
三维空间深度位置上基于空间的返回抑制*
2015-02-05 22:18王爱君李毕琴张明
心理学报 2015年7期
王爱君李毕琴张明
(1东北师范大学心理学院,长春130024)(2苏州大学心理学系,苏州215123)(3江西师范大学心理学院,南昌330027)
1 引言
人类生活在一个三维的真实世界里,每天接触的物体也是具有不同深度平面的三维客体。虽然这种三维客体被投射到视网膜上的时候为二维的图像,但大脑却能够从二维的视网膜图像重新建构对真实世界的三维表征(Durand et al.,2007;Georgieva,Peeters,Kolster,Todd,&Orban,2009;Harris&Drga,2005;Hayman,Verriotis,Jovalekic,Fenton,&Jeffery,2011;Janssen,Vogels,&Orban,2000;Sereno,Trinath,Augath,&Logothetis,2002;Yamane,Carlson,Bowman,Wang,&Connor,2008)。因此,视觉系统能够有效地将注意定位到不同深度位置上的客体(Previc,1998)。然而,目前关于视觉空间注意的理论模型基本都是基于二维平面(Eriksen&Yeh,1985;LaBerge&Brown,1989;Posner,Davidson,&Snyder,1980),而人类视觉系统最为重要的一个特性就是能够感知深度线索。所以,注意是如何在大脑重构出的深度位置之间进行定向/重定向则是需要关注的问题。
注意在视觉空间中分布规律的研究主要为基于位置(Location-based)注意的空间分布,其中包括聚光灯模型(Spotlight model)(Posner,1984)、变焦镜模型(Zoom-lens model)(Eriksen&St James,1986)和空间梯度模型(Spatial gradient model)(Downing&Pinker,1985;LaBerge&Borwn,1989;Mangun&Hillyard,1988)。基于位置的注意理论认为,注意分布是以特定的空间位置或区域为基础,被试根据任务要求,注意焦点可以像聚光灯一样移动,注意范围可以像变焦镜头一样进行缩放以及注意在焦点处集中程度最高,并向四周逐渐扩散。Posner将注意类比为聚光灯的视觉定向理论(Visual orienting theory)系统地阐述了二维平面上不同注意定向方式对视觉加工的影响,并首次在二维平面内采用外源性位置提示的空间启动范式发现了当线索与目标之间的时间间隔(Stimulus Onset Asynchrony,SOA)小于300 ms时,被试对呈现在线索化位置上的目标反应较快(Posner,Rafal,Choate,&Vaughan,1985),即早期的易化效应。然而关于三维空间注意定向的研究还存在着争议。一些研究发现,被试对于出现在与线索化位置具有不同深度平面位置上的目标反应更慢,并且当无效线索引起注意在空间内转移时,由近处向远处转移注意时出现的注意损耗比从远处向近处转移的注意损耗大(Andersen&Kramer,1993;Downing&Pinker,1985;Gawryszewski,Riggio,Rizzolatti,&Umiltá,1987;Maringelli,McCarthy,Steed,Slater,&Umiltà,2001)。因此提出了注意在深度位置上的定向是基于观察者中心的理论模型。该理论认为,刺激距离观察者越近,加工的就越充分,并以观察者为中心向远处逐渐扩散。然而,另一些研究则发现,被试对出现在线索化位置上的目标和与线索化位置不在同一深度的非线索化位置上目标的反应时并无差异。表明了注意并不能有效地在深度位置上进行定向,即出现“深度盲”现象(Ghirardelli&Folk,1996;Iavecchia&Folk,1994)。近期的一个研究将Posner空间预提示范式应用到三维空间,将目标刺激分别置于三维空间的近处空间和远处空间,考察了三维空间中深度位置上注意定向/重定向的早期易化效应。行为结果发现,对于突然意外地出现在距离我们较近位置上目标刺激的注意重定向要快于远离我们的目标刺激的重定向。功能性磁共振成像结果发现,除了经典的右侧颞顶联合区(right temporoparietal junction,TPJ)存在激活外,还存在另外两个大脑网络涉及了三维空间注意重定向的不同方面:双侧前运动皮层(bilateral premotor cortex)涉及了空间注意在不同深度平面内的重定向(人类的双侧前额叶皮层被认为涉及提取和加工来自视差形成的三维客体的结构)(Georgieva et al.,2009);包括后扣带皮层(posterior cingular cortex,PCC)、眶前额叶皮层(orbital prefrontal cortex,OPC)以及左侧角回(left angular gyrus,AG)的“默认网络(default-mode network)”促进了对于突然出现的客体进行注意重定向(Chen,Weidner,Vossel,Weiss&Fink,2012)。
注意定向不仅包括早期易化,还涉及晚期抑制。Posner等(1985)的研究发现,提示条件下,注意在空间位置上的定向对相继目标刺激的检测具有不同的时序特征。当SOA大于300 ms时,被试对呈现在线索化位置上的目标反应较慢,这种晚期的抑制效应被称之为返回抑制(Inhibition of return,IOR)(Posner et al.,1985)。基于空间的返回抑制是注意定向对位置加工的抑制效应,有利于提高个体进行空间搜索的效率(Klein,2000)。已有关于返回抑制的研究大多集中在二维平面(Fuentes,Vivas,&Humphreys,1999;Klein,2000;张阳,张明,2006;Zhang,Zhou,&Zhang,2012;也见综述,张阳,彭春花,孙洋,张明,2013),且现有研究对三维空间深度位置上是否存在基于空间的返回抑制现象仍然有一定的争议。Theeuwes和Pratt(2003)的研究认为,深度位置上不存在基于空间的返回抑制,即“深度盲”。