
人类大脑结构和功能的性别差异:来自脑成像研究的证据*
2015-02-01 01:50杨天亮
心理科学进展 2015年4期
杨天亮 辛 斐 雷 旭
(认知与人格教育部重点实验室(西南大学); 西南大学心理学部, 重庆 400715)
1 引言
近年来, 认知神经科学发展迅速, 取得了丰硕的成果。性别作为影响人类认知和行为的重要因素, 也成为了认知神经科学领域关注的话题。不同于个体差异, 性别差异专指男性群体和女性群体之间某种稳定的差异, 是群体水平上的比较。而个体差异是个体在成长过程中由于先天和后天因素所表现出的相对稳定而又不同于他人的特点, 是基于个体水平的比较(Willerman, 1979)。对于认知和行为方面的性别差异, 研究表明男性较女性具有更高的空间认知能力, 而女性在言语能力上(如言语流畅性、言语记忆等)较男性更胜一筹(Halpern, 2013; Hamilton, 2008)。众所周知, 大脑是认知和行为表现的基础, 因此对大脑结构和功能性别差异的考察是理解认知和行为层面性别差异的关键。
早期脑解剖研究发现, 男性和女性在脑结构上存在差异, 比如男性大脑体积显著大于女性;女性胼胝体尾部呈球状, 而男性呈圆柱状等(Cosgrove, Mazure, & Staley, 2007; Hines, 2004)。这些研究虽然为脑性别差异的理解奠定了基础,但是由于方法学上的限制(如死亡原因、死亡时间等), 使得研究结果的信度和效度受到一定的影响。幸运的是, 现代脑成像技术的进步为我们研究健康人类大脑结构和功能的性别差异开辟了新的道路。对脑结构性别差异的研究, 通常采用结构磁共振成像(Structural magnetic resonance imaging,sMRI)以及弥散张量成像(Diffusion tensor imaging,DTI); 对脑功能性别差异的考察, 通常采用脑电图(Electroencephalogram, EEG)、正电子发射断层扫描技术(Positron emission tomography, PET) 以及功能性磁共振成像(Functional magnetic resonance imaging, fMRI)。在脑功能方面, 值得关注的是静息态下大脑的性别差异。静息态是指受试者清醒、平静呼吸、最大限度地减少身体主动与被动运动、以及尽量不做任何思维活动的状态。这种状态刻画的是大脑内部自发产生(与外界刺激无关)的神经活动。并且研究表明这种自发脑活动的时空特性表征了大脑功能活动的基本框架, 且与各种认知活动密切相关(Gusnard & Raichle, 2001)。由于静息态应用简单、可操作性好, 并且可以避免由于任务设计和被试反应所导致的研究结果的不可对比性等特点, 脑功能性别差异的静息态研究近年来成为不少研究者关注的焦点。
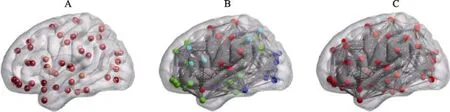
图1 大脑性别差异研究的三个层次:脑局部区域, 脑子系统以及全脑连接组。A是对脑局部区域的描述, 图中的每个节点表示一个独立的脑区; B是对脑子系统的描述, 图中每一种颜色的节点集之间的连接组成了一个脑子系统; C是对全脑连接组的描述, 图中所有节点之间的连接构成了一个全脑结构/功能连接组。
功能分化和功能整合作为大脑的两大组织原则, 大脑在局部功能特殊化和脑区间的相互作用之间达到了很好的平衡(Friston, 2002)。虽然不同的脑区具有相对不同的功能, 但即使完成一项十分简单的任务, 也总是需要多个脑区相互协作,形成一个脑子系统发挥其功能。例如完成大脑自发认知功能和外部环境监控功能的默认网络(default mode network, DMN), 就是由内侧前额叶、楔前叶、后扣带回以及内侧颞叶这些联系紧密的脑区构成的(Smith et al., 2009; 李雨, 舒华,2014)。而当完成一项比较复杂的任务时, 可能需要全脑相互作用、互相协调, 共同构成一个全局网络(即全脑连接组)来发挥其功能(见图1)。目前,利用脑成像技术, 已有大量工作发现脑局部区域、脑子系统以及全脑连接组都存在着广泛的性别差异。复杂网络技术作为分析全脑连接组内在属性的一种手段, 为我们揭示脑性别差异提供了一个新的视角。该技术将大脑看作包含大量节点和边(即脑连接)的网络, 并采用图论中的各种指标(如全局效率、局部效率等)来刻画脑连接组织模式的各种属性, 其中脑网络中的节点表征不同的解剖区域或功能区域, 边表征节点之间的相互作用(Sporns, 2011)。
此外, 脑性别差异是发展变化的, 即在不同的年龄段, 性别差异的程度有所不同(Gong, He,& Evans, 2011)。而且已有研究提示, 脑性别差异是基因和环境共同作用的产物。动物研究显示产前激素、性染色体等对脑性别差异的形成具有重要的影响(Arnold & Breedlove, 1985; Arnold &Chen, 2009)。同时, 脑性别差异还受性别特异性的基因表达, 类固醇激素以及后天因素(如早期儿童护理、成长环境、学习训练等)的影响(Cicchetti,2013; Giedd, Raznahan, Mills, & Lenroot, 2012; Kang et al., 2011; 邹吉林, 王美芳, 曹仁艳, 闫秀梅,2009)。本文通过对研究文献的梳理与总结, 首先探讨了脑性别差异领域的一些认识误区, 接着在脑局部区域、脑子系统以及全脑连接组三个层面,对静息态脑功能和脑结构的性别差异及其在年龄上的发展变化的相关研究进行了综述, 最后简要展望了未来可能的研究方向。
2 脑性别差异领域的一些认识误区
