
紫外诱变对侧耳菌生长、酶活性及纤维降解的影响
2015-01-23 01:19:50刘延国王盼盼
饲料工业 2015年4期
■刘延国 李 杰 张 琳 栗 敏 王盼盼
(东北农业大学动物营养研究所,黑龙江哈尔滨 150030)
白腐真菌能够分解木质素和秸秆纤维吸引人们进行广泛的研究。张辉等[1]采用白腐真菌处理玉米和油菜秸秆,结果表明在发酵培养20 d时,油菜秸秆的木质素降解率达到58.07%;在发酵35 d时,玉米秸秆的木质素降解率达到33.62%。刘朝贵等[2]采用侧耳菌(平菇)处理稻草秸秆,木质纤维成分比处理前降低11.94%~14.7%。韦丽敏等[3]利用多种菌处理秸秆,结果显示黄孢原毛平革菌和侧耳菌的木质素降解率均为27%左右,但前者纤维素的降解率为42%,而侧耳菌纤维素降解率仅有18%。降低秸秆中木质素含量,而保留纤维素是处理秸秆的目标。以上研究结果表明平菇生长速度较快,可利用多种木质纤维物质生长,且要求的生存条件较低,是目前处理秸秆饲料的主要菌种之一。虽然平菇生长较快,但至少也需要20 d[4],所以提高菌种的生长速度和抗逆性是提高秸秆饲料营养价值的关键。紫外诱变(UV)是最简单和常用的提高菌种生长速度和产酶量的真菌育种技术。王谦等[5]用紫外线照射糙皮侧耳的原生质体筛选出高效分解秸秆的菌株08P217,处理秸秆后木质素和纤维素的降解率均提高0.7倍以上。为此,在北方气候条件下,以生长速度较快的侧耳菌为出发菌株,采用紫外诱变技术,培育生长速度较快和产酶量较高的菌株,以提高处理秸秆的效率,为采用白腐真菌处理秸秆饲料提供理论依据和技术参数。
1 材料与方法
1.1 菌种及试剂
菌种:试验所用平菇购于黑龙江省微生物研究所,经过本实验室筛选获得优良菌种P2。
平板与斜面培养基(PDA):20%土豆汁、2%的葡萄糖、2%琼脂,pH值自然。
初筛平板培养基:1%麦芽糖、2%葡萄糖、0.2%酵母膏、0.2%蛋白胨、0.05%MgSO4·7H2O、0.4%KH2PO4、2%琼脂、0.5 mmol/l愈创木酚。
秸秆粉试管培养基:粉碎玉米秸秆87%(其纤维组成见表1)、小麦麸8%、石灰5%、料水比1∶2、121 ℃灭菌30 min,pH值自然。

表1 玉米秸秆粉培养基纤维组成(%)
1.2 试验方法
1.2.1 菌株的活化与保存
将保存的菌种接种到PDA斜面培养基上,在25℃培养7 d,再转接活化2~3次,于4℃保存备用。
1.2.2 孢子悬液的制备
用钟罩法收集孢子于无菌蒸馏水中,适当稀释,血球板法计数,使孢子浓度为1×108个/ml,于4℃保存备用。