该研究发现,当注意在同一平面内定向/重定向时才会出现基于空间的返回抑制(比如,近处空间/远处空间的左侧和右侧位置间),而注意在不同的深度位置间转移并不存在基于空间的返回抑制(比如,近处空间的左侧与远处空间的左侧间),但研究却发现,同一平面线索化位置产生的抑制效应可以扩散到与其不同深度平面同一位置(比如,线索化近处空间的左侧位置,如果该位置得到了抑制,那么远处空间的左侧位置也会产生抑制效应)。Bourke,Partridge和Pollux(2006)的研究对Theeuwes和Pratt(2003)的结论产生了质疑,他们认为未出现基于空间的返回抑制的原因是由于后部平面内的“8”字部分的被前面的“8”字所遮挡,而非Theeuwes和Pratt(2003)所认为返回抑制的“深度盲”现象是与上丘(Superior colliculus)相关(一些研究认为上丘并不负责编码视觉空间的深度,具有“深度盲”的特性(Klier,Wang,&Crawford,2001))。因此,为了避免产生遮挡,他们增加了呈现在左右视野中不同深度平面两个“8”字的水平距离。结果发现深度位置上出现了少量基于空间的返回抑制。但是,由于该研究中增大了深度位置上两个刺激的水平距离(X轴方向),因此,研究结果中出现的少量返回抑制可能混淆了注意在水平平面(X轴方向)定向/重定向带来的影响。除此之外,上述关于三维空间中基于空间返回抑制的研究均存在一定的问题,即由于返回抑制研究范式中央线索化(二次线索化)的作用,导致了注意在单纯深度位置上的转移并不是沿着直线方向(如无效线索条件下,线索化近处空间左侧位置后,中央注视点二次线索化,而之后的目标刺激则会出现在远处空间左侧位置)。而以往二维平面返回抑制的研究中,注意大多都沿着直线方向转移。研究表明,相对于直线转移,注意沿着非直线转移到“新异”刺激(非线索化下的目标)的距离增大,导致了注意重定向到“新异”刺激的速度变慢,进而返回抑制量减小。Bennett和Pratt(2001)认为相对于直线转移,注意沿着非直线转移的过程会削弱返回抑制的量。Pratt,O'donnell和Morgan(2000)认为,返回抑制范式中的二次线索化位置如果是固定的中央注视点位置也会削弱返回抑制的量。此外,注意动量假说认为,实验中每个试次均存在三种注意运动方式:从中央注视点到线索、从线索返回到中央注视点以及从注视点到目标。当注意第三次运动的路径和方向与第二次相同时,注意定向/重定向到目标的速度最快(Pratt,Adam,&McAuliffe,1998;Pratt&Abrams,1999),即当目标出现在与线索化位置正相反直线位置上的非线索化位置上反应最快。因此,三维空间中如果注意沿着直线方式转移是否会出现基于空间的返回抑制是本研究需要探讨的问题。
此外,关于三维空间返回抑制的研究中,沈模卫、高在峰、张光强、水仁德、乔歆新和李伟健等(2007)针对二维平面注意诱导范式下眼跳因素与注意因素未能有效分离,从而降低了空间返回抑制机制研究内部效度的角度出发,以随机点立体图构建三维倾斜平面(含倾斜方向和倾斜程度信息的平面),将多个倾斜平面置于视野同一位置可不必执行眼跳(或者眼跳计划)以排除眼跳因素干扰。实验范式为先给予被试2 s的注意引导,然后呈现测试平面,通过将引导倾斜平面与测试倾斜平面的差异作为返回抑制是否出现的指标。结果发现,注意引导平面与测试平面方向一致条件下的反应显著慢于方向不一致条件下的反应,表明了视差线索构造的三维倾斜平面中存在与注意有关的返回抑制效应。而本研究主要采用虚拟现实技术构建一个三维场景,将Posner经典的二维平面线索化范式应用到三维空间,考察注意在三维场景的近处空间和远处空间进行定向/重定向而产生的基于空间的返回抑制。之所以采用虚拟现实三维场景是因为,虚拟现实提供了一种仿真实三维环境的途径,并且能够精确地记录和操纵行为反应(Gamberini,Seraglia,&Priftis,2008;Standen&Brown,2005)。研究分为两个实验,实验1采用线索化范式,操纵目标深度和线索有效性两个因素,被试的注意转移方式为沿着不同深度位置上的对角线方向(如无效线索条件下,线索化近处空间左侧位置后,中央注视点二次线索化,而之后的目标刺激则只会出现在远处空间右侧位置)进行直线转移。但是这样的设置可能会存在注意在水平平面(X轴)上转移带来的混淆,而不是纯深度上(Z轴)的注意转移。因此,实验2则通过将注意转移方式设置为沿着不同深度位置上单纯的深度位置(Z轴)间,以进一步明确三维空间深度位置上是否存在基于空间的返回抑制。三维空间的返回抑制可分为近处空间的返回抑制和远处空间的返回抑制,将不同空间条件下的返回抑制量进行比较可以探讨注意在不同深度空间主导下进行定向/重定向的异同。
2 实验1
2.1 方法
2.1.1 被试
被试为东北师范大学25名学生(男生11人,女生14人),年龄介于18~25岁,所有被试视力正常或者矫正视力正常,之前均未参加过类似实验,实验后被试获得一定的报酬。
2.1.2 实验仪器与材料
所有的刺激均呈现在由Nvidia GeForce FX 5200显卡驱动的ASUS 3D 27英寸显示器上,屏幕的分辨率为1200×1060,刷新率为120 Hz。双眼视差由Nvidia 3D视觉系统调节实现,与显示器同步的Nvidia 3D快门式眼镜将图像分开地呈现在两只眼睛中,每只眼镜呈现的图像分辨率为800×600,刷新率为60Hz。实验中所有3D刺激呈现在黑色的背景上由Presentation软件编程(Neurobehavioral Systems Inc.)实现。实验在隔音、微暗的房间内单独进行。被试通过一具有良好时间精度的游戏手柄(莱仕达光影PXN-8633PRO)对实验任务做出反应。