目前, 人类对大脑性别差异的认识仍存在不少误区。走出这些误区, 有助于我们更好地理解性别对大脑的影响。第一个误区是, 性别对大脑的影响是微小且不稳定的。在行为学研究中, 我们经常发现男女在某些认知任务上是不存在性别差异的(Grabowski, Damasio, Eichhorn, & Tranel,2003; Piefke, Weiss, Markowitsch, & Fink, 2005),比如 Grabowski等人(2003)发现男女在图像命名能力上不存在差异, Piefke等人(2005)在考察自传体记忆检索能力时也未能发现性别差异。考虑到大脑是认知和行为表现的基础, 研究者认为性别对大脑几乎是没有影响的。然而随着脑成像技术的发展, 研究逐渐发现大脑存在着广泛的性别差异(Chou, Cheng, Chen, Lin, & Chu, 2011; Kawachi et al., 2002; Lü et al., 2010), 比如Chou等人(2011)基于弥散张量成像, 发现女性海马、扣带回以及胼胝体等区域的各向异性分数显著高于男性;Kawachi等人(2002)基于正电子发射断层扫描技术, 发现男性颞中回和内侧额叶的葡萄糖代谢强度显著高于女性。
第二个误区是, 同一性别内个体的大脑表现出明显的差异, 并且这种个体差异大大弱化了大脑的性别间差异。由于遗传和环境的交互影响,男性和女性的大脑都表现出明显的个体差异, 例如 Schmithorst, Holland和 Dardzinski (2008)发现同一性别内不同个体的胼胝体体积存在着显著差异。因此在性别差异的研究中, 个体差异对性别差异的影响程度一直是研究者争论的焦点。有研究者使用统计分析手段(如方差分析)发现了人类大脑存在着显著的个体差异, 然而大脑之间的性别差异却不显著(Lüders, Steinmetz, & Jäncke,2002; Larsen et al., 2006; Larsen, Hermans, & Engels,2012), 例如Lüders等人(2002)通过分析100名健康被试的大脑(50名男性, 50名女性), 发现脑体积越大, 大脑的灰质比例就越小, 然而脑体积与灰质的这种关系在性别上是没有差异的。但也有研究者通过对研究流程的控制和研究方法的创新,控制了个体差异对性别差异的影响, 认为性别内的个体差异并不足以改变性别间脑差异的趋势(Im et al., 2008; Leonard et al., 2008), 例如Leonard等人(2008)使用协方差分析的方法, 发现女性颞横回的体积较男性大等。随着科学的进步, 男女两性在大脑上的差异已得到公认。Cahill (2006)通过总结脑性别差异研究的现状, 发现大脑存在着广泛的性别差异, 例如男性的脑体积比女性大,女性的灰质百分比大于男性等, 所以Cahill (2006)认为个体间差异即便存在, 也可能居于次要地位。
最后一个误区是, 如果某种行为没有性别差异的话, 那么这种行为的神经基础也不存在性别差异。在没有出现脑成像技术以前, 通过对脑损伤病人的研究, 早期研究者认为如果某种行为没有性别差异的话, 那么这种行为的神经基础也没有性别差异(Hines, 2004)。然而随着脑成像技术的出现和迅速发展, 大量研究发现即使男性和女性在某种行为表现上没有差异, 但这种行为的神经基础也可能存在着显著的性别差异(Hugdahl, Thomsen,& Ersland, 2006), 例如Hugdahl等人(2006)考察了男性和女性在心理旋转能力上的神经基础。虽然所选取的男女被试在心理旋转任务上的行为表现没有差异, 但是研究发现在进行心理旋转任务时,男性在大脑顶叶显示出了强烈的激活, 而女性则在额下回显示出了强烈的激活。
3 脑结构性别差异的探索发现
回顾脑结构性别差异的研究, 我们发现脑灰质和脑白质在脑局部区域、脑子系统以及全脑结构连接组层面上存在着广泛的性别差异(见表1)。而且脑结构性别差异并不是一成不变的, 在不同的年龄段上其差异程度有所不同。
脑灰质和白质分别富含神经元胞体和神经纤维(或神经元间的连接)。根据研究假设对胞体和连接的强调程度, 可以采用不同的模态加以探讨。对脑灰质性别差异的研究主要采用结构磁共振成像, 基于该技术我们可以利用灰质体积(通常用某一脑局部区域内的体素个数表示, 描述的是绝对体积)、皮层厚度(指大脑皮层上的灰质内表面与外表面之间的距离)以及灰质密度(反映了神经元、树突和突触的数量情况)等指标去定量刻画脑灰质信息, 进而考察脑结构的性别差异(见表1)。结构磁共振成像的优势在于它对脑组织的分辨率高,并且所获得的解剖关系十分明确。对脑白质性别差异的研究主要采用弥散张量成像, 该技术利用水分子在不均匀组织中弥散具有各向异性的特性来描述白质结构及其潜在的纤维束, 并在三维空间内定量分析组织内水分子的弥散活动。采用该技术所探讨的脑性别差异暗示着神经信息传递以及结构连接之间的差异。弥散张量成像技术的优势在于成像参数多, 包含的信息量大。利用弥散张量成像产生的矢量参数可以定量分析脑结构的性别差异(见表1)。常见的矢量参数主要有反映水分子平均弥散能力的平均弥散率(Mean diffusivity,MD), 和反映水分子空间弥散方向的各向异性分数(Fractional anisotropy, FA)。
3.1 脑结构的性别差异
