
冀北辽河源不同林龄油松天然次生林土壤微生物生物量及酶活性1)
2015-02-08 03:02高晶韩海荣康峰峰立天宇宋小帅赵伟红于晓文赵金龙
东北林业大学学报 2015年9期
高晶 韩海荣 康峰峰 立天宇 宋小帅 赵伟红 于晓文 赵金龙
(北京林业大学,北京,100083)
冀北辽河源不同林龄油松天然次生林土壤微生物生物量及酶活性1)
高晶 韩海荣 康峰峰 立天宇 宋小帅 赵伟红 于晓文 赵金龙
(北京林业大学,北京,100083)
以冀北辽河源地区不同林龄油松天然次生林为研究对象,研究其土壤微生物生物量碳、氮质量分数,土壤脲酶、过氧化氢酶、多酚氧化酶以及β-葡萄糖苷酶活性随油松林龄的时间变化趋势。结果表明:在生长季期间,土壤微生物生物量碳、氮质量分数,脲酶、过氧化氢酶和β-葡萄糖苷酶活性从5—8月份逐渐增大,然后逐渐减小,但是多酚氧化酶活性则相反,而且酶活性的衰减程度延迟于微生物生物量碳、氮质量分数的;随着油松天然次生林林龄的增加,土壤微生物量碳、氮质量分数,脲酶、过氧化氢酶和β-葡萄糖苷酶活性逐渐增加,而多酚氧化酶活性则减小。相关性分析表明,土壤微生物生物量碳、氮质量分数与脲酶、过氧化氢酶和β-葡萄糖苷酶活性呈极显著正相关,而与多酚氧化酶活性呈负相关。土壤温度和含水率与各指标之间呈极显著正相关性。
油松天然次生林;土壤微生物生物量;酶活性
土壤微生物是土壤中具有生命力的重要组成部分,参与土壤的形成与发育,并且在物质转化和能量传递等过程中发挥着重要作用[1]。其中,土壤微生物生物量是指土壤中个体体积小于5 000 μm3的微生物数量,是负责参与调控有机质和能量转化的重要媒介,因其周转速度快、灵敏性高,用于反映土壤微生物活性的变化程度[2];另外,土壤酶是由动物、植物根系、微生物以及残体的分泌或分解合成,并参与催化土壤中所有生物化学过程的重要生物催化剂,是联接微生物多样性、植被种类以及土壤理化性质等生态因子的重要纽带[3]。所以,土壤微生物生物量和酶活性是表征土壤质量特征及其动态变化的重要生物学指标,应重视并加强土壤微生物和酶活性在森林生态系统中的动态研究。
油松(Pinustabulaeformis)属于松科松属,是暖温带湿润半湿润气候区的地带性植被,广泛分布于我国北方地区,跨度为北纬31°13′~43°33′,东经103°20′~124°45′,总共占地22.810万hm2。因其具有适应性强、根系发达、抵御不利环境强的特点,已成为华北地区山地植被的主要建群种[4]。不同林龄油松的土壤理化性质以及凋落物各养分含量存在不同程度的差异,比如土壤有机质、全氮、全磷质量分数,以及凋落物碳氮比(C/N)、纤维素与氮的比值等,都会影响土壤中微生物的数量和多样性,继而改变土壤微生物生物量和酶活性的大小[5]。目前,我国对于油松天然次生林的研究主要集中在不同林龄的土壤理化性质、生物量碳储量和土壤酶活性的研究[6-8]。而有关土壤微生物生物量和酶活性对不同林龄油松天然次生林响应的动态研究鲜有报道,尤其是探讨土壤微生物生物量与酶活性之间的耦合关系。因此,本研究拟通过对冀北辽河源地区不同林龄油松天然次生林与碳、氮循环相关的脲酶、过氧化氢酶、多酚氧化酶和β-葡萄糖苷酶,以及微生物生物量碳氮的测定,分析各指标的时间动态和随林龄的变化特征,探讨土壤酶活性与微生物生物量碳氮之间的相关关系,以期更深入了解油松天然次生林土壤的生态过程和功能。
1 研究区概况
研究地点位于河北省平泉县辽河源国家自然保护区内(东经118°22′39″~118°37′21″,北纬41°01′30″~41°21′15″),地处内蒙古高原和冀北山地的过渡地带。该区域处于暖温带向寒温带过渡地带,半湿润半干旱大陆性季风型山地气候,全年平均气温7.3 ℃,无霜期120~130 d,冬季积雪5个月。年平均降水量550 mm,主要集中在7—9月份,年平均蒸发量1 800 mm。该区域土壤以山地棕壤和淋溶褐土为主。土层较厚,一般为50~100 cm[9]。