1.2.3 紫外线诱变剂
经过试验确定紫外照射孢子的致死率为70%~80%时为最佳诱变剂量[6],将所储备的孢子连续诱变10批,照射后在红光灯下适当稀释,分别涂布于PDA培养基中,无处理的孢子液作对照,置于25℃培养箱中避光培养。挑选出优先萌发且长势良好的菌株接入斜面培养基。
1.2.4 初选
①通过菌丝生长情况的比较,在斜面培养基上进行初筛,从中选出长速快于对照菌株、长势好、菌丝粗壮洁白的菌株与亲本进行拮抗试验,挑选拮抗程度较大的菌株。
②用打孔器将等量诱变菌株菌块接种于初筛平板培养基上,从中挑选变色圈大的菌株,以出发菌为对照。结合生长和产酶两个因素,表现突出的诱变菌株进入复选。
1.2.5 复选
1.2.5.1 PDA试管菌株长速的比较
将筛出的菌株在培养基上传代3次,取等量菌块接种于斜面PDA试管,以出发菌为对照,每株菌5个重复,25℃下恒温避光培养。待菌丝萌发长齐后划线,以游标卡尺(精确度0.02 mm)测量各菌菌丝的生长速度。
1.2.5.2 酶活性及纤维降解率比较
将筛选出的菌种按试验分组接种到秸秆粉试管,以出发菌为对照,每株菌5个重复,在25℃恒温避光培养,菌丝20 d都长满秸秆粉试管,于此时进行相关的测定。参照巩竞的方法[7],取复筛菌株试管秸秆粉各4 g,打碎,混匀,加双蒸水 40 ml,置于4℃冰箱中浸提过夜;次日,用4层纱布过滤离心,取上清即得粗酶液,4℃低温保藏。其残渣65℃下烘至恒重用于粗纤维成分的测定。
1.3 测定指标及方法
致死率(%)=[对照菌数量(cfu)-处理后菌数量(cfu)]/对照菌数量(cfu)×100;
菌丝生长速度(mm/d)=菌丝长度(mm)/菌丝生长时间(d)。
木质素过氧化酶(LiP)酶活的测定:采用亚甲基蓝法;漆酶(Lac)酶活的测定:采用邻联甲苯胺法;纤维素酶酶活的测定:采用DNS法[7]。
发酵秸秆纤维成分的测定:采用ANKOM220 Fiber Analyzer仪器,按照Van Soest分析法,分析中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)、半纤维素(HCEL)、纤维素(CEL)和木质素(ADL)。
1.4 数据统计
试验数据以“平均值±标准差”表示,采用SPSS 17.0软件中的One-Way ANOVA过程进行单因素方差分析,Duncan's法进行各组间多重比较,以P<0.05作为差异显著性判断标准。
2 结果
2.1 紫外诱变剂量的确定(见图1、图2)
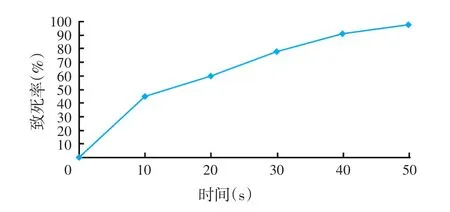