基于结构磁共振成像数据, 研究者利用灰质体积、皮层厚度以及灰质密度指标分析了脑结构在脑局部区域上的性别差异, 所得结论基本一致。利用灰质体积指标, 研究发现女性额叶、顶叶和枕叶某些区域的灰质体积显著大于男性(Allen, Damasio, Grabowski, Bruss, & Zhang, 2003;Chen, Sachdev, Wen, & Anstey, 2007; Ruigrok et al.,2014)。比如 Chen等人(2007)以 411名 44到 48岁的健康成年人为研究对象, 发现女性右侧顶下小叶的灰质体积显著大于男性。Ruigrok等人(2014)在对以往关于脑结构性别差异的研究进行元分析时发现, 女性额叶(如额下回、额中回等)、枕叶(如枕叶皮层外侧等)以及顶叶(如顶叶盖部等)的灰质体积显著大于男性。随着结构磁共振成像数据处理技术的进步, 研究者进而考察了皮层厚度以及灰质密度在脑局部区域性别差异上的表现。利用这些指标, 研究发现女性额叶、顶叶某些区域的灰质密度和皮层厚度显著大于男性(Luders et al., 2006; Lü et al., 2010; Wang, Shen,Tang, Zang, & Hu, 2012)。比如, Lü 等人(2010)以184名18到67岁的健康成年人为研究对象, 发现女性额叶和顶叶某些区域的皮层厚度显著大于男性, 如额上回、顶上小叶等。此外, Wang等人(2012)以140名18到26岁的健康成年人为研究对象, 发现女性额上回的灰质密度显著高于男性。

表1 有关脑结构和功能在脑局部区域、脑子系统以及全脑连接组层面上性别差异的研究
在脑局部区域层次上, 来自弥散张量成像技术的研究表明女性海马、扣带回以及胼胝体等区域具有更高的各向异性, 存在这样差异的原因主要是女性这些区域的髓鞘化程度较男性更高(Chou et al., 2011; Menzler et al., 2011)。例如, Menzler等人(2011)以 25名健康成年男性(31.8±8.6岁)和25名健康成年女性(31.1 ± 10.6岁)为研究对象,发现女性海马、扣带回以及胼胝体的各向异性分数显著高于男性。此外, Chou等人(2011)以40名健康成年男性(25.7 ± 6.4岁)和40名健康成年女性(25.9 ± 7.2岁)为研究对象也发现女性海马、扣带回以及胼胝体的各向异性分数较男性高。
众所周知, 大脑是一个整体, 各个脑区之间并不是孤立存在的, 而是相互协作的。因此, 单从脑局部区域探讨脑结构的性别差异可能会忽略掉脑区之间交互作用的信息, 即脑连接信息。由于弥散张量成像可以直观地刻画脑区之间真实的结构连接, 因此研究多采用弥散张量成像去探讨结构连接的性别差异(见表1)。通过回顾脑结构连接性别差异的研究, 我们发现当前研究多在全脑连接组层面进行探索。例如Ingalhalikar等人(2014)以949名8到22岁的参与者为研究对象, 在全脑连接组层面探讨了脑连接的性别特异性。研究人员利用弥散张量成像技术, 通过白质纤维跟踪的方法, 分析了脑区之间复杂的相互作用, 并且对每个大脑的结构连接组建模。结果显示, 在全脑结构连接组层次, 男性大脑幕上区域半球内连接显著强于女性, 而女性半球间连接显著强于男性。大脑的这种连接模式使得男性大脑可以把出现脑后方的知觉感受和由脑前方控制的行为意向连接起来, 这样男性的大脑就形成了一个促进协调运动的有效系统; 相比之下, 女性大脑的连接模式很可能促进了左半球的分析和逻辑推理能力与右半球的空间、直觉信息处理能力之间的整合,这样就使得女性更加擅长语言情感的表达, 并且也更加感性。此外, 在全脑结构连接组层次, 研究还发现女性脑结构网络在脑局部水平上的信息传递效率显著高于男性(Gong et al., 2009; Yan et al.,2011)。例如, Gong等人(2009)以95名19到85岁的健康成年人为研究对象, 利用 AAL (automated anatomical labeling, AAL)模板 (Tzourio-Mazoyer et al., 2002) 将大脑分为78个脑区构建了加权的脑结构网络, 发现女性脑结构网络较男性具有更高的局部效率。此后, Yan等人(2010)以73名18到27岁的健康成年人为研究对象, 也证实了女性脑结构网络在脑局部水平上的信息传递效率较男性更为高效。
结构脑成像研究表明, 脑性别差异在脑局部区域和全脑连接组层面上广泛存在, 并且在脑局部区域层面上的研究结果基本一致, 比如女性额叶、顶叶和枕叶某些区域的灰质体积显著大于男性。但是我们应该注意到当前有些研究所选取样本的样本数量相对较小, 比如Menzler等人(2011)的研究仅选取了50名被试, 这可能会影响研究结果的统计效力。此外, 当前关于脑结构性别差异的研究主要集中在脑局部区域层面, 而对于脑连接(脑子系统和全脑连接组)性别差异的研究较少,未来应该加强这方面的研究以期对脑结构的性别差异有一个更加精确、全面的了解。总之, 当前脑结构性别差异的证据启示我们, 在进行脑结构的相关研究时应该将性别作为一个变量进行分析。
3.2 脑结构性别差异在年龄上的发展变化
通过对既有研究资料的综述可以发现, 在正常的发育和老化过程中, 男女大脑表现出不同的发展轨迹, 并且脑结构性别差异的发展变化既存在于脑局部区域层面又存在于脑连接层面(全脑结构连接组)。
在脑局部区域层面上, 基于结构磁共振成像技术, 多数研究发现额叶、颞叶、顶叶以及小脑的灰质体积存在着显著的年龄与性别的交互作用(Allen et al., 2003; Sowell et al., 2007; Xu et al.,2000)。例如, Sowell等人(2007)以176名7到87岁的健康成年人为研究对象发现, 男性背侧前额叶皮层和颞下回灰质体积的萎缩程度显著高于女性。Xu等人(2000)以331名30到79岁的健康成年人为研究对象发现, 随着年龄增长, 男性前额叶皮层后部右侧区域灰质体积的萎缩程度显著高于女性; 而且随着年龄增长, 颞叶中部右侧、顶叶以及小脑只在男性被试上出现了萎缩现象。利用弥散张量成像技术, 目前研究发现女性儿童右半球大部分脑区的各向异性分数与年龄呈正相关, 而男性儿童左半球大部分脑区的各向异性分数与年龄呈正相关(Schmithorst & Holland, 2007; Schmithorst et al., 2008)。Schmithorst等人(2008)以 106名 5到18岁的健康未成年人为研究对象, 探讨了脑白质的性别差异在童年和青少年阶段的发展趋势。结果显示, 女性儿童右侧额叶、右侧弓状束等区域的各向异性分数随着年龄的增长而增长, 男性儿童则是左侧额叶等区域的各向异性分数随着年龄的增长而增长。