2 材料与方法
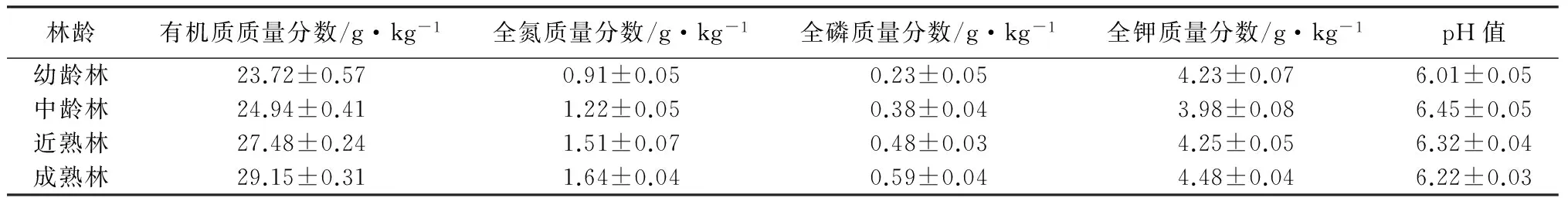
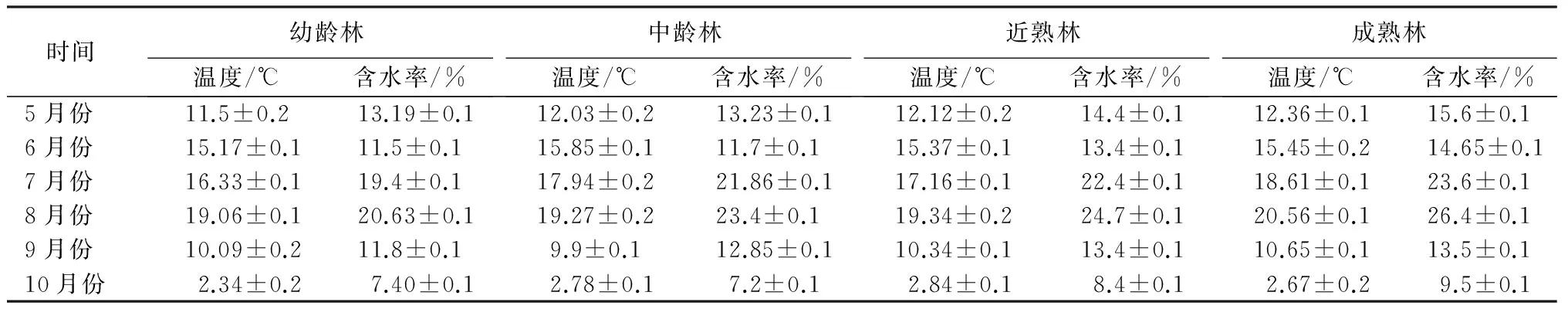
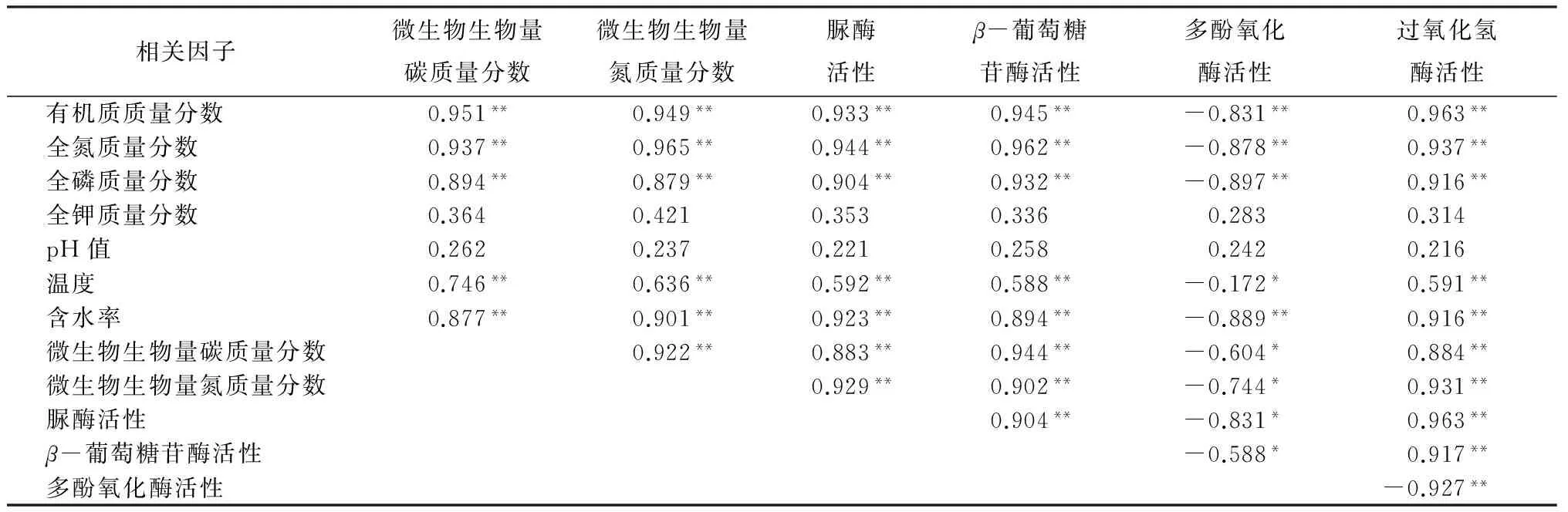
样地选择:于2012年7—9月份,在研究区域内,针对4个不同林龄(Y)的油松天然次生林(幼龄林Y≤30 a、中龄林30 a 表1 油松天然次生林标准地概况 样品采集:分别于2013年5—10月份的每个月20号,在每个样方内,使用土钻按“S”形随机采集5个0~10 cm深度的土样,并且将3个相同林龄重复样地的土样均匀混合,然后过2 mm孔径的土筛去除植物残根和石砾,装入自封袋内并保存于冰箱内4 ℃,用于测量土壤微生物量碳、氮质量分数,土壤脲酶、多酚氧化酶、β-葡萄糖苷酶、过氧化氢酶活性;另取其中一部分土样自然风干,研磨过1 mm孔径土筛,用于测量土壤理化性质。 土壤样品分析:使用LI-8100配套的土壤温度传感器测定0~10 cm深度的土壤温度;采用烘干法测定土壤含水率。土壤有机质质量分数采用重铬酸钾氧化—外加热法测量,土壤全氮质量分数采用浓硫酸—高氯酸消煮—凯式定氮仪进行测量,土壤全磷采用钼锑抗比色法测量,土壤全钾采用火焰光度计法测量,土壤pH值采用酸度计测定[10]。 土壤微生物量碳(MBC)质量分数的测定方法采用氯仿熏蒸浸提法。称取3份10 g新鲜土样到50 mL烧杯中,然后置于装有60 mL三氯甲烷的真空干燥器中,用真空抽气泵反复抽取真空,直至三氯甲烷沸腾1 min,最后放置于黑暗条件下25 ℃培养24 h。同时,再称取3份10 g新鲜土样,不经过三氯甲烷熏蒸作为对照。所有土样浸润于40 mL 0.5 mol·L-1的K2SO4溶液,并在振荡器上震荡30 min(300 r·min-1)后过滤,滤液中的土壤有机碳质量分数的测定采用高温外热重铬酸钾氧化法。土壤微生物量碳(MBC)质量分数的计算公式:wM,B,C=EC/KE,C,式中:wM,B,C表示土壤微生物量碳质量分数;EC表示熏蒸与未熏蒸土样有机碳质量分数的差值;KE,C为转换系数,取值0.45[11-12]。 土壤微生物量氮(MBN)质量分数的测定方法采用氯仿熏蒸浸提法,其中熏蒸浸提步骤同土壤微生物量碳质量分数的。滤液中全氮的测定采用浓硫酸—高氯酸消煮—凯式定氮法。土壤微生物量氮质量分数(wM,B,N)的计算公式:wM,B,N=EN/KE,N,式中:EN表示熏蒸与未熏蒸土样全氮质量分数的差值;KE,N为转换系数,取值0.45[11-12]。 脲酶活性采用苯酚钠—次氯酸钠比色法,多酚氧化酶活性采用邻苯三酚比色法,β-葡萄糖苷酶活性采用比色法,过氧化氢酶活性采用高锰酸钾滴定法[1]。 数据处理:采用Excel对数据进行处理,采用SPSS 19.0软件对数据进行统计分析。用单因素方差分析(one-way ANOVA)和最小差异显著法(LSD,α=0.05),对不同林龄油松天然次生林土壤微生物量碳、氮质量分数和4种土壤酶活性的差异显著性进行检验。采用Pearson法对各个指标之间的相关关系进行分析,利用Sigmaplot 10.0绘图软件绘图。 3.1 不同林龄油松天然次生林土壤微生物生物量碳、氮质量分数的时间动态 由表2可见,不同林龄油松天然次生林土壤微生物生物量碳质量分数的变化具有显著的季节动态。其中,幼林龄土壤微生物生物量碳质量分数从5月份的(63.31±5.29)mg·kg-1增加到8月份的(215.04±12.85)mg·kg-1,随后逐渐下降到10月份的(38.64±8.46)mg·kg-1。