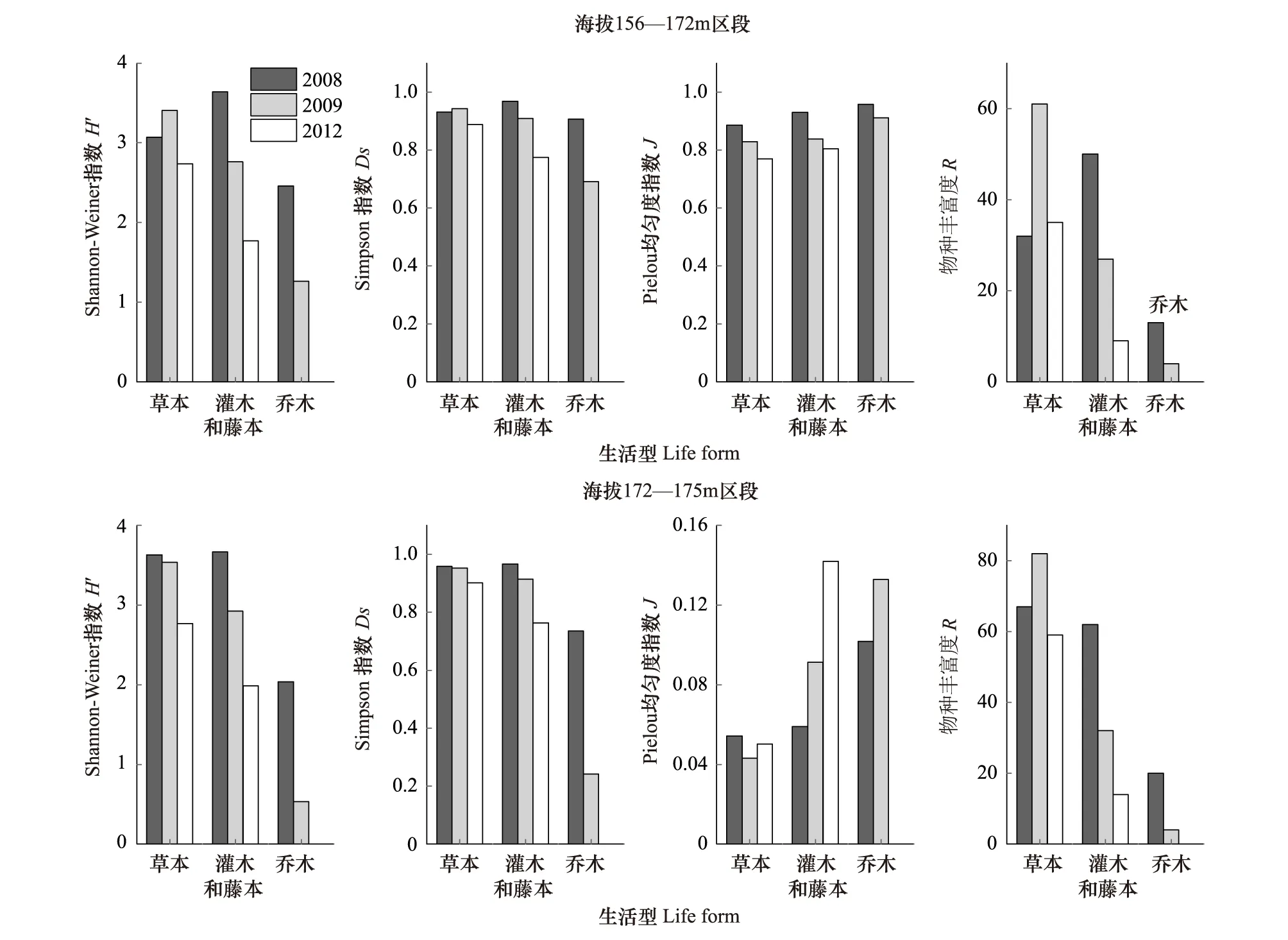
图3 2008—2012年三峡水库消落带植被的物种多样性变化Fig.3 Changes in species diversity of plant community in Hydro-fluctuation Belt of the Three Gorges Reservoir(2008—2012)
3 结论与讨论
3.1 植物的生态适应性与消落带水陆生境变化
消落带原生植物在经历多次水位涨落后大幅度减少。其中,三峡水库水位变动的时机、持续时间、幅度、频率、变化率改变了消落带的物理环境和资源养分的分布[9],以及不同生态适应型的植物对变化环境的响应程度的差异是其发生变化的主要原因[19]。植物生态适应性是在漫长的历史演变过程中通过不断进化而形成的相对固定的一种特性,并在其结构和生理等方面产生了一系列适应性特化[20]。水生植物只所以能够适应缺氧环境,其原因在于其根、茎、叶之间已形成了一整套通气系统。也有一些水生植物能够贮存自身呼吸释放的CO2和光合作用释放的O2,来满足自身光合和呼吸代谢的需要[21]。而大多数陆生植物缺乏这种组织结构和功能,长期深水淹没下植物死亡的主要原因是体内缺氧而导致的窒息死亡。一般认为,植物对渐变的环境,可以通过自身的形态变异或生理过程调节来增加其与环境的适合度[22],但这种“变异”和“调节”需经历一个渐变过程,对消落带原生植物而言,首次经历水库水位涨落完全是一种瞬时、偶发的极端环境变化。在这种突变的环境条件下,植物的“变异”和“调节”能力是极为有限的。因此,植物对首次经历水库水位涨落的反应更为强烈。经历水库水位涨落后消落带上出现的“新”植物,并非是植物与环境长期协同进化意义上的“新种”。 其中,有些植物种可能来自邻近区域,而更多的可能是在经历水库水位涨落前以种子库的形式在消落带存在的植物种。
经过多次水位涨落后成为消落带优势种的生活史对策大多具有r对策种群特征。主要表现是营养生长期相对较短,植物个体较小,初次结实的时间到来较早,能产生大量种子,种子的体形小,可借助风力传播,并迅速占领由于种种原因形成的空旷地[20]等。同时,与Baker提出的理想杂草(the ideal weed)特征非常吻合。 比如能不连续的发芽(自我控制)且种子能在较长时间内保持生命力;幼苗生长迅速;开花前无性生长期短;只要条件允许则能维持连续的种子生产;能在广泛的环境中结实,对气候和土壤变异具有较强的忍耐性(可塑性常常高),对长距离和短距离传播都有特殊的适应方式;如果是多年生植物,则具有可进行无性繁殖的根茎或根状茎[23]。毛马唐、狗尾草、鬼针草、碎米莎草等优势种多属于田间和坡地常见的一年生草本。