目前, 利用结构连接信息对脑结构性别差异发展变化的研究较少, 基本没有脑子系统方面的相关研究。但是在全脑结构连接组层次上,Ingalhalikar等人(2014)利用弥散张量成像, 研究了8到22岁的949名健康被试在发育过程中脑连接的性别特异性。结果发现, 13岁之前男女脑结构连接几乎没有什么差异, 但在 14到 17岁的青春期间, 脑连接的性别差异迅速扩大, 到了成年早期男性表现出显著的半球内连接优势, 女性则表现出显著的半球间连接优势(Ingalhalikar et al.,2014), 这可能意味着青春期是男性半球内连接优势和女性半球间连接优势形成的一个关键时期。
以上研究表明, 脑局部区域和脑连接(全脑结构连接组)的性别差异都存在着年龄上的发展变化。但是现有的研究缺乏在脑子系统层面对脑结构性别差异发展变化的考察。此外, 既有研究基本都是横向研究, 缺少纵向研究, 而且大都只是针对人生的某个时期(比如儿童或青少年时期等),而没有涵盖人的整个发展历程, 因此很难对脑结构性别差异发展变化这一完整的动态过程进行梳理。然而, 弄清脑结构性别差异的发展变化是很有必要的, 它可以引出一系列的研究问题, 比如男女脑结构性别差异是从什么时候开始的?什么时候形成稳定的脑结构性别差异?……解答这些问题有助于更好地理解认知和行为层面性别差异的发展变化。同时, 脑结构性别差异发展变化的研究启示我们, 在进行脑结构性别差异的研究时应该考虑研究对象的年龄特征。
4 脑功能性别差异的探索发现
作为脑性别差异研究的另外一个重要领域,脑功能在脑局部区域、脑子系统以及全脑连接组层面上也存在着广泛的性别差异(见表1), 并且脑功能性别差异也存在着年龄上的发展变化。静息态下脑功能性别差异的研究多采用脑电图、正电子发射断层扫描技术以及功能性磁共振成像。其中脑电图通过记录大脑自发性、节律性的电活动来定量描述脑功能活动, 具有较高的时间分辨率。然而目前利用脑电图探讨静息态脑功能性别差异的研究比较少(见表1)。正电子发射断层扫描技术通过追踪放射性物质在大脑中的代谢情况来定量分析大脑的功能活动, 具有较高的灵敏度、特异性以及空间分辨率。基于该技术我们可以利用局部脑血流量和葡萄糖代谢强度等指标来定量描绘脑功能活动, 进而考察脑功能的性别差异。但是利用正电子发射断层扫描技术, 现有的研究多用于考察脑局部区域的性别差异(见表1)。功能性磁共振成像技术通过分析神经元活动对脑局部血氧和血流的影响所引起的脑局部磁场的变化来定量描述大脑的功能活动, 兼有较高的空间分辨率。采用该技术多在功能连接层面进行脑性别差异的考察(见表1), 其中功能连接指的是脑区的时间序列在时间上的统计依赖性。
4.1 脑功能的性别差异
在脑局部区域层面上, 基于正电子发射断层扫描技术, 研究者利用局部脑血流量和葡萄糖代谢强度指标考察了脑功能性别差异, 但是这方面的研究结论并不一致(Andreason, Zametkin, Guo,Baldwin, & Cohen, 1994; George, Ketter, Parekh,Herscovitch, & Post, 1996; Kawachi et al., 2002)。George等人(1996)以6名健康成年男性(34.5±12.1岁)和 6名健康成年女性(35±8.8岁)为研究对象,使用基于体素的形态测量法发现男性颞叶和额叶的局部脑血流量显著高于女性, 而女性脑干的局部脑血流量显著高于男性。Kawachi等人(2002)以44名47到80岁的健康成年人为研究对象, 采用基于体素的形态测量法, 利用葡萄糖代谢强度指标也发现了男性的颞叶和额叶优势, 具体表现为男性颞中回和内侧额叶的葡萄糖代谢强度显著高于女性。然而基于感兴趣区域的方法, Andreason等人(1994)以39名19到35岁的健康成年被试为研究对象, 发现女性额叶的葡萄糖代谢强度显著高于男性。研究结果的不一致首先可能源于图像分析方法的不同:George等人(1996)和 Kawachi等人(2002)的研究所采用的图像分析方法是基于体素的形态测量法, 这种方法是通过对不同图像中的体素进行逐个比对, 然后在脑成像中确定有差异的脑区; 而Andreason等人(1994)的研究采用的则是基于感兴趣区域的方法, 这种方法首先是根据自己的理论假设确定有差异的脑区, 然后对该脑区进行分析。其次, 不同研究所使用的被试数量是不同的, 小的样本量会降低研究结果的统计效力。如 George等人(1996)的研究仅采用 12名被试, 而Andreason等人(1994)的研究使用了39名被试。再之, 研究间所使用被试的年龄范围也是不一致的, 这样就降低了不同研究之间的可比性。例如Kawachi等人(2002)的研究所采用被试的年龄范围是47到80岁, 而Andreason等人(1994)研究的被试年龄范围是19到35岁。最后需要提及的是, Reiman等人(1996)发现女性大脑的葡萄糖代谢强度在不同的月经周期阶段显著不同, 这也有可能是造成上述研究结果不一致的一个原因(Reiman, Armstrong, Matt, & Mattox, 1996)。