相似的,中林龄、近熟林和成熟林的土壤微生物生物量碳质量分数的季节变化均呈现先升高后降低的趋势,都在8月份达到最大值,在10月份降到最小值,其中,土壤微生物生物量碳质量分数在中林龄的变化幅度为(198.78±6.49)mg·kg-1,在近熟林的变化幅度为(244.98±5.44)mg·kg-1,在成熟林的变化幅度为(295.34±4.61)mg·kg-1。在整个生长季中,土壤微生物生物量碳质量分数均随林龄的增加而增加,经过多重比较分析,在7、8、9月份,4个林龄的微生物生物量碳质量分数差异显著;除了幼龄林在6月份与中龄林差异不显著外,在5、6、10月份,幼龄林显著低于其他3个林分的。 由表2可见,在整个生长季中,不同林龄油松天然次生林土壤微生物生物量氮质量分数的变化趋势也是从5月份开始增加到8月份的最大值,随后降低到10月份的最小值。其中,幼龄林微生物生物量氮质量分数的变化幅度为(15.85±0.82)mg·kg-1,中龄林微生物生物量氮质量分数的变化幅度为(18.55±0.52)mg·kg-1,近熟林微生物生物量氮质量分数的变化幅度为(22.11±0.86)mg·kg-1,成熟林微生物生物量氮质量分数的变化幅度为(23.82±1.18)mg·kg-1。而且,经过多重比较分析发现,不同林龄的土壤微生物生物量氮质量分数均随林龄的增加而显著增加。 表2 不同林龄油松天然次生林土壤微生物量碳、氮质量分数,土壤酶活性的季节变化 3.2 不同林龄油松天然次生林土壤酶活性的时间动态 本研究的结果显示,不同林龄油松天然次生林中,土壤脲酶、β-葡萄糖苷酶、多酚氧化酶和过氧化氢酶活性均表现出较明显的季节变化(表2),土壤脲酶、β-葡萄糖苷酶和过氧化氢酶活性均从5月份开始升高,在8月份达到最大值,随后逐渐降低。但是,与土壤微生物量碳、氮质量分数不同,酶活性的最小值发生在5月份,而且从8—10月份的降低幅度小于5—8月份的升高幅度。另外,脲酶活性的整体变化幅度较小,而β-葡萄糖苷酶和过氧化氢酶活性的整体变化幅度较大。其中,脲酶活性5—8月份的变化幅度为(291.06±5.59)U·kg-1·h-1(表2);β-葡萄糖苷酶活性5—8月份的变化幅度为(4.00±0.11)U·kg-1·h-1(表2);过氧化氢酶活性从5月份到8月份的变化幅度为(18.31±0.58)U·g-1·h-1(表2)。相反的,多酚氧化酶活性则从5月份开始降低,在8月份达到最小值,然后逐渐升高,5—8月份的变化幅度为(305.67±5.84)U·g-1·h-1(表2)。在整个生长季期间,脲酶、β-葡萄糖苷酶和过氧化氢酶活性均随林龄的增加而增加。其中,经过多重比较分析发现,近熟林和成熟林的脲酶活性在整个生长季的差异都不显著,而与幼龄林和中龄林之间的差异则显著;各林龄的β-葡萄糖苷酶和过氧化氢酶活性在7、8、9、10月份的差异显著,幼龄林和中龄林的β-葡萄糖苷酶活性在5月份与近熟林的无差异,5—6月份的过氧化氢酶活性与近熟林的无差异。 3.3 土壤微生物生物量碳、氮质量分数,酶活性与土壤理化性质、温度和含水率的相关性分析 各林龄油松天然次生林的土壤化学性质见表3,其中,土壤有机质、全氮和全磷质量分数随林龄的升高而增加,而土壤全钾质量分数随林龄的增加呈现先降低后升高的趋势;各林龄不同月份的土壤温度、含水率情况见表4,土壤温度从5月份开始逐渐升高,在8月份达到最大值,然后迅速下降,在10月份达到最小值,而且随林龄的增加呈现递增趋势。土壤含水率从5月份开始先减小后增加,在8月份达到最大值,然后迅速降低,在10月份达到最小值,而且随林龄的增加而增加。通过相关性分析可以看出(表5),土壤有机质、全氮、全磷质量分数,含水率,温度分别与土壤微生物生物量碳、氮质量分数,脲酶、β-葡萄糖苷酶、过氧化氢酶活性呈极显著正相关(p<0.01),而分别与多酚氧化酶呈极显著负相关(p<0.01);土壤全钾和pH值与土壤微生物生物量碳、氮质量分数,脲酶、β-葡萄糖苷酶、多酚氧化酶和过氧化氢酶活性的相关性不显著。