每个试次中三维虚拟场景的背景为黑色,场景中包括近处空间2个白色方块占位符,以及远处空间2个白色方块占位符,因此在虚拟的三维空间中建构了4个空间位置(如图1)。被试坐在距离显示器屏幕150 cm位置的椅子上,头部固定在下巴托上,使得其眼睛视线与显示屏最中央的位置保持水平。刺激之间不同的距离是由双眼视差所产生,相对于中央注视点(0视差),近处空间和远处空间的视差为±52.40弧分。被试自我报告当其注视屏幕最中间的注视点时,他们能够清晰地感知到近处空间的客体凸出在显示器屏幕外,而远处空间的客体凹在显示器屏幕里面。近处空间、远处空间中左侧和右侧占位符的水平距离在视角上均匹配(16.85°)(例如,近处空间与远处空间中两个占位符空间位置的视网膜距离是相同的)。由于近处空间和远处空间中刺激的大小相同,就导致了这两个空间的刺激在视网膜上的大小存在差异:近处空间占位符的视角为1.98°,远处空间占位符的视角为1.52°。如果不同空间刺激的大小在视网膜上匹配的话,被试在主观上就会感觉远处空间的刺激就会异常的大,这不符合我们日常生活中体验到事物的生态效度(Andersen,1990;Gawryszewski et al.,1987)。为了使得不同空间的刺激之间相互不产生遮挡,每个空间的刺激都稍微地倾斜一定的角度,近处空间与远处空间刺激的位置与垂直方向倾斜了2.36°。其中一半被试完成近处空间2个占位符的空间位置较低的试次,而另外一半被试完成近处空间2个占位符的空间位置较高的试次。实验中目标刺激为蓝色的小球,外周线索化的提示线索为其中一个占位符快速地消失(set off),中央线索化的提示线索为中央注视点快速地消失。
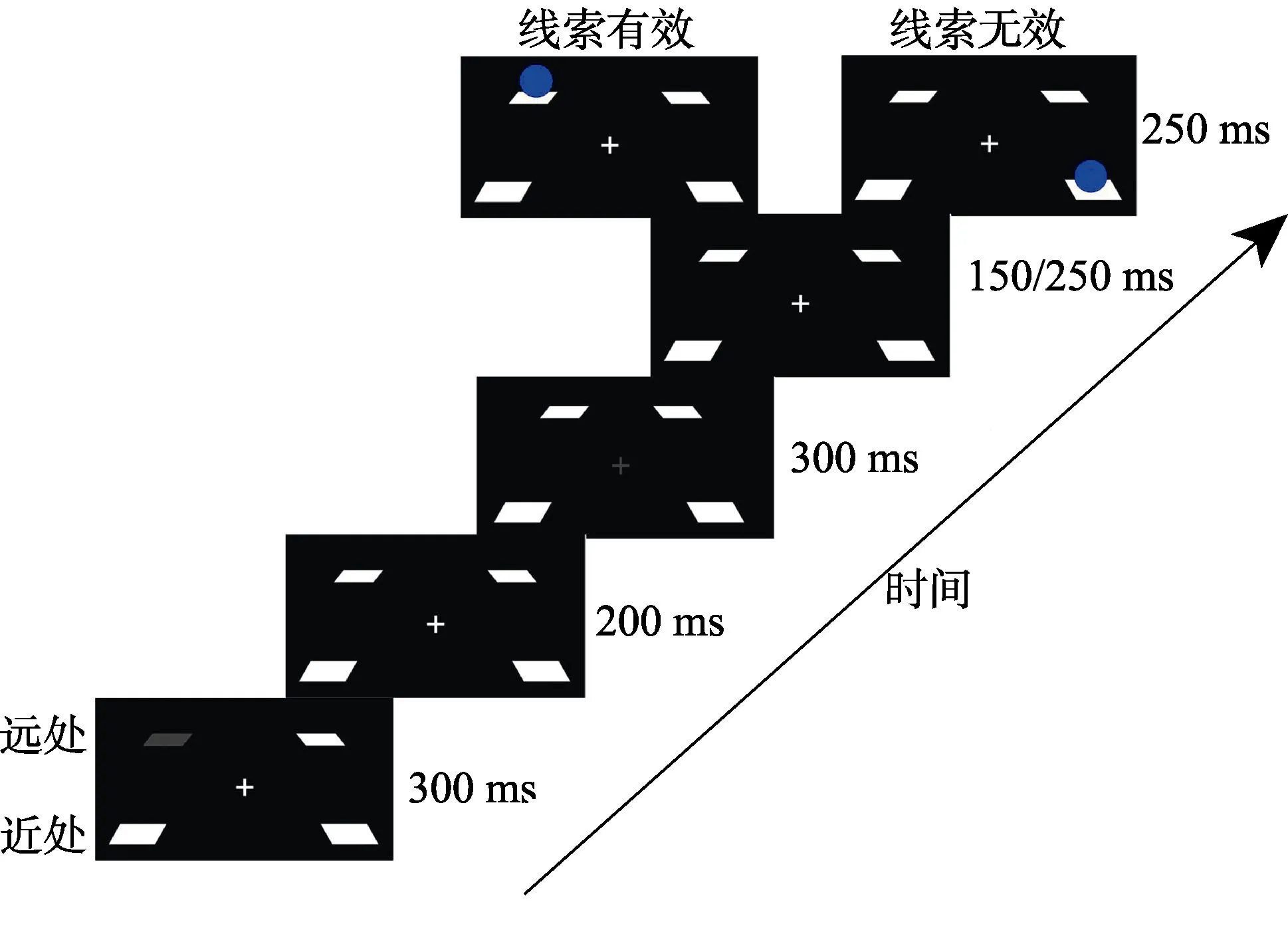
图1 实验1流程图
2.1.3 实验设计与实验程序
实验为2×2的被试内设计,自变量1为目标的深度,包括两个水平,目标出现在近处空间位置和目标出现在远处空间位置,自变量2为线索有效性,包括两个水平,线索有效和线索无效。因变量为被试进行探测任务的反应时和准确率。
实验流程如图1所示:在每个试次的开始,近处空间或远处空间的一个占位符消失,此为外周线索化,时间为300 ms;200 ms的时间间隔(ISI)后,中央注视点消失,此为二次线索化(中央线索化),时间为300 ms;150 ms或者250 ms的时间间隔(ISI)(两个时间在实验中随机出现,目的是防止被试形成暂时的注意定势)后目标刺激(蓝色小球)出现,时间为250 ms;实验任务为要求被试看到蓝色小球出现就既快又准地按键进行探测反应。实验中线索有效性为50%。整个实验过程中都要求被试盯住最中央注视点(0视差),因此,不论目标深度还是线索有效性,实验中4个条件的双眼视差都是恒定的。
实验中每个被试需要完成224个试次,每种条件下48个试次,其中还包括32个捕捉试次。正式实验之前要求被试先完成16个试次的练习,以熟悉实验任务和适应三维空间立体感。正式实验分成4组,每组56个试次。每组结束后休息60s,以缓解视觉疲劳。
2.2 结果与分析
实验为简单的探测任务,总体的漏报率仅为0.37%,所以结果分析中只分析被试的反应时数据(捕捉试次不进入数据分析)。剔除正负三个标准差之外的数据和漏报的数据,总剔除率为1.6%。各个条件下的平均反应时见图2。
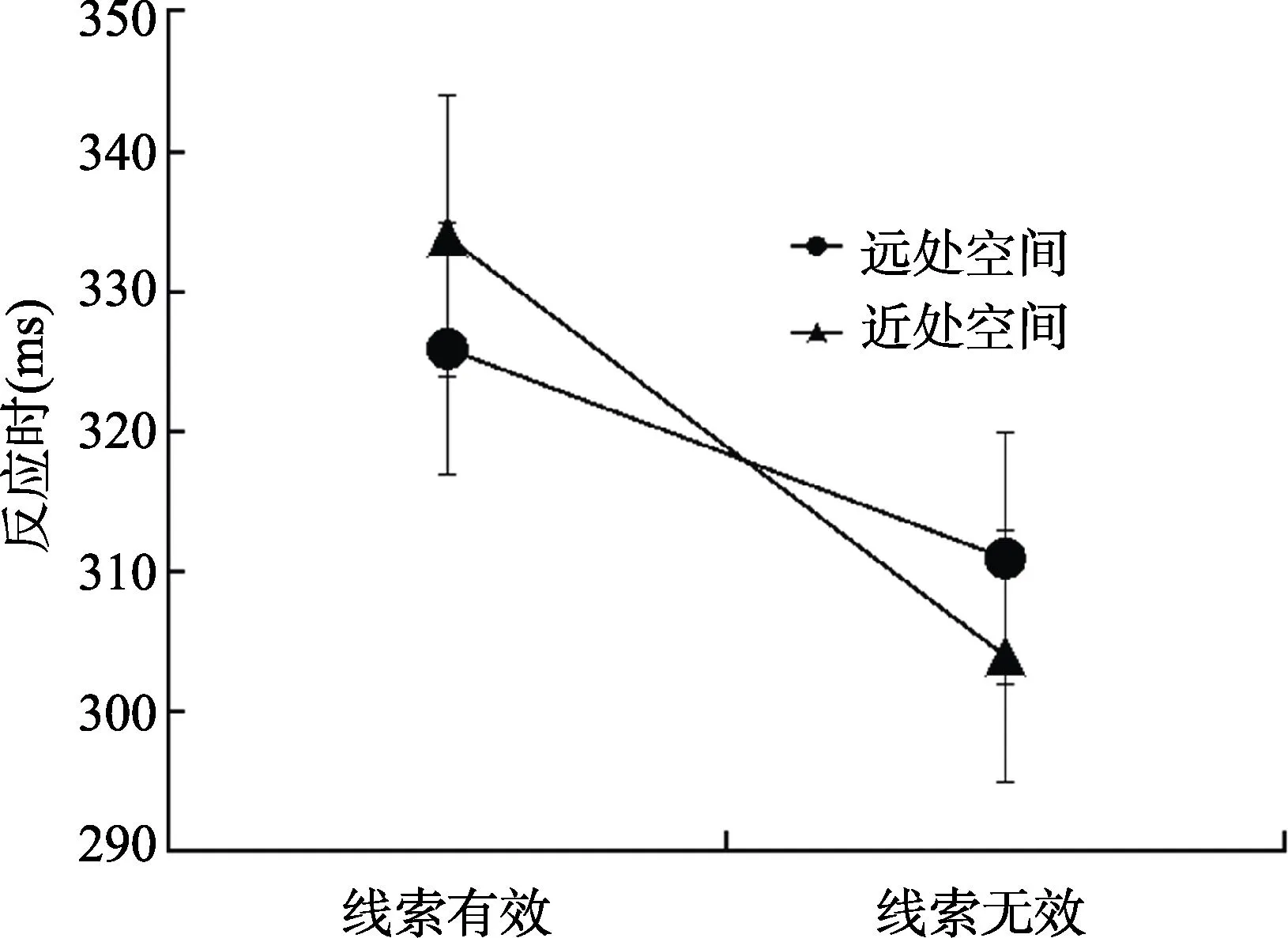
图2 实验1各个条件下对目标刺激的平均反应时间和标准误(ms)
对被试的反应时数据进行2(目标深度:近处空间vs.远处空间)×2(线索有效性:有效vs.无效)的重复测量方差分析。结果显示,目标深度的主效应不显著,F
(1,24)=0.13,p
>0.05;线索有效性的主效应显著,F
(1,24)=66.75,p
<0.001,η=0.74,有效线索条件下的反应(330ms)显著慢于无效线索条件下的反应(307ms),表明了被试在非线索化条件下的反应均快于线索化条件下的反应,即出现了典型的返回抑制。此外,目标深度和线索有效性的交互作用显著,F
(1,24)=15.59,p
=0.001,η=0.35。进一步将目标出现在远处空间和近处空间时线索有效和线索无效条件下的反应时分别进行配对样本t
检验。结果显示,当目标出现在远处空间时,线索有效与线索无效之间的反应时差异显著,线索有效条件下的反应(326ms)显著慢于线索无效条件下的反应(311ms),t
(24)=5.68,p
<0.001,d
=1.64,即出现了典型的返回抑制效应(效应量=15ms)。当目标出现在近处空间时,线索有效与线索无效之间的反应时同样差异显著,线索有效条件下的反应(334 ms)显著慢于线索无效条件下的反应(304ms),t
(24)=7.89,p
<0.001,d
=2.27,即同样出现了典型的返回抑制效应(效应量=30ms)。从图2中可以看出在线索有效条件下,目标出现在近处空间的反应(334ms)慢于目标出现在远处空间的反应(326ms),t
(24)=2.29,p
<0.05,d
=0.66。而在线索无效条件下,目标出现在近处空间的反应(304ms)快于目标出现在远处空间的反应(311ms),t
(24)=3.76,p
=0.001,d
=1.08。上述结果说明了,三维空间深度位置上的注意定向与重新定向中存在返回抑制效应,但目标出现在近处空间时的返回抑制大于目标出现在远处空间。注意从近处空间离开再重新定向到近处空间时的反应最慢,而注意从远处空间突然重新定向到近处空间时的反应最快。由于本实验中注意在三维空间深度位置上进行定向/重新定向的过程中,存在水平平面(X轴)注意定向/重新定向的混淆,即当被试在深度位置上进行注意定向/重新定向时既跨了水平平面(X轴)也跨了深度平面(Z轴)。因此不能将实验结果笼统地归结于注意是沿着单纯的深度位置上进行定向/重新定向而产生基于空间的返回抑制。为了更清楚地确定注意在三维空间深度位置上进行定向/重新定向的过程中是否能够产生基于空间的返回抑制,本研究进行了实验2。实验2保留实验1中的4个空间位置,除此之外,在中间平面(注视点所在平面,双眼视差为0)注视点的两侧各增加一个空间位置(二次线索化的位置)。因此,当被试在此虚拟三维空间的空间位置上进行注意定向/重新定向时,仅仅是在单纯的深度位置之间进行。
3 实验2
3.1 方法
3.1.1 被试
被试为东北师范大学26名学生(男生12人,女生14人),年龄介于18~25岁,所有被试视力正常或者矫正视力正常,之前均未参加过类似实验,实验后被试获得一定的报酬。
3.1.2 实验仪器与材料