与脑结构性别差异的研究一样, 从脑连接的角度探讨脑功能子系统以及全脑功能连接组的性别差异也是有必要的。在脑子系统层面, 研究结论并不一致(Biswal et al., 2010; Weissman-Fogel,Moayedi, Taylor, Pope, & Davis, 2010; Zuo et al.,2010)。利用1414名年龄大于18岁并且平均年龄小于 60岁的健康成年被试的静息态功能性磁共振数据, Biswal等人(2010)使用时间独立成分分析的方法发现, 女性后扣带回、前额叶皮层内侧以及顶下小叶的功能连接强度显著高于男性, 而扣带回背侧、脑岛、颞上回、额叶内侧上部以及枕叶的功能连接强度显著低于男性。然而,Weissman-Fogel等人(2010)利用49名25到50岁的健康成年被试的功能性磁共振数据, 使用空间独立成分分析的方法却没有发现默认网络、执行控制网络以及突显网络存在任何脑连接上的性别差异, 因此 Weissman-Fogel等人(2010)认为静息态功能连接的研究完全可以忽略性别的影响。以上研究结果的不一致可能源于研究所使用被试年龄范围和被试数量的差异。Biswal等人(2010)的研究所采用的被试数量为1414名, 并且被试的年龄范围几乎跨越人的整个生命周期, 而 Weissman-Fogel等人(2010)的研究仅采用了49名被试, 而且被试的年龄范围只包含青年期和中年期。此外,研究间所使用的数据分析方法也是不一样的, 比如 Biswal等人(2010)的研究使用的是时间独立成分分析的方法, 而 Weissman-Fogel等人(2010)的研究使用的则是空间独立成分分析的方法。空间独立成分分析在一定程度上是用时间的独立性换取空间的独立性, 进而获取表现相互独立成分的图像和这些独立成分相关的时序波形; 而时间独立成分析方法则是用空间的独立性换取时间的独立性, 进而获取独立成分的时序波形和与其相对应的图像。
在全脑功能连接组层面上, 研究者利用静息态功能性磁共振技术探讨了大脑在网络属性上的性别差异, 但是这方面的研究结论存在分歧 (Wang et al., 2008; Wu et al., 2013)。Wu 等人(2013)以 291名5.6到18.4岁的健康儿童和青少年为研究对象,基于AAL模板将全脑分为90个区域用于构建二值的脑功能网络, 结果发现男性脑功能网络较女性具有更高的全局效率和局部效率。然而 Wang等人(2008)以20名21到25岁的健康成年人为研究对象, 采用同样的方法, 却未能发现脑网络在全局效率和局部效率上的性别差异。研究结果之间的分歧可能是由于样本大小以及被试年龄范围的差异。Wu等人(2013)的研究采用的被试数量为291名, 并且所采用的被试是不大于18岁的儿童和青少年, 而 Wang等人(2008)的研究仅有 20名被试, 并且被试主要是21到25岁的青年。
综上所述, 脑功能性别差异在脑局部区域、脑子系统以及全脑连接组层面上广泛存在, 这提示我们未来在进行脑功能的有关研究时应该纳入性别这一变量。通过讨论分析, 我们发现研究结论之间的不一致可能受样本大小、样本年龄范围、数据分析手段等差异的影响。未来研究应该着重考虑这些因素对研究结果的影响, 以期获得脑功能性别差异更一致的研究结论。此外, 女性月经周期对脑功能的影响也启示我们在研究脑功能性别差异时, 应该采集女性的月经周期信息, 以期获得更加精确、可靠的研究结论。
4.2 脑功能性别差异在年龄上的发展变化
脑功能作为对脑结构上不同神经元、神经元集群或者脑区动态活动的一种直观描述, 同脑结构性别差异一样, 脑功能的性别差异也存在着发展变化。并且脑功能性别差异的发展变化既存在于脑局部区域又存在于脑连接(脑子系统)。在脑局部区域层次上, 利用正电子发射断层扫描技术,研究表明左侧丘脑、左侧尾状核以及下丘脑的葡萄糖代谢强度存在着显著的年龄与性别的交互作用(Kawachi et al., 2002; Murphy et al., 1996)。例如,Kawachi等人(2002)以44名47到80岁的健康成年被试为研究对象, 发现男性左侧丘脑的葡萄糖代谢强度随着年龄的增长而显著增长, 而女性左侧尾状核和下丘脑的葡萄糖代谢强度随着年龄的增长而显著下降。
利用脑连接信息, 研究者对脑功能性别差异的发展变化进行了初步的探索。空间对称的大脑左右半球内的神经元内源性的自发活动具有高度的相似性(即, 功能同伦), 这使得左、右侧脑半球在认知和行为功能上维持基本一致(Zuo et al.,2010)。基于体素-镜像同伦连接的方法可用于检测这一特征, 反映脑半球间的信息交流整合。Zuo等人(2010)应用基于体素-镜像同伦连接这一方法,通过分析214名7到85岁健康被试的静息态功能性磁共振数据, 探讨了大脑左右半球间的同伦功能连接性别差异的发展趋势。结果发现男性背外侧前额叶皮层的功能同伦性随着年龄的增长而增强, 而女性则表现出随着年龄的增长而减弱; 男性杏仁体的功能同伦性随着年龄的增长而减弱,而女性却随着年龄的增长而增强(Zuo et al., 2010)。α波作为正常人脑电波的基本节律, 在人清醒、安静并闭眼时这种节律更为明显。考虑到α波的这种优势, Barry等人(2004)利用静息态 EEG技术,研究了80名8到12岁的健康儿童在发育过程中的α波相干性的性别特异性。结果显示男性儿童颞叶区域α波相干性从8岁到9岁缓慢下降, 12岁之前又加速上升; 然而, 女性儿童颞叶区域 α波相干性在9岁时缓慢下降, 10岁或11岁快速增强并达到顶峰, 之后又呈现下降的趋势(Barry et al.,2004)。
总而言之, 脑功能性别差异的发展变化既存在于脑局部区域又存在于脑连接(脑子系统), 这启示我们在考察脑功能性别差异时应该考虑被试的年龄特征。但是此类研究较少, 甚至没有研究从全脑功能连接组的角度去探讨脑功能性别差异的发展变化。此外, 有些研究所采用被试的年龄跨度比较小, 并且被试的数量较少, 如 Kawachi等人(2002)的研究仅采用了 44名被试, 并且被试年龄只涵盖中年期和老年期:小的样本数量影响研究结果的统计效力; 窄的被试年龄跨度使得我们不能对脑功能性别差异这个发展变化历程有一个整体的了解。同时, 关于脑性别差异发展变化的研究基本都是横向研究, 此类研究不能保证组群的可比性, 并且也不能观测到同一个体发展的连续性。