另外,多酚氧化酶活性分别与β-葡萄糖苷酶、过氧化氢酶、脲酶活性,微生物生物量碳、氮质量分数均呈显著负相关(p<0.05)。土壤微生物生物量碳、氮质量分数分别与脲酶、β-葡萄糖苷酶、过氧化氢酶活性呈极显著正相关(p<0.01)。 表3 不同林龄油松天然次生林土壤化学性质 表4 供试土壤温度及含水率 表5 土壤微生物生物量碳、氮质量分数,酶活性与土壤理化性质、温度、含水率的Pearson相关系数 土壤微生物生物量碳质量分数虽然只占土壤有机碳的1%~4%,但却占到微生物干物质的40%~50%,是土壤活性有机碳库的重要组成部分,也是土壤有效养分重要的源与汇。土壤微生物生物量氮质量分数一般占土壤全氮质量分数的2%~7%,是土壤中有机—无机态氮转化的一个关键环节[13]。作为土壤微生物数量的指标,土壤微生物生物量因其周转快、灵敏度高的特点,经常被用作指示土壤总有机质、全氮的状态和积累程度[14]。 本研究表明,土壤微生物生物量表现出明显的季节差异,从5月份开始增加,到8月份达到最大值然后逐渐下降。这是由于随着温度的升高,微生物的繁殖和生长得到促进,凋落物分解加剧,分解后的碳水化合物、养分元素等随降水从地表转移到土壤中,反过来继续供给微生物生长和繁殖,从而固定较多的微生物生物量碳、氮[15];反之,温度降低,微生物活性和数量均受到抑制,导致微生物生物量降低。针对含水率是否对土壤微生物生物量有影响没有明确的结论,谢涛等[16]的研究表明,土壤微生物生物量与土壤含水率无显著相关性。但是也有学者发现,土壤含水率会显著影响土壤微生物生物量[17]。而在本研究中,土壤微生物生物量与含水率有极显著的正相关关系,说明土壤含水率的高低对土壤微生物生物量的大小构成了限制。 另外,随着油松天然次生林林龄的增加,土壤微生物生物量碳、氮质量分数均表现出增加的趋势,即由大到小的顺序为成熟林、近熟林、中龄林、幼龄林。而且土壤微生物生物量碳、氮质量分数与土壤有机质、全氮和全磷质量分数呈极显著正相关,这是由于随着林龄的增加,凋落物逐年积累,根系逐年增大,这些分别通过凋落物的分解、养分的释放和运输,使土壤中有机质等营养物质质量分数增加,从而为土壤微生物新陈代谢和自身合成提供越来越多的碳源、氮源以及能量来源[18]。而且,随着林龄的增加,自疏作用会导致油松天然次生林林冠层的郁闭度降低,使得林内光照更充足,水热条件更有利于土壤微生物生长和繁殖,并且林地地表的灌木、草本等植被多样性增加,使得土壤中微生物多样性得到丰富[19]。综上所述,油松天然次生林林龄对土壤微生物生物量的影响较为敏感,随着油松林龄的增加,土壤积累有机质等养分的能力越来越强。 土壤酶主要来源于土壤微生物、植被根系分泌以及动植物残体分解,而且作为生物催化剂,参与几乎所有与物质循环和能量流动相关的生物化学反应[20]。土壤酶活性的高低能够直接反映土壤生物活性的大小,与土壤微生物生物量一样,都是被用于评价土壤质量变化的敏感生物学指标[21]。 多酚氧化酶属于氧化还原胞外酶,将土壤中芳香族有机化合物氧化成醌来进行碳捕获,进一步转化为腐殖质[21]。在本研究中,多酚氧化酶活性随油松天然次生林林龄的增加而降低,而且与土壤有机质、全氮、全磷质量分数呈极显著负相关,可能是由于地表凋落物现存量增加,土壤有机质质量分数较多,腐殖质层厚度增加,从而抑制多酚氧化酶活性[22-23]。 β-葡萄糖苷酶是将纤维素分解为葡萄糖的限制酶,降低有机物质的分子结构。一般与有机质质量分数呈显著正相关[24]。过氧化氢酶负责氧化还原生化过程中的有害中间产物,反映了与生物化学相关的物质和能量转化的强度[25]。在本研究中,β-葡萄糖苷酶和过氧化氢酶活性均随林龄的增加而增加,而且与土壤有机质、全氮、全磷质量分数呈极显著正相关,表明油松天然次生林一直处于有机质等养分积累阶段,而且养分周转速率加快,土壤质量上升。 