图1 P2紫外诱变孢子致死率
图2 P2紫外诱变30 s萌发菌株
如图1、图2所示,随着照射时间的延长,孢子的萌发数降低,在0~30 s内致死率快速升高,在处理30 s时致死率达78.07%,30 s后上升缓慢,确定以30 s为最佳诱变剂量。
2.2 诱变菌株初选
以30 s为诱变剂量,连续诱变10批,挑出萌发菌株194株,其中26株为正突变,正突变率为13.4%,结合亲本对峙试验(见图3)和初筛平板培养基筛选试验(见表2),筛选出直径较大的5株优良诱变菌株,分别为16、18、20、89、102号,其变色圈为出发菌变色圈直径的1.5倍以上。
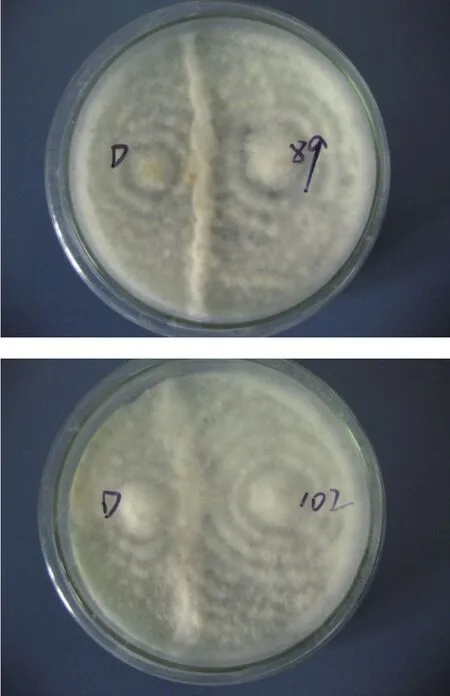
图3 诱变菌株与亲本对峙试验
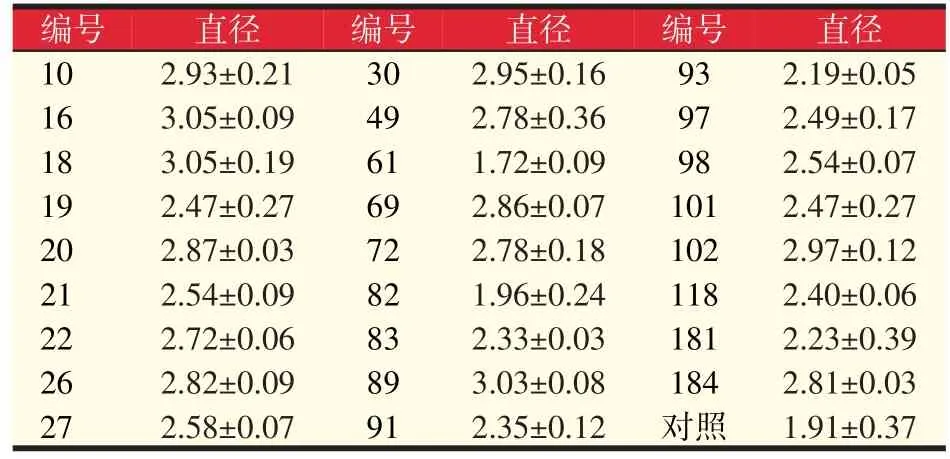
表2 诱变菌种愈创木酚变色圈直径(cm)
2.3 诱变菌株复选
2.3.1 生长速度
诱变菌株的菌丝生长情况见表3。如表3所示,各诱变菌株生长速度均显著快于出发菌,各个菌株生长速度89>20>102>18>16>对照组,其中89、20、102、18号诱变菌株的生长速度极显著地高于对照组(P<0.01),而16号诱变菌株生长速度显著高于对照组(P<0.05)。

表3 诱变菌株的菌丝生长情况(mm/d)
2.3.2 酶活性(见表4)
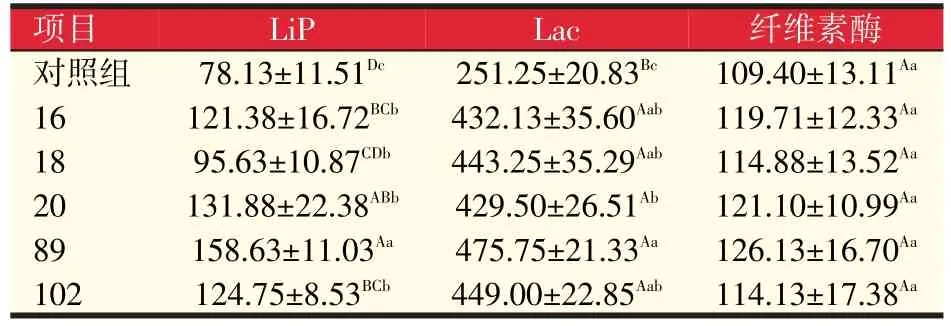
表4 诱变菌株胞外酶活性(U/ml)
如表4所示,与出发菌株对照组相比,诱变菌株木质素过氧化酶(LiP)和漆酶(Lac)活性均不同程度地高于对照组,纤维素酶差异不显著,其中诱变菌株89的酶活性最高,极显著高于对照组(P<0.01),诱变菌株89、20和102号的LiP酶活性分别为对照组2.03、1.69倍和1.60倍,诱变菌株89、102、18号的Lac酶活性分别为对照组1.89、1.79倍和1.76倍。
2.3.3 纤维成分变化(见表5)
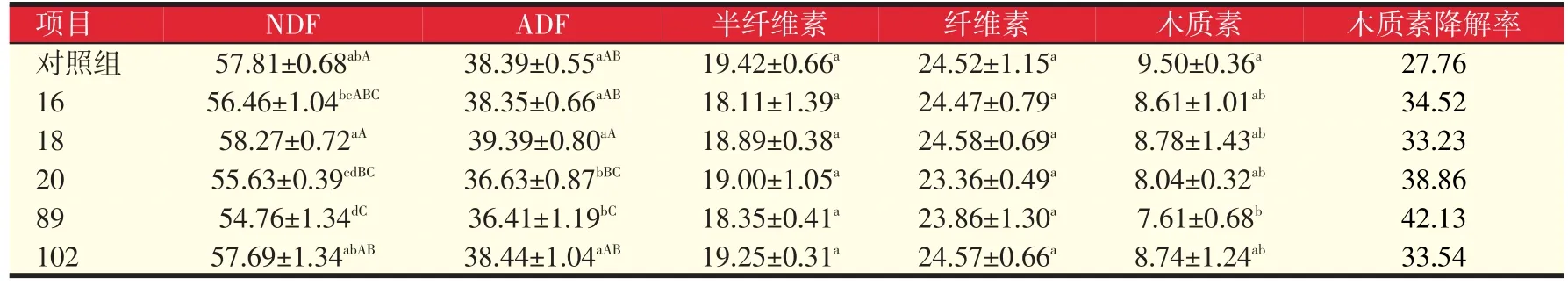
表5 诱变菌株处理秸秆纤维成分变化(%)
如表5所示,各诱变菌株均不同程度降低了部分纤维成分的含量。其中89号菌株纤维含量降低程度最大,20号菌株次之。与对照相比,89号和20号菌株NDF含量分别降低5.28%和3.77%,达到极显著水平(P<0.01)。二者的ADF含量分别降低5.16%(P<0.01)和4.58%(P<0.05)。而木质素含量分别降低了19.89%(P<0.05)和15.37%(P>0.05)。按木质素降解率计算,89号菌株木质素降解率较对照组提高了51.76%。