5 总结与展望
尽管一些具体结论还存在着分歧, 但脑性别差异的研究到目前为止取得了丰硕的成果:脑性别差异广泛存在于脑局部区域、脑子系统以及全脑连接组; 脑性别差异存在着年龄上的发展变化;脑性别差异是基因和环境共同作用的结果等。总的来说, 脑性别差异的研究启示我们在研究人类大脑时, 应该将性别作为一个变量进行分析。
虽然已有的成果在一定程度上推进了脑性别差异的研究, 但仍存在着一些不足之处。首先, 在对脑性别差异发展变化性进行考察时, 现有的研究多是横向研究, 此类研究往往不能保证组群间的可比性, 并且不能观测同一个体发展的连续性。这可能会影响研究结果的可靠性。其次, 既有的关于脑性别差异的研究多是数据驱动的。这类研究往往缺少一些至关重要的变量, 比如女性被试的月经周期阶段等, 这可能会使研究结果的科学性降低。再次, 不同研究中所使用的样本量差异较大, 这也很容易造成研究结果的不一致,并且小的样本数量容易影响研究结果的统计效力。最后, 脑子系统以及全脑连接组的构建是基于脑连接信息, 目前研究中所采用的脑连接信息大多是没有方向的。这种分析虽然简单, 但是忽略了脑连接中重要的方向信息(Lei et al., 2011)。
未来的研究应该从以下几个方面进行加强:第一, 当前研究主要集中于健康被试的脑性别差异, 然而很多心理疾病呈现出性别上的二态性,比如男性和女性精神分裂症患者的起病年龄、症状病程演变特点等都存在着差异(Falkenburg &Tracy, 2014)。虽然已有少量研究开始探讨具有性别特异性的心理疾病的脑机制, 但是研究还不够充分。未来应该加强对具有性别特异性的心理疾病脑机制的研究, 以期提高研究的临床价值和科学意义。第二, 已有研究提示产前激素、性染色体、早期儿童护理以及成长环境等对脑性别差异的形成具有重要的影响(Arnold & Chen, 2009;Cicchetti, 2013; Giedd et al., 2012; Kang et al., 2011),这似乎反映了基因和环境的共同影响。可是, 基因和环境各自在多大程度上影响着脑性别差异?基因和环境是如何交互影响脑性别差异的?今后有关双生子的研究将有助于进一步揭示基因和环境对脑性别差异的影响程度及其交互作用。第三,大脑的功能活动是一个动态变化的过程, 该过程的发生是十分迅速的, 时间在几百毫秒内(Biswal et al., 2010; Mizraji & Lin, 2011)。现有的关于脑功能性别差异的研究都是在比较长的时间尺度上(秒、分钟等)进行的。如何在更小的时间尺度上去探究脑功能的性别差异也是未来的一个研究方向。第四, 在探讨脑性别差异时, 采用多模态的手段, 比如将高时间分辨率的脑电图和高空间分辨率的功能性磁共振成像结合起来(Lei, Qiu, Xu, &Yao, 2010; Lei, Wang, Yuan, & Mantini, 2014), 通过整合不同脑成像技术所具有的优势, 更精确地揭示脑性别差异。
李雨, 舒华. (2014). 默认网络的神经机制, 功能假设及临床应用.心理科学进展, 22(2), 234–249.
邹吉林, 王美芳, 曹仁艳, 闫秀梅. (2009). 性别发展的生物学取向研究述评.心理科学进展, 17(5), 973–982.
Allen, J. S., Damasio, H., Grabowski, T. J., Bruss, J., &Zhang, W. (2003). Sexual dimorphism and asymmetries in the gray-white composition of the human cerebrum.Neuroimage, 18(4), 880–894.
Andreason, P. J., Zametkin, A. J., Guo, A. C., Baldwin, P., &Cohen, R. M. (1994). Gender-related differences in regional cerebral glucose metabolism in normal volunteers.Psychiatry Research, 51(2), 175–183.
Arnold, A. P., & Breedlove, S. M. (1985). Organizational and activational effects of sex steroids on brain and behavior:A reanalysis.Hormones and Behavior, 19(4), 469–498.
Arnold, A. P., & Chen, X. Q. (2009). What does the “four core genotypes” mouse model tell us about sex differences in the brain and other tissues?Frontiers in Neuroendocrinology, 30(1), 1–9.
Barry, R. J., Clarke, A. R., McCarthy, R., Selikowitz, M.,Johnstone, S. J., & Rushby, J. A. (2004). Age and gender effects in EEG coherence: I. Developmental trends in normal children.Clinical Neurophysiology, 115(10), 2252–2258.