脲酶属于酰胺酶,参与有机氮尿素中酰胺键的水解过程,生产矿质态NH3和CO2,其中矿质态铵根离子是植被氮素养分的直接来源,所以通常用来表征土壤中氮素的供应状况。在本研究中,脲酶活性随油松林龄的增加而增加,表明土壤中有机氮的转化速率是逐渐增加的。而且,相关性分析表明,土壤有机质、全氮质量分数与脲酶活性均呈极显著正相关,是由于在碳水有机化合物分解过程中,常伴随含氮化合物的转化[26],这进一步说明有机质质量分数高的土壤有利于提高氮素的转化率[27]。 此外,对这4种土壤酶活性进行相关性分析后发现,脲酶、β-葡萄糖苷酶和过氧化氢酶这3种酶活性两两呈极显著正相关且随林龄的增加而增加,而与多酚氧化酶活性呈极显著负相关,说明这4种酶协同参与了凋落物、根系等有机残体转化为可被植物吸收利用的养分物质的生化过程,而且含碳有机化合物的分解通常伴随着含氮有机化合物的转化[28]。因此,油松林龄的增加对加速养分循环和提高碳氮利用率有重要的生态学意义。土壤微生物生物量碳、氮质量分数可以反映土壤有效养分的可利用状况以及微生物活性的高低[20],而且,相关分析表明,土壤微生物生物量碳、氮质量分数分别与土壤有机质、全氮和全磷质量分数呈极显著正相关关系,都随着油松林龄的增加而增加,表明土壤有机质、全氮和全磷养分质量分数的高低对土壤微生物生物量碳、氮质量分数的大小起到决定性的作用,这是由于土壤有机质等养分为微生物的生长和繁殖提供了物质基础[29]。同样,相关分析表明,土壤有机质、全氮和全磷质量分数与脲酶、β-葡萄糖苷酶、过氧化氢酶均呈极显著正相关,这是由于土壤有机质等养分是酶催生化反应的物质基础,当有机质等养分质量分数增加时,会加速酶促反应将有机化合物转化为可被根系直接吸收利用的养分。因此,有机质等养分与土壤酶活性之间形成了一种正反馈机制[30]。另外,有些水解酶、转化酶等会通过吸附作用,与有机质形成“酶—腐殖质复合物”[31],所以,土壤中的过氧化氢酶、脲酶等酶活性与土壤有机质等养分质量分数呈正相关关系[28]。 综上所述,土壤微生物生物量和酶活性的高低均反映了有机质等养分的积累与转化情况[29]。在生长季期间,所有土壤生物活性指标(除了多酚氧化酶)的变化均呈现“钟型”趋势,即从5月份开始增加,到8月份开始降低。但是,各土壤酶的活性在8月份后的衰减程度延迟于微生物量碳、氮质量分数的,这说明微生物已经完成了生命循环或者进入休眠期,而微生物细胞的溶解为维持酶活性提供了物质基础[32]。 在本研究中,随着林龄的增加,土壤有机质、全磷、全氮质量分数,土壤微生物生物量碳、氮质量分数,脲酶、过氧化氢酶和β-葡萄糖苷酶活性都增加且互相之间有显著正相关性,而与多酚氧化酶活性有负相关性,说明土壤微生物生物量碳、氮质量分数与酶活性在土壤碳氮循环和代谢中的重要作用,从而可表征土壤肥力[28],可作为评价不同林龄土壤质量状况较好的生物学指标[33]。由此可见,在冀北辽河源地区,随着油松天然次生林林龄的增大,其土壤生化强度和微生物群落稳定性更高,土壤生态系统趋于稳定。 [1] 林先贵.土壤微生物研究原理与方法[M].北京:高等教育出版社,2010:2-6. [2] 黎荣彬.土壤微生物生物量碳研究进展[J].广东林业科技,2008,24(6):65-69. [3] Allison S, Vitousek P. Responses of extracellular enzymes to simple and complex nutrient inputs[J]. Soil Biology and Biochemistry,2005,37(5):937-944. [4] 李国雷,刘勇,李瑞生,等.油松人工林土壤质量的演变[J].