实验仪器、实验材料的设置与实验1相同。不同的是本实验在虚拟三维空间中间平面(双眼视差为0)的注视点两侧(与近处空间的占位符具有相同的水平位置)增加了两个白色的占位符。因此构成了三维空间中的6个空间位置(图3)。
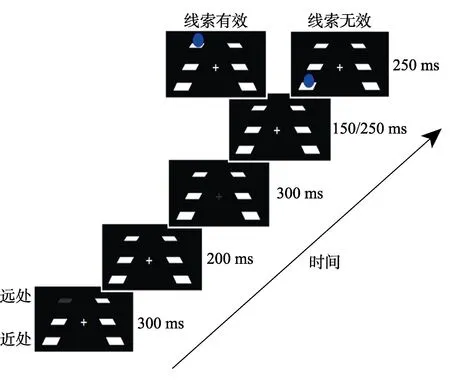
图3 实验2流程图
3.1.3 实验设计与实验程序
实验范式和实验设计与实验1相同。
实验流程:每个试次的开始,近处空间或远处空间的一个占位符消失,此为外周线索化,时间为300 ms;200 ms的时间间隔(ISI)后,中间平面相应于提示线索一侧的占位符消失,此为二次线索化(中央线索化),时间为300 ms;150 ms或者250 ms的时间间隔(ISI)(两个时间在实验中随机出现,目的是防止被试形成暂时的注意定势)后目标刺激(蓝色小球)出现,时间为250 ms;实验任务为要求被试看到蓝色小球出现就既快又准地按键进行探测反应。实验中线索有效性为50%。整个实验过程中都要求被试盯住最中央注视点(0视差),因此,不论目标深度还是线索有效性,实验中4个条件的双眼视差都是恒定的(如图3)。
同样地,实验中每个被试需要完成224个试次,每种实验条件48个试次。其中包括32个捕捉试次。正式实验之前要求被试先完成16个试次的练习,以熟悉实验任务和适应三维空间立体感。正式实验分成4组,每组56个试次。每组结束后休息60 s,以缓解视觉疲劳。
3.2 结果与分析
实验为简单的探测任务,总体的漏报率仅为0.5%,所以结果分析中只分析被试的反应时数据。剔除正负三个标准差之外的数据和漏报的数据,总剔除率为1.5%。各个条件下的平均反应时见图4。

图4 实验2各个条件下对目标刺激的平均反应时间和标准误(ms)
对被试的反应时数据进行2(目标深度:近处空间vs.远处空间)×2(线索有效性:有效vs.无效)的重复测量方差分析。结果显示,目标深度的主效应不显著,F
(1,25)=0.28,p
>0.05;线索有效性的主效应显著,F
(1,25)=58.12,p
<0.001,η=0.70,有效线索条件下的反应(392ms)显著慢于无效线索条件下的反应(371ms),表明了被试在非线索化条件下的反应均快于线索化条件,即出现了典型的返回抑制。此外,目标深度和线索有效性的交互作用显著,F
(1,25)=9.33,p
=0.005,η=0.27。进一步将目标出现在远处空间和近处空间时线索有效和线索无效条件下的反应时分别进行配对样本t
检验。结果显示,当目标出现在远处空间时,线索有效与线索无效之间的反应时差异显著,线索有效条件下的反应(391 ms)显著慢于线索无效条件下的反应(375 ms),t
(25)=4.36,p
<0.001,d
=1.23,即出现了典型的返回抑制效应(效应量=16 ms)。当目标出现在近处空间时,线索有效与线索无效之间的反应时同样差异显著,线索有效条件下的反应(394 ms)显著慢于线索无效条件下的反应(367ms),t
(25)=8.27,p
<0.001,d
=2.34,即同样出现了典型的返回抑制效应(效应量=27ms)。从图4中可以看出线索有效条件下,目标出现在近处空间的反应(394ms)慢于目标出现在远处空间的反应(391ms),虽然差异不显著,但是存在差异的趋势。而在线索无效条件下,目标出现在近处空间的反应(367ms)快于目标出现在远处空间的反应(375ms),同样地,两者虽然差异不显著,但是存在差异趋势。本实验的结果同实验1的结果一致,说明了在三维空间深度位置上的注意定向与重新定向中存在返回抑制效应,但目标出现在近处空间时的返回抑制大于目标出现在远处空间。注意从近处空间离开再重新定向到近处空间时的反应最慢,而注意从远处空间突然重新定向到近处空间时的反应最快。
由于本实验中的二次线索化的位置也涉及到了线索化左侧视野和右侧视野。为了验证左侧视野和右侧视野是否对返回抑制产生不同的影响,我们将被试的反应时数据进行了2(目标深度:近处空间vs.远处空间)×2(线索有效性:有效vs.无效)×2(视野:左侧vs.右侧)的重复测量方差分析。结果显示,视野的主效应不显著,左侧视野和右侧视野间的反应时不存在显著差异,同时,目标深度和视野的交互作用不显著,线索有效性和视野的交互作用不显著,并且目标深度、线索有效性和视野的交互作用也不显著。所有上述检验的F
值均小于1。因此可以认为,左侧视野和右侧视野对于返回抑制效应并不存在显著影响。将实验1的返回抑制量与实验2的返回抑制量结合起来进行分析,可以考察注意沿着两种不同的直线方式进行转移时产生的返回抑制差异。如果两者之间存在显著差异,则说明了实验1结果中三维空间深度位置间出现的返回抑制混淆了注意沿着同一平面(X轴方向)定向/重定向带来的影响;如果两者之间不存在显著差异,则一方面可以说明实验1的结果并不涉及注意沿着同一平面(X轴方向)定向/重定向带来的影响,另一方面也可进一步证实三维空间中深度位置上确实存在基于空间的返回抑制,即返回抑制并不是“深度盲”。通过文中脚注2的算法分别得到了实验1和实验2目标出现在远处空间和近处空间返回抑制的量。将实验1目标出现在远处空间的返回抑制量和实验2目标出现在远处空间的返回抑制量进行独立样本t
检验,结果发现两者的返回抑制量并不存在显著差异,t
<1。同样地,将实验1目标出现在近处空间的返回抑制量和实验2目标出现在近处空间的返回抑制量进行独立样本t
检验,结果发现两者的返回抑制量也不存在显著差异,t
<1。因此可以认为实验1得到的结果并不涉及注意沿着同一平面(X轴方向)定向带来的影响,也可进一步证实三维空间中深度位置上确实存在基于空间的返回抑制,即返回抑制并不是“深度盲”。4 总讨论
本研究采用虚拟现实技术构建了一个虚拟的三维空间场景,将Posner经典的二维平面的注意定向范式应用到了三维空间中近处空间和远处空间,目的是要探究三维空间深度位置上是否存在基于空间的返回抑制效应。实验1和实验2均通过操纵目标深度和线索有效性,考察了在深度位置间注意沿不同方向进行直线转移时而产生的基于空间的返回抑制。实验结果发现,无论注意沿着何种直线转移方式(实验1和实验2)进行定向/重定向均能产生基于空间的返回抑制,即被试对出现在线索化空间位置的目标反应慢于出现在非线索化空间位置的目标。研究表明,与二维平面类似,注意能够有效地在三维空间深度空间位置上进行定向/重定向而产生基于空间的返回抑制,而非“深度盲”。