Biswal, B. B., Mennes, M., Zuo, X. N., Gohel, S., Kelly, C.,Smith, S. M., … Milham, M. P. (2010). Toward discovery science of human brain function.Proceedings of the National Academy of Sciences of the United States of America, 107(10), 4734–4739.
Cahill, L. (2006). Why sex matters for neuroscience.Nature Reviews Neuroscience, 7(6), 477–484.
Chen, X. H., Sachdev, P. S., Wen, W., & Anstey, K. J. (2007).Sex differences in regional gray matter in healthy individuals aged 44–48 years: A voxel-based morphometric study.Neuroimage, 36(3), 691–699.
Chou, K. H., Cheng, Y., Chen, I. Y., Lin, C.P., & Chu, W. C.(2011). Sex-linked white matter microstructure of the social and analytic brain.Neuroimage, 54(1), 725–733.
Cicchetti, D. (2013). Annual research review: Resilient functioning in maltreated children–past, present, and future perspectives.Journal of Child Psychology and Psychiatry,54(4), 402–422.
Cosgrove, K. P., Mazure, C. M., & Staley, J. K. (2007).Evolving knowledge of sex differences in brain structure,function, and chemistry.Biological Psychiatry, 62(8),847–855.
Falkenburg, J., & Tracy, D. K. (2014). Sex and schizophrenia:A review of gender differences.Psychosis, 6(1), 61–69.
Friston, K. (2002). Functional integration and inference in the brain.Progress in Neurobiology, 68(2), 113–143.
George, M. S., Ketter, T. A., Parekh, P. I., Herscovitch, P., &Post, R. M. (1996). Gender differences in regional cerebral blood flow during transient self-induced sadness or happiness.Biological Psychiatry, 40(9), 859–871.
Giedd, J. N., Raznahan, A., Mills, K. L., & Lenroot, R. K.(2012). Review: Magnetic resonance imaging of male/female differences in human adolescent brain anatomy.Biology of Sex Differences, 3(1), 19, doi: 10.1186/2042-6410-3-19
Gong, G., He, Y., & Evans, A. C. (2011). Brain connectivity gender makes a difference.The Neuroscientist, 17(5),575–591.
Gong, G., Rosa-Neto, P., Carbonell, F., Chen, Z. J., He, Y., &Evans, A. C. (2009). Age-and gender-related differences in the cortical anatomical network.The Journal of Neuroscience,29(50), 15684–15693.
Grabowski, T. J., Damasio, H., Eichhorn, G. R., & Tranel, D.(2003). Effects of gender on blood flow correlates of naming concrete entities.Neuroimage, 20(2), 940–954.
Gusnard, D. A., & Raichle, M. E. (2001). Searching for a baseline: Functional imaging and the resting human brain.Nature Reviews Neuroscience, 2(10), 685–694.
Halpern, D. F. (2013).Sex differences in cognitive abilities.Hillsdale, NJ: Erlbaum.
Hamilton, C. (2008).Cognition and sex differences. New York: Palgrave Macmillan.
Hines, M. (2004).Brain gender. New York: Oxford University Press.
Hugdahl, K., Thomsen, T., & Ersland, L. (2006). Sex differences in visuo-spatial processing: An fMRI study of mental rotation.Neuropsychologia, 44(9), 1575–1583.
Im, K., Lee, J. M., Lyttelton, O., Kim, S. H., Evans, A. C., &Kim, S. I. (2008). Brain size and cortical structure in the adult human brain.Cerebral Cortex, 18(9), 2181–2191.
Ingalhalikar, M., Smith, A., Parker, D., Satterthwaite, T. D.,Elliott, M. A., Ruparel, K., … Verma, R. (2014). Sex differences in the structural connectome of the human brain.Proceedings of the National Academy of Sciences of the United States of America, 111(2), 823–828.
Kang, H. J., Kawasawa, Y. I., Cheng, F., Zhu, Y., Xu, X. M., Li,M. F., … Šestan, N. (2011). Spatio-temporal transcriptome of the human brain.Nature, 478(7370), 483–489.
Kawachi, T., Ishii, K., Sakamoto, S., Matsui, M., Mori, T., &Sasaki, M. (2002). Gender differences in cerebral glucose metabolism: A PET study.Journal of the Neurological Sciences, 199(1), 79–83.
Larsen, C. C., Bonde Larsen, K., Bogdanovic, N., Laursen,H., Graem, N., Badsberg Samuelsen, G., & Pakkenberg, B.(2006). Total number of cells in the human newborn telencephalic wall.Neuroscience, 139(3), 999–1003.
Larsen, J. K., Hermans, R. C., & Engels, R. C. (2012). Food intake in response to food-cue exposure. Examining the influence of duration of the cue exposure and trait impulsivity.Appetite, 58(3), 907–913, doi: 10.1016/j.appet.2012.02.004
Lei, X., Ostwald, D., Hu, J. H., Qiu, C., Porcaro, C., Bagshaw, A.P., & Yao, D. Z. (2011). Multimodal functional network connectivity: An EEG-fMRI fusion in network space.PloS One, 6(9), e24642.
Lei, X., Qiu, C., Xu, P., & Yao, D. Z. (2010). A parallel framework for simultaneous EEG/fMRI analysis:Methodology and simulation.Neuroimage, 52(3), 1123–1134.
Lei, X., Wang, Y. L., Yuan, H., & Mantini, D. (2014).Neuronal oscillations and functional interactions between resting state networks.Human Brain Mapping, 35(7),3517–3528.
Leonard, C. M., Towler, S., Welcome, S., Halderman, L. K.,Otto, R., Eckert, M. A., & Chiarello, C. (2008). Size matters: Cerebral volume influences sex differences in neuroanatomy.Cerebral Cortex, 18(12), 2920–2931.
Luders, E., Narr, K. L., Thompson, P. M., Rex, D. E., Woods,R. P., DeLuca, H., … Toga, A. W. (2006). Gender effects on cortical thickness and the influence of scaling.Human Brain Mapping, 27(4), 314–324.