林业科学,2008,44(9):76-81. [5] Saswati M, Vadakepuram C J. Influence of leaf litter types on microbial functions and nutrient status of soil: ecological suitability of forest trees for afforestation in tropical laterite wastelands[J]. Soil Biology and Biochemistry,2010,42(12):2306-2315. [6] 赵伟红,康峰峰,韩海荣,等.冀北辽河源地区不同林龄油松天然次生林土壤理化特征的研究[J].西北林学院学报,2014,29(3):1-8. [7] 程小琴,韩海荣,康峰峰.山西油松人工林生态系统生物量、碳积累及其分布[J].生态学杂志,2012,31(10):2455-2460. [8] 陈琦,尹粉粉,曹靖,等.秦岭西部不同发育阶段油松和日本落叶松人工林土壤酶活性变化和分布特征[J].生态与农村环境学报,2010,26(5):466-471. [9] 杨玉梅,谷建才,周奇,等.辽河源针叶树种生长与土壤因子的相关分析[J].东北林业大学学报,2012,40(5):16-18. [10] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科学技术出版社,2000:146-190. [11] Vance E D, Brookes P C, Jenkinson D S. An extraction method for measure microbial biomass C[J]. Soil Biology and Biochemistry,1987,19(6):703-707. [12] Pandey C B, Singh G B, Singh S K, et al. Soil nitrogen and microbial biomass carbon dynamics in native forest and derived agricultural land and uses in a humid tropical climate of India[J]. Plant Soil,2010,333(1):453-467. [13] 刘纯,刘延坤,金光泽.小兴安岭6种森林类型土壤微生物量的季节变化特征[J].生态学报,2014,34(2):451-459. [14] 杨凯,朱教君,张金鑫,等.不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化[J].生态学报,2009,29(10):5500-5507. [15] Pascoal C, Cá ssio F. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river[J]. Applied Andenvironmental Microbiology,2004,70(9):5266-5273. [16] 谢涛,郑阿宝,王国兵,等.苏北不同林龄杨树林土壤活性碳的季节变化[J].生态学杂志,2012,31(5):1171-1178. [17] 刘爽,王传宽.五种温带森林土壤微生物生物量碳氮的时空格局[J].生态学报,2010,30(12):3135-3143. [18] 邸雪颖,耿莹莹,孙龙,等.兴安落叶松林土壤微生物生物量季节动态及影响因素[J].东北林业大学学报,2012,40(5):1-4. [19] 阎德仁,王晶莹,杨茂仁.落叶松人工林土壤衰退趋势[J].生态学杂志,1997,16(2):62-66. [20] 鲁顺保,周小奇,芮亦超,等.