从图2和图4中可以看出,实验1和实验2均产生了基于空间的返回抑制。一方面,这可以归结为三维空间提供了较高生态效度的视觉场景,利于在水平平面(X轴)和深度平面(Z轴)诱发返回抑制。与此观点一致的是,一些研究认为返回抑制能够优化视觉搜索的效率(Klein,2000;Klein&MacInnes,1999),并且是一种以空间环境(Environmental coordinate)和视网膜(Retinotopic coordinate)双重坐标定义的运作机制(Mathôt&Theeuwes,2010)。因此,本研究发现的三维空间深度位置上基于空间的返回抑制也同样对优化视觉搜索效率有着重要意义。另一方面,注意在二维平面内的定向/重定向存在着双向的模式,即早期的易化和晚期的抑制(Maruff,Yucel,Danckert,Stuart,Currie,1999;McAuliffe&Pratt,2005;Pratt,Hillis,&Gold,2001),即使这种易化或抑制机制可能受到多种因素的制约,如线索和目标的时空关联(McAuliffe&Pratt,2005)、线索与目标的时间间隔(Maylor,1985;Maylor&Hockey,1987;Posner et al.,1985)、线索的凸显性(Mele,Savazzi,Marzi,&Berlucchi,2008)和线索的物理特征(Pratt et al.,2001)等。同样地,注意在三维空间定向/重定向而产生易化的研究也受到多种条件的制约。由于这些条件的制约导致了一些研究认为注意在深度位置上的定向存在“深度盲”现象(Ghirardelli&Folk,1996;Iavecchia&Folk,1994),而另一些研究则认为注意可以在深度位置上有效的定向/重定向(Downing&Pinker,1985;Gawryszewski et al.,1987;Maringelli et al.,2001)。因此我们认为,在排除一些影响因素的制约下,三维空间注意定向/重定向的过程中也会产生返回抑制。此外,从实验1与实验2的结果来看,两个实验条件下返回抑制的量不存在显著差异,说明了无论注意沿着何种方向的直线转移均能产生基于空间的返回抑制,也表明了空间返回抑制效应敏感于深度线索,而非以往研究得出的三维空间深度位置上不存在基于空间的返回抑制(Theeuwes&Pratt,2003),也非三维空间深度位置上存在少量基于空间返回抑制(Bourke et al.,2006)。
从实验1和实验2的结果可以发现,目标出现在近处空间和目标出现在远处空间的返回抑制量存在差异。当目标出现在近处空间时产生的返回抑制量大于目标出现在远处空间时产生的返回抑制量(图2和图4)。进一步分析发现,造成这样结果的原因是,在线索无效条件下,目标出现在近处空间的反应快于目标出现在远处空间,t
(50)=2.88,p
<0.01,d
=0.83。这与以往的研究发现较为一致,即尽管由近及远和由远到近的路径完全一致,但当目标突然地出现在近处空间时的反应要快于目标突然地出现在远处空间(Chen et al.,2012)。这一结果也符合视觉空间注意的梯度理论和观察中心的空间表征理论,注意梯度理论认为,注意的强度是以注意焦点为中心向四周扩散(Gawryszewski et al.,1987)。加之,Andersen和Kramer(1993)提出了注意的空间分布具有基于观察者为中心的空间表征,该理论认为,注意的分布是基于观察者中心(viewer-centered)的,在深度上体现为,注意资源的分布从距离观察者较近的位置向远处位置逐渐递减,在反应时间上表现为从近处到远处成递增趋势。更重要的是,在我们的日常生活中,所处的环境都是三维空间的,所接触的物体都是具有深度的三维客体。那么,一个重要的生态学观点认为,人类为了更好的生存,当一个具有潜在威胁性的刺激突然逼近我们时,我们能够迅速地将注意定位到这些具有潜在威胁的刺激上。因此,对其的反应较快。相反,距离我们较远的刺激尽管对于日常生活也重要,但是这些刺激不具有紧急性和直接性,从而获得的注意资源也较少,相对应的反应也较慢(Gawryszewski et al.,1987;Graziano&Cooke,2006)。综上所述,目标突然出现在近处空间时可以加大返回抑制的量,而目标出现在远处空间时则减小了返回抑制的量。结合图2和图4总体上来看,实验2的反应时长于实验1的反应时。根据我们的预实验结果发现,当注意沿着单纯的深度位置进行注意定向/重定向时,由于这两个深度位置的客体投射到我们视网膜上的位置距离较近,因此,被试不能有效地在近处空间和远处空间的两个位置上进行注意的定向/重定向,进而导致了相对于实验1的注意转移方式来说,被试在此条件下进行定向/重定向时的任务更难,所以反应较慢。此外,两个实验中都要求被试在整个实验过程中盯住中央注视点做内隐性的注意定向。因此,被试的注意焦点更多时间还是集中在了中央注视点的位置,而中央注视点的位置正好为实验1中注意转移方式下的二次线索化位置。所以,相对于实验2而言,被试沿着单纯深度位置间路径进行直线转移的任务而言,在实验1中被试沿着深度位置的对角线方向进行直线转移的任务会较为容易。
5 结论
注意能够有效地在三维空间深度位置上定向/重定向而产生基于空间的返回抑制;返回抑制现象并不是“深度盲”。
致谢
:感谢华南师范大学心理学院的李悠和美国达特茅斯学院心理与脑科学研究所的程优对本论文提供的建议和帮助。Andersen,G.J.(1990).Focused attention in three-dimensional space.Perception&Psychophysics,47
(2),112–120.Andersen,G.J.,&Kramer,A.F.(1993).Limits of focused attention in three-dimensional space.Perception&Psychophysics,53