毛马唐自然生长多见于路旁、田野。喜湿、喜光, 种子耐受水淹的能力较强;狗尾草多生于林边、山坡、路边和荒芜的园地及荒野,对干燥生境有较强的适应能力;鬼针草多生长在路边荒地、山坡及田间,碎米莎草多生于田间、山坡、路旁阴湿处,为秋熟农田的主要杂草。这些植物都具有生活周期短,能在消落带成陆期获得成熟的种子,且种子细小,容易随风和水体四处扩散。鬼针草的瘦果上还具倒刺毛,能沾钩在动物体表进行种子传播。这些植物的种子都能够以休眠的形式越冬,并在适宜温度和水分条件满足时快速萌发完成早期定居过程,进而提高其在植物中的竞争力[24-25];而当萌发条件不具备时,还能够推迟萌发来分散其在不可预测环境中的风险性[26-27],增加幼苗在更适宜的环境条件下出现和生长的机会[28]。狗牙根为多年生草本,具有能够克隆繁殖的匍匐状地下根茎。据野外调查,经历水深2—7 m,淹水100d左右和伏旱季节后,狗牙根的种群密度和生物量不但没有降低,反而比对照还有所提高;在经历水深22—27 m,淹水250d左右的海拔区段,其种群的增殖能力还能维持在较高水平[29]。盐肤木和算盘子为在消落带适应性较强的灌木树种。这两种植物自然分布在山坡、沟谷杂木疏林和灌丛中。盐肤木的萌芽更新能力较强。种皮坚硬,表面具蜡质和油质,在生产上用未经处理的种子播种一般难以发芽,需要进行层积催芽处理。湖北算盘子,多生于山坡灌丛中,耐干旱瘠薄,果期6—9 月,与消落带成陆期一致。目前对这两种木本植物的适应机制还缺乏深入研究,初步分析认为,除与其耐干旱瘠薄,能够在消落带成陆期结实以及特殊的种子结构有关外,长时间淹水对于消除其种子萌发障碍可能还具有一定的促进作用。
3.2 物种多样性和植物生活型组成的时空变化分析
消落带海拔156—172 m区段的植物群落经历了4 次水库水位涨落,而海拔172—175 m区段仅经历了2次水库水位涨落;海拔156—172 m区段每年持续水淹时间为105—243 d,淹水深度在3—19 m,而海拔172—175 m区段每年持续水淹时间约30d左右,淹水深度在0—3 m。虽然不同海拔区段的植物受到水库水位涨落干扰的强度和频次不同,但物种多样性变化总体呈下降的趋势是一致的。这与王强等在三峡水库蓄水初期研究消落带植物物种多样性空间分布格局的结论相吻合[9]。一般认为,一个较成熟的群落往往具有较高的物种多样性、较高的均匀度和较低的生态优势度。如果植物群落朝正向演替(或正向波动),生态系统的物质和能量可得到进一步累积,生物多样性会相应提高;相反,若植物群落朝逆向演替(或负向波动),生物多样性势必下降[30]。根据本研究结果分析认为,目前,三峡库区消落带植物群落仍处在逆向演替的早期阶段,群落的物种组成和结构仍处在不稳定状态。