Lü, B., Li, J., He, H. G., Li, M., Zhao, M. C., Ai, L. K., …Wang, Z. C. (2010). Gender consistency and difference in healthy adults revealed by cortical thickness.Neuroimage,53(2), 373–382.
Lüders, E., Steinmetz, H., & Jäncke, L. (2002). Brain size and grey matter volume in the healthy human brain.Neuroreport, 13(17), 2371–2374.
Menzler, K., Belke, M., Wehrmann, E., Krakow, K., Lengler,U., Jansen, A., … Knake, S. (2011). Men and women are different: Diffusion tensor imaging reveals sexual dimorphism in the microstructure of the thalamus, corpus callosum and cingulum.Neuroimage,54(4), 2557–2562.
Mizraji, E., & Lin, J. (2011). Logic in a dynamic brain.Bulletin of Mathematical Biology, 73(2), 373–397.
Murphy, D. G., DeCarli, C., Mclntosh, A. R., Daly, E.,Mentis, M. J., Pietrini, P., …Rapoport, S. I. (1996). Sex differences in human brain morphometry and metabolism:Anin vivoquantitative magnetic resonance imaging and positron emission tomography study on the effect of aging.Archives of General Psychiatry, 53(7), 585–594.
Piefke, M., Weiss, P. H., Markowitsch, H. J., & Fink, G. R.(2005). Gender differences in the functional neuroanatomy of emotional episodic autobiographical memory.Human Brain Mapping, 24(4), 313–324.
Reiman, E. M., Armstrong, S. M., Matt, K. S., & Mattox, J. H.(1996). The application of positron emission tomography to the study of the normal menstrual cycle.Human Reproduction, 11(12), 2799–2805.
Ruigrok, A. N., Salimi-Khorshidi, G., Lai, M. C.,Baron-Cohen, S., Lombardo, M. V., Tait, R. J., & Suckling,J. (2014). A meta-analysis of sex differences in human brain structure.Neuroscience and Biobehavioral Reviews,39, 34–50.
Schmithorst, V. J., & Holland, S. K. (2007). Sex differences in the development of neuroanatomical functional connectivity underlying intelligence found using Bayesian connectivity analysis.Neuroimage, 35(1), 406–419.
Schmithorst, V. J., Holland, S. K., & Dardzinski, B. J. (2008).Developmental differences in white matter architecture between boys and girls.Human Brain Mapping, 29(6),696–710.
Smith, S. M., Fox, P. T., Miller, K. L., Glahn, D. C., Fox, P. M.,Mackay, C. E., … Beckmann, C. F. (2009). Correspondence of the brain's functional architecture during activation and rest.Proceedings of the National Academy of Sciences of the United States of America, 106(31), 13040–13045.
Sowell, E. R., Peterson, B. S., Kan, E., Woods, R. P., Yoshii,J., Bansal, R., … Toga, A. W. (2007). Sex differences in cortical thickness mapped in 176 healthy individuals between 7 and 87 years of age.Cerebral Cortex, 17(7),1550–1560.
Sporns, O. (2011). The human connectome: A complex network.Annals of the New York Academy of Sciences,1224(1), 109–125.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D.,Crivello, F., Etard, O., Delcroix, N., … Joliot, M. (2002).Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain.Neuroimage, 15(1), 273–289.
Wang, L. B., Shen, H., Tang, F., Zang, Y. F., & Hu, D. W.(2012). Combined structural and resting-state functional MRI analysis of sexual dimorphism in the young adult human brain: An MVPA approach.Neuroimage, 61(4), 931–940.
Wang, L., Zhu, C. Z., He, Y., Zhong, Q. H., & Zang, Y. F.(2008). Gender effect on functional networks in resting brain.Medical Imaging and Informatics, 4987, 160–168.
Weissman-Fogel, I., Moayedi, M., Taylor, K. S., Pope, G., &Davis, K. D. (2010). Cognitive and default-mode resting state networks: Do male and female brains “rest” differently?Human Brain Mapping, 31(11), 1713–1726.
Willerman, L. (1979).The psychology of individual and group differences. San Francisco: Freeman.
Wu, K., Taki, Y., Sato, K., Hashizume, H., Sassa, Y., Takeuchi,H., … Fukuda, H. (2013). Topological organization of functional brain networks in healthy children: Differences in relation to age, sex, and intelligence.PloS One, 8(2),e55347.
Xu, J., Kobayashi, S., Yamaguchi, S., Iijima, K. I., Okada, K., &Yamashita, K. (2000). Gender effects on age-related changes in brain structure.American Journal of Neuroradiology,21(1), 112–118.
Yan, C. G., Gong, G. L., Wang, J. H., Wang, D. Y., Liu, D. Q.,Zhu, C. Z., … He, Y. (2011). Sex-and brain size–related small-world structural cortical networks in young adults:A DTI tractography study.Cerebral Cortex, 21(2), 449–458.
Zuo, X. N., Kelly, C., Di Martino, A., Mennes, M.,Margulies, D. S., Bangaru, S., … Milham, M. P. (2010).Growing together and growing apart: Regional and sex differences in the lifespan developmental trajectories of functional homotopy.The Journal of Neuroscience, 30(45),15034–15043.
猜你喜欢
中国医药科学(2022年1期)2022-05-05
中学生数理化·七年级数学人教版(2022年11期)2022-02-22
中华书画家(2021年12期)2022-01-06
数学物理学报(2021年2期)2021-06-09
留学(2019年10期)2019-06-21
发明与创新(2016年38期)2016-08-22
中国继续医学教育(2015年6期)2016-01-07
中外医疗(2015年16期)2016-01-04
医学研究杂志(2015年9期)2015-07-01
中国康复理论与实践(2015年7期)2015-05-09