(6),658–667.Bennett,P.J.,&Pratt,J.(2001).The spatial distribution of inhibition of return.Psychological Science,12
(1),76–80.Bourke,P.A.,Partridge,H.,&Pollux,P.M.J.(2006).Additive effects of inhibiting attention to objects and locations in three-dimensional displays.Visual Cognition,13
(5),643–654.Chen,Q.,Weidner,R.,Vossel,S.,Weiss,P.H.,&Fink,G.R.(2012).Neural mechanisms of attentional reorienting in three-dimensional space.The Journal of Neuroscience,32
(39),13352–13362.de Gonzaga Gawryszewski,L.,Riggio,L.,Rizzolatti,G.,&Umiltá,C.(1987).Movements of attention in the three spatial dimensions and the meaning of“neutral”cues.Neuropsychologia,25
(1A),19–29.Downing,C.,&Pinker,S.(1985).The spatial structure of visual attention.In:M.I.Posner&O.S.Marin(Eds.),Attention and performance XI
(pp.171–187).Hillsdale,NJ:Erlbaum.Durand,J.B.,Nelissen,K.,Joly,O.,Wardak,C.,Todd,J.T.,Norman,J.F.,...Orban,G.A.(2007).Anterior regions of monkey parietal cortex process visual 3D shape.Neuron,55
(3),493–505.Eriksen,C.W.,&St James,J.D.(1986).Visual attention within and around the field of focal attention:a zoom lens model.Perception&Psychophysics,40
(4),225–240.Eriksen,C.W.,&Yeh,H.H.(1985).Allocation of attention in the visual field.Journal of Experimental Psychology:Human Perception and Performance,11
(5),583–597.Fuentes,L.J.,Vivas,A.B.,&Humphreys,G.W.(1999).Inhibitory tagging of stimulus properties in inhibition of return:Effects on semantic priming and flanker interference.The Quarterly Journal of Experimental Psychology Section A:Human Experimental Psychology,52
(1),149–164.Gamberini,L.,Seraglia,B.,&Priftis,K.(2008).Processing of peripersonal and extrapersonal space using tools:Evidence from visual line bisection in real and virtual environments.Neuropsychologia,46
(5),1298–1304.Georgieva,S.,Peeters,R.,Kolster,H.,Todd,J.T.,&Orban,G.A.(2009).The processing of three-dimensional shape from disparity in the human brain.The Journal of Neuroscience,29
(3),727–742.Ghirardelli,T.G.,&Folk,C.L.(1996).Spatial cuing in a stereoscopic display:Evidence for a“depth-blind”attentional spotlight.Psychonomic Bulletin&Review,3
(1),81–86.Graziano,M.S.,&Cooke,D.F.(2006).Parieto-frontal interactions,personal space, and defensive behavior.Neuropsychologia,44
(13),2621–2635.Harris,J.M.,&Drga,V.F.(2005).Using visual direction in three-dimensional motion perception.Nature Neuroscience,8
(2),229–233.Hayman,R.,Verriotis,M.A.,Jovalekic,A.,Fenton,A.A.,&Jeffery,K.J.(2011).Anisotropic encoding of three-dimensional space by place cells and grid cells.Nature Neuroscience,14
(9),1182–1188.Iavecchia,H.P.,&Folk,C.L.(1994).Shifting visual attention in stereographic displays:A time course analysis.Human Factor,36