森林类型对土壤有机质、微生物生物量及酶活性的影响[J].应用生态学报,2011,22(1):2567-2573. [21] 胡嵩,张颖,史荣久,等.长白山原始红松林次生演替过程中土壤微生物生物量和酶活性的变化[J].应用生态学报,2013,24(2):366-372. [22] 张超,刘国彬,薛萐,等.黄土丘陵区不同林龄人工刺槐林土壤酶演变特征[J].林业科学,2010,46(12):23-29. [23] 葛晓改,肖文发,曾立雄,等.三峡库区不同林龄马尾松土壤养分与酶活性的关系[J].应用生态学报,2012,23(2):445-451. [24] Sinsabaugh R L, Gallo M E, Lauber C W, et al. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition[J]. Biogeochemistry,2005,75(2):201-215. [25] 陈彩虹,叶道碧.4种人工林土壤酶活性与养分的相关性研究[J].中南林业科技大学学报,2010,30(6):64-68. [26] 关荫松.土壤酶及其研究方法[M].北京:农业出版社,1986:294-296. [27] 焦晓光,隋跃宇,张兴义.土壤有机质含量与土壤脲酶活性关系的研究[J].农业系统科学与综合研究,2008,24(4):493-495. [28] 范艳春,王鹏程,肖文发,等.三峡库区2类典型森林5种土壤酶季节动态及其与养分的关系[J].华中农业大学学报,2014,33(4):39-44. [29] 王莹,王彦梅,陈龙池.湖南会同地区森林植被转变对土壤微生物生物量碳和酶活性的影响[J].生态学杂志,2010,29(5):905-909. [30] 严金龙.湿地稻田土壤酶分布与活性及生态功能指示[D].南京:南京农业大学,2011:22-23. [31] 黄巧云,李学垣.粘粒矿物,有机质对酶活性的影响[J].土壤学进展,1995,23(4):12-18. [32] Bhattacharya P, Pal R, Chakraborty A, et al. Microbial biomass and activity in a laterite soil amended with municipal solid waste compost[J]. Journal of Agronomy and Crop Science,2001,187:207-211. [33] 赵汝东,樊剑波,何园球,等.不同母岩区马尾松人工林土壤酶活性及微生物学性质研究[J].生态环境学报,2011,20(2):253-258. Soil Microbial Biomass and Enzymes Activity ofPinustabulaeformisForest with Different Ages in Liaohe River of Northern Hebei// Gao Jing, Han Hairong, Kang Fengfeng, Li Tianyu, Song Xiaoshuai, Zhao Weihong, Yu Xiaowen, Zhao Jinlong (Beijing Forestry University, Beijing 100083, P. R. China)//Journal of Northeast Forestry University,2015,43(9):78-83. We studied the temporal variation tendency of soil microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), activity