不同调查时期、不同海拔区段消落带植物生活型组成总的变化趋势是:乔木、灌木和藤本减少,草本增加。这与前人预测和调查的结果相一致[1, 6]。植物生活型是不同植物对相同生境趋同适应的外在表现[31]。植物生活型对水陆生境变化的响应策略,决定了植物群落的物种组成和替代变化趋势,也是消落带植物群落在水陆生境变化下演替的基础。在三峡水库反复周期性水位涨落影响下,草本植物(包括一年生和多年生植物)生活型取代乔木、灌木和藤本生活型是三峡库区消落带植物生活型组成变化的必然趋势。
3.3 奉节以东巫山和秭归段与其他区域消落带植物群落动态特征的比较
三峡水库奉节以东巫山和秭归段库岸山高坡陡,森林资源丰富,且植被的自然度较高,而中部和西南部地区的库岸地势较平缓,以低山丘陵为主,因开发较早,海拔600m以下大多被开垦为农耕地,自然植被已遭到严重破坏。对消落带植物群落的动态而言,水文条件变化是影响其最主导的因素,而植物区系、地形、土壤、人为干扰等也具有重要影响。然而以往的研究往往规避或未能充分考虑这些因素[10]。王强等于2008—2010年监测位于奉节西部的开县典型消落带植被的结果表明,在坡度15°的水田和旱地撂荒地60m×30m的样地上,经历水库水位涨落影响后,不同调查年份的优势植物有苍耳(Xanthiumsibiricum)、双穗雀稗(Paspalumpaspaloides)狗牙根、狗尾草等[9];王业春等在库区西南部的忠县,选择3 个地质地貌和土地利用历史相似的近自然消落带(土地利用历史为农用耕地),对不同水位高程(海拔160m和170m)植物群落和土壤特征的研究结果表明,不同水位高程的植被物种组成完全相同,且盖度、生物量、生物多样性指数均没有显著差异,只是群落优势种存在差异[10]。与本研究结果相比,奉节以东巫山和秭归段消落带的植物组成和优势种以及群落结构的变化等都与上述研究区域有所不同。另外,奉节以东巫山和秭归段消落带的坡度较大,土壤冲刷严重。土壤是植物生存和分布的基础条件,土壤流失速度快,消落带植物群落变化的速度也必然会相应加快。
3.4 消落带植被动态研究方法分析
获取经历水库水位涨落前消落带植物群落组成和结构的初始资料后,对经历水库水位涨落影响后植物的“消长”动态定位“跟踪”,再根据不同植物多年的“消长”变化趋势,确定用于消落带植被恢复的备选植物,较之“以空间代替时间”和一次性调查得出的结果具有一定的可靠性;同时,通过对不同植物和群落结构变化的长期监测,即可获得消落带植物群落演替的确凿证据。三峡水库运行初期是消落带植物群落种类组成和结构快速变化时期,在此时期,定位监测具有其它方法不可替代的重要意义。