(4),606–618.Janssen,P.,Vogels,R.,&Orban,G.A.(2000).Selectivity for 3D shape that reveals distinct areas within macaque inferior temporal cortex.Science,288
(5473),2054–2056.Klein,R.M.(2000).Inhibition of return.Trends in Cognitive Sciences,4
(4),138–147.Klein,R.M.,&MacInnes,W.J.(1999).Inhibition of return is a foraging facilitator in visual search.Psychological Science,10
(4),346–352.Klier,E.M.,Wang,H.Y.,&Crawford,J.D.(2001).The superior colliculus encodes gaze commands in retinal coordinates.Nature Neuroscience,4
,627–632.LaBerge,D.,&Brown,V.(1989).Theory of attentional operations in shape identification.Psychological Review,96
(1),101–124.Mangun,G.R.,&Hillyard,S.A.(1988).Spatial gradients of visual attention:Behavioral and electrophysiological evidence.Electroencephalography and Clinical Neurophysiology,70
(5),417–428.Maringelli,F.,McCarthy,J.,Steed,A.,Slater,M.,&Umiltà,C.(2001).Shifting visuo-spatial attention in a virtual three-dimensional space.Cognitive Brain Research,10
(3),317–322.Maruff,P.,Yucel,M.,Danckert,J.,Stuart,G.,&Currie,J.(1999).Facilitation and inhibition arising from the exogenous orienting of covert attention depends on the temporal properties of spatial cues and targets.Neuropsychologia,37
(6),731–744.Mathôt,S.,&Theeuwes,J.(2010).Gradual remapping results in early retinotopic and late spatiotopic inhibition of return.Psychological Science,21
(12),1793–1798.Maylor,E.A.,(1985).Facilitatory and Inhibitory components of orienting in visual space.In:M.I.Posner&O.S.Marin(Eds.),Attention and Performance XI
(pp.189–204).Hillsdale,NJ:Lawrence Erlbaum Associates.Maylor,E.A.,&Hockey,R.(1987).Effects of repetition on the facilitatory and inhibitory components of orienting in visual space.Neuropsychologia,25
(1A),41–54.McAuliffe,J.,&Pratt,J.(2005).The role of temporal and spatial factors in the covert orienting of visual attention tasks.Psychological Research,69
(4),285–291.McDonald,J.J,Hickey,C.,Green,J.J,&Whitman,J.C.(2009).Inhibition of return in the covert deployment of attention:Evidence from human electrophysiology.Journal of Cognitive Neuroscience,21
(4),725–733.Mele,S.,Savazzi,S.,Marzi,C.A.,&Berlucchi,G.(2008).Reaction time inhibition from subliminal cues:Is it related to inhibition of return?.Neuropsychologia,46
(3),810–819.Posner,M.I.(1984).Current research in the study of selective attention.In E.Donchin(Ed.),Cognitive psychology:Event-related potentials and the study of cognition
(Vol.2,pp.37–50).Hillsdale,N.J.:Lawrence Erlbaum.Posner,M.I.,Davidson,B.J.&Snyder,C.R.R.(1980).Attention and the detection of signals.Journal of Experimental Psychology:General,109