of urease, catalase, polyphenol oxidase, andβ-glucosidase ofPinustabulaeformiswith different ages in Liao River source region. MBC, MBN, urease, catalase andβ-glucosidase in soil of all forest age groups increased gradually from May to August, and then decreased until October. However, soil polyphenol oxidase exhibited the opposite trend. The attenuation degree of enzyme activity delayed than that of MBC and MBN. With the increasing age ofP.tabulaeformisforest, MBC, MBN, urease, catalase andβ-glucosidase increased, while the polyphenol oxidase decreased. By correlation analysis, MBC and MBN had a statistically significantly positive correlation with the activity of urease, catalase andβ-glucosidase. However, there was negative correlation between MBC/MBN and polyphenol oxidase. Soil temperature and water content had a significantly positive correlation with all indices. MBC, MBN and enzymes activity of soil were the biological indices to evaluate soil quality of different age groups inP.tabulaeformisforest. Pinustabulaeformisforest; Soil microbial biomass; Enzymes activity 高晶,女,1985年3月生,北京林业大学林学院,博士研究生。E-mail:gaojingcrystal@126.com。 康峰峰,北京林业大学林学院,讲师。 E-mail:phoonkong@163.com。 2015年3月17日。 Q148;S714.3;S718.8 1)林业公益性行业科研专项(201204101)。 责任编辑:任 俐。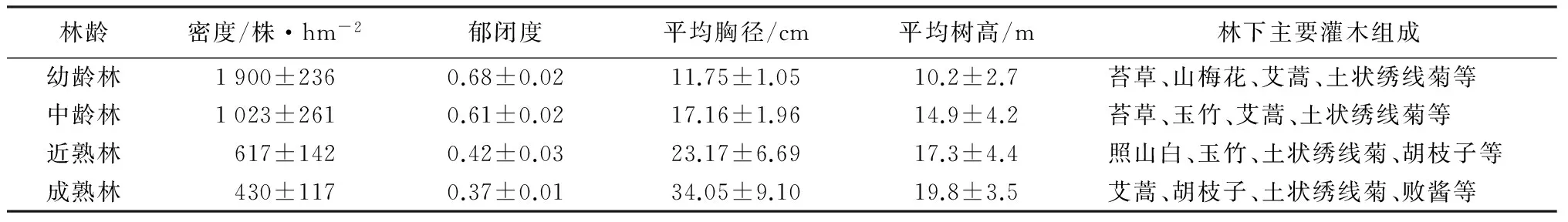
3 结果与分析

4 结论与讨论
猜你喜欢
防护林科技(2022年5期)2022-08-22
防护林科技(2020年10期)2020-12-22
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
四川林业科技(2020年1期)2020-08-31
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
现代农业研究(2017年11期)2018-01-12
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
现代园艺(2017年20期)2017-02-03