固定样地监测的不足在于很难反映较大地理空间尺度上消落带植物群落的宏观变化趋势,而且受人力、物力的限制,很难做到大范围的定位观测。如何将固定样地监测与遥感数据的宏观监测相结合揭示较大地理空间尺度上消落带植物群落的动态变化规律还有待深入研究。
致谢:野外调查得到秭归县和巫山县林业局的大力支持,刘正宇研究员、刘涛老师,周火明副教授、程瑞梅研究员、金江群、巴哈尔古丽、康义、王晓荣、王祥福等硕士研究生参加了部分年度调查,特此致谢。
[1] 胡波, 张平仓, 任红玉, 罗慧敏, 岑奕. 三峡库区消落带植被生态学特征分析. 长江科学院院报, 2010, 27(11):81- 85.
[2] 熊俊, 袁喜, 梅朋森, 张丽萍, 许文年, 黄应平. 三峡库区消落带环境治理和生态恢复的研究现状与进展. 三峡大学学报(自然科学版), 2011, 33(2):23- 28.
[3] 郭泉水, 康义, 洪明, 金江群, 朱妮妮, 聂必红, 王佐庆. 三峡库区消落带陆生植被对首次水陆生境变化的响应. 林业科学, 2013, 49(5):1- 9.
[4] 黄世能, 王伯荪. 热带次生林群落动态研究:回顾与展望. 世界林业研究, 2000, 13(6):7- 13.
[5] 熊平生, 谢世友, 谢金宁. 初探三峡水库湿地面临的问题及其对策. 国土与自然资源研究, 2004, (4):62- 63.
[6] 王勇, 刘义飞, 刘松柏, 黄宏文. 三峡库区消涨带植被重建. 植物学通报, 2005, 22(5):513- 522.
[7] 白宝伟, 王海洋, 李先源, 冯义龙, 智丽. 三峡库区淹没区与自然消落区现存植被的比较. 西南农业大学学报(自然科学版), 2005, 27(5):684- 691.
[8] 卢志军, 李连发, 黄汉东, 陶敏, 张全发, 江明喜. 三峡水库蓄水对消涨带植被的初步影响. 武汉植物学研究, 2010, 28(3):303- 314.
[9] 王强, 袁兴中, 刘红, 张跃伟, 陈忠礼, 李波. 三峡水库初期蓄水对消落带植被及物种多样性的影响. 自然资源学报, 2011, 26(10):1680- 1693.
[10] 王业春, 雷波, 张晟. 三峡库区消落带不同水位高程植被和土壤特征差异. 湖泊科学, 2012, 24(2):206- 212.
[11] 肖文发, 周志翔, 黄志霖, 王鹏程, 吴昌广. 长江三峡库区森林景观格局与景观恢复研究. 北京:科学出版社, 2012.
[12] Whittaker R J, Willis K J, Field R. Scale and species richness:towards a general hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4):453- 470.