(2),160–174.Posner,M.I.,Rafal,R.D.,Choate,L.&Vaughan,J.(1985).Inhibition of return:Neural basis and function.Cognitive Neuropsychology,2
(3),211–228.Pratt,J.,Adam,J.,&McAuliffe,J.(1998).The spatial relationship between cues and targets mediates inhibition of return.Canadian Journal of Experimental Psychology,52
(4),213–216.Pratt,J.,&Abrams,R.A.(1999).Inhibition of return in discrimination tasks.Journal of Experimental Psychology:Human Perception and Performance,25
(1),229–242.Pratt,J.,Hillis,J.,&Gold,J.M.(2001).The effect of the physical characteristics of cues and targets on facilitation and inhibition.Psychonomic Bulletin&Review,8
(3),489–495.Pratt,J.,O'donnell,C.,&Morgan,A.(2000).The role of the fixation location in inhibition of return.Canadian Journal of Experimental Psychology,54
(3),186–195.Previc,F.H.(1998).The neuropsychology of 3-D space.Psychological Bulletin,124
(2),123–164.Sereno,M.E.,Trinath,T.,Augath,M.,&Logothetis,N.K.(2002).Three-dimensional shape representation in monkey cortex.Neuron,33
(4),635–652.Shen,M.W.,Gao,Z.F.,Zhang,G.Q.,Shui,R.D.,Qiao,X.X.,&Li,W.J.(2007).Inhibition of return on three-dimensional slant plane.Acta Psychologica Sinica,39
(6),951–958.[沈模卫,高在峰,张光强,水仁德,乔歆新,李伟健.(2007).三维倾斜平面的返回抑制.心理学报,39
(6),951–958.]Standen,P.J.,&Brown,D.J.(2005).Virtual reality in the rehabilitation of people with intellectual disabilities:review.Cyberpsychology&Behavior,8
(3),272–282.Theeuwes,J.,&Pratt,J.(2003).Inhibition of return spreads across 3-D space.Psychonomic Bulletin&Review,10
(3),616–620.Yamane,Y.,Carlson,E.T.,Bowman,K.C.,Wang,Z.H.,&Connor,C.E.(2008).A neural code for three-dimensional object shape in macaque inferotemporal cortex.Nature Neuroscience,11
(10),1352–1360.Zhang,M.,&Zhang,Y.(2006).Object based-inhibition of return in dynamic displays:Object inhibition or updating of spatial inhibition?.Acta Psychologica Sinica,38
(6),798–804.[张明,张阳.(2006).动态范式中基于客体的返回抑制-是客体抑制还是空间抑制的动态更新?.心理学报,38
(6),Zhang,Y.,Peng,C.H.,Sun,Y.,&Zhang,M.(2013).Cognitive mechanism of visual inhibition of return.Advances in Psychological Science,21
(11),1913–1926.[张阳,彭春花,孙洋,张明.(2013).视觉返回抑制的认知机制.心理科学进展,21
(11),1913–1926.]798–804.]
Zhang,Y.,Zhou,X.L.,&Zhang,M.(2012).Temporary inhibitory tagging at previously attended locations:Evidence from event-related potentials.Psychophysiology,49
(9),1191–1199.猜你喜欢
体育科技(2022年2期)2022-11-07
幼儿教育·教育教学版(2020年8期)2020-12-23
幼儿教育·教育教学版(2020年8期)2020-12-23
幼儿教育·教育教学版(2020年8期)2020-12-23
中学生数理化·高三版(2019年1期)2019-07-03
中学生数理化·高三版(2019年1期)2019-07-03
炎黄地理(2018年7期)2018-10-20
阅读(科学探秘)(2018年4期)2018-05-14
试题与研究·高考数学(2016年1期)2016-10-13
现代装饰·理论(2016年1期)2016-03-01