[13] Wills K J, Whittaker R J. Species diversity-scale matters. Science, 2002, 295(5558):1245- 1248.
[14] 黄忠良, 彭少麟, 易俗. 影响季风常绿阔叶林幼苗定居的主要因素. 热带亚热带植物学报, 2001, 9(2):123- 128.
[15] 中国长江三峡集团公司水情信息网, 宜昌. http://www.ctg.com.cn/inc/sqsk.php.
[16] 中国植被编辑委员会. 中国植被. 北京:科学出版社, 1980.
[17] 贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征. 植物生态学报, 1998, 22(4):304- 311.
[18] 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6):533- 548.
[19] 丛静, 尹华群, 卢慧, 宿秀江, 杨敬元, 李迪强, 张于光. 神农架保护区典型植被的物种多样性和环境解释. 林业科学, 2013, 49(5):30- 35.
[20] 李俊清. 森林生态学. 北京:高等教育出版社, 2010.
[21] Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science, 2002, 7(9):405- 410.
[22] 常杰, 葛滢. 生态学. 杭州:浙江大学出版社, 2001.
[23] Baker H G, Stebbins G L. The Genetics of Colonizing Species. New York:Academic Press, 1965.
[24] Grime JP, Mason G, Curtis AV, Rodman J, Band S R. A comparative study of germination characteristics in a local Flora. Journal of Ecology, 1981, 69(3):1017- 1059.
[25] Bohumil M. Germination requirements of invasive and non-invasiveAtriplexspecies:a comparative study. Flora-Morphology, Distribution, Functional Ecology of Plants, 2003, 198(1):45- 54.
[26] Venable D L, Brown J S. The selective interactions of dispersal, dormancy and seed size as adaptations for reducing risk in variable environments. The American Naturalist, 1988, 131(3):360- 384.
[27] Philippi T. Bet-hedging germination of desert annuals:variation among populations and maternal effects inLepidiumlasiocarpum. The American Naturalist, 1993, 142(3):488- 507.
[28] Kevin J R, Andrew R D. Seed aging, delayed germination and reduced competitive ability inBromustectorum. Plant Ecology, 2001, 155(2):237- 243.
[29] 洪明, 郭泉水, 聂必红, 康义, 裴顺祥, 金江群, 王祥福. 三峡库区消落带狗牙根种群对水陆生境变化的响应. 应用生态学报, 2011, 22(11):2829- 2835.
[30] 李瑞. 植被动态研究进展及展望. 中国水土保持科学, 2012, 10(2):115- 120.
[31] 蒋有绪, 郭泉水, 马娟. 中国森林群落分类及其群落学特征. 北京:科学出版社, 1998.
Plant community dynamics in the Hydro-fluctuation Belt of the Three Gorges Reservoir at the Zigui and Wushan Section, East of Fengjie County, China
ZHU Nini1,GUO Quanshui1,*,QIN Aili1,PEI Shunxiang2,MA Fanqiang1,ZHU Li1,3, JIAN Zunji1
1ResearchInstituteofForestEcology,EnvironmentandProtection,ChineseAcademyofForestryKeyLaboratoryofForestEcologyandEnvironmentofStateForestryAdministration,Beijing100091,China2ExperimentalCentreofForestryinNorthChina,ChineseAcademyofForestry,Beijing102300,China3CollegeofAgriculture,HenanUniversityofScienceandTechnology,Luoyang471003,China
The plant community growing in the hydro-fluctuation belt is an important component of reservoir management, playing a significant role in the maintenance of riparian ecosystems and ecological functioning. Obtaining timely and accurate data on plant community dynamics could provide theoretical and practical information for vegetation restoration and reconstruction in the Three Gorges Reservoir. This study collected data over 5 years (2008—2012) for the plant community from four permanent monitoring plots at different elevations in Zigui and Wushan section of the Three Gorges Reservoir (East of Fengjie County). The dynamic characteristics, plant community composition, dominant plants, plant life forms, and species diversity were studied. The results indicated that compared to 2008, the elevation from 156 to 172 m was subject to water level fluctuation on one occasion by 2009 and four occasions by 2012. The number of native plants declined from 147 species in 2008 (belonging to 55 families) to 33 species (belonging to 18 families) by 2009; the rate of reduction for families and species was 67.3%and 77.6%, respectively. The number of native plants declined to 39 species (14 families) by 2012, with the rate of reduction for families and species being 74.5%and 73.5%, respectively. However, 49 (by 2009) and 23 (by 2012) “new” species appeared during the study period, raising the total number of species by 59.8%and 32.9%, respectively, in certain sampled plots. The elevation from 172 to 175 m was subject to water level fluctuations on two occasions by 2012. The number of native plants declined from 91 species (40families) to 20species (13 families). The rate of reduction was 67.5%and 78.0%, respectively. However, 21 “new” species accounted for about 44.7%of all sampled plant species. Based on the comparison of several survey results on the number of “disappeared” and “appeared” plant species, the results indicated that the most sensitive response of the plant community was to the first water level fluctuation. Subsequently, even though the plant community was subject to several water level fluctuations, the rate of change declined. The “growth” dynamics of ecologically adapted plants and the composition of dominant species differed at different altitudes. At an altitude of 156—172 m, the dominant herbaceous species wereBidenspilosa(Asteraceae),Cynodondactylon(Poaceae),Digitariachrysoblephara(Poaceae),Setariaviridis(Poaceae), andCyperusiria(Cyperaceae). The dominant shrub species wereRhuschinensis(Anacardiaceae) andGlochidionpuberum(Euphorbiaceae). At an altitude of 172—175 m,Glochidionwilsonii(Euphorbiaceae),Vitexnegundo(verbenaceae) andParthenocissusquniquefolia(Vitaceae) became dominant shrub species, except forBidenspilosa, andDigitariachrysoblephara. Annual and perennial herbs were the dominant plant life-form in the hydro-fluctuation belt. Species diversity decreased as the number of water level fluctuations increased. Periodic fluctuation in the water level of the Three Gorges Reservoir led to flooding-drying habitat alterations. The adaptability of different ecological plants to changing habitats was the main driving factor of plant community change.
plant community dynamics; plant life form; species diversity; The Three Gorges Reservoir; water-level fluctuation belt
林业十二五科技支撑计划项目(2011BAD38BO4)
2014- 04- 24; < class="emphasis_bold">网络出版日期:
日期:2015- 05- 19
10.5846/stxb201404240818
*通讯作者Corresponding author.E-mail:guoqs@caf.ac.cn
朱妮妮,郭泉水,秦爱丽,裴顺祥,马凡强,朱 莉,简尊吉.三峡水库奉节以东秭归和巫山段消落带植物群落动态特征.生态学报,2015,35(23):7852- 7867.
Zhu N N,Guo Q S,Qin A L,Pei S X,Ma F Q,Zhu L, Jian Z J.Plant community dynamics in the Hydro-fluctuation Belt of the Three Gorges Reservoir at the Zigui and Wushan Section, East of Fengjie County, China.Acta Ecologica Sinica,2015,35(23):7852- 7867.
猜你喜欢
云南画报(2021年11期)2022-01-18 03:15:34
中国水利(2020年14期)2020-08-02 16:19:30
铁道通信信号(2018年8期)2018-11-10 05:15:42
铁道通信信号(2018年8期)2018-11-10 05:15:38
成才之路(2017年21期)2017-07-27 20:52:40
安徽农学通报(2017年9期)2017-05-19 23:03:05
现代农业科技(2016年22期)2017-03-24 12:21:57
铁道通信信号(2016年4期)2016-06-01 12:10:19
学周刊·下旬刊(2015年10期)2015-07-22 12:36:17
湖泊科学(2014年4期)2014-02-27 10:30:32