3 讨论
3.1 紫外诱变及初筛
紫外诱变处理侧耳菌,方法简便,是真菌最常用的育种方法之一[8]。研究表明,适当剂量的紫外线处理能引起DNA的断裂和交联,核酸和蛋白质的交联,胞嘧啶和尿嘧啶的水合作用,形成胸腺嘧啶二聚体。紫外诱变时诱变剂的剂量与灯的功率、照射时间和灯与微生物的距离相关[9]。在本试验中,采用紫外灯照射30 s,照射孢子悬浮液浓度为1×108个/ml左右,致死率为70%~80%。试验结果表明,诱变时间越长,正突变越高,同时负突变率也较高。因此,30 s的紫外照射进行诱变处理有利于提高正诱变率[10]。根据出发菌的生长特性,本研究采用孢子进行诱变,可避免菌落不纯的现象[11]。George等[12]认为,能在以愈创木酚为指示剂的培养基上产生变色圈表明微生物具有木质素降解能力,且变色圈的大小深浅与产酶能力有关。本试验采用了以愈创木酚为唯一碳源的方式,进行木质素降解能力的筛选,诱变菌株和出发菌有明显的拮抗带,表明诱变后为新菌株,且愈创木酚培养基的变色圈明显增大,表明诱变菌株木质素降解酶代谢能力提高。
3.2 酶活性及纤维降解能力
本试验结果表明,通过紫外诱变获得一枚菌株89号,其LiP和Lac活性分别为出发菌的2.03倍和1.89倍,菌丝生长速度提高18.21%,LiP、Lac酶活性高峰出现在处理秸秆的第15~20 d。本试验中纤维素酶活性高峰出现的相对滞后,这正是处理秸秆作为饲料所希望的,分解木质素,而保留纤维素作为家畜的营养。张杰[13]采用射线处理白腐真菌,其中处理效果好的菌株其LiP及Lac活性提高1.4~2倍,而且酶活高峰出现在12~18 d,这与本试验结果接近。荚荣等[14]对白腐真菌F4孢子悬液进行紫外和N+离子注入诱变,获得菌株漆酶活性是出发菌株的16倍。陈敏等[15]对刺芹侧耳(Pleurotus eryngiiGIM 5.280)的原生质体进行紫外和60Coγ双重诱变,获得一株漆酶的高产菌株,与出发菌相比酶活表达量提高54.3%。这些试验结果均表明紫外诱变可以导致菌株产生分解木质素的酶活性显著提高。由秸秆粉木质降解程度可知,木质素降解能力与木质素过氧化酶(LiP)及漆酶(Lac)酶活性密切相关。研究表明,LiP催化氧化β-1非酚型木质素模型化合物为其芳香正离子自由基,使木质素断裂形成3,4-二甲氧基苯乙醇自由基和质子化形式的藜芦醛,并进一步将醛氧化成酸[16],再经过LiP对低分子木质素和单环芳香化合物的开环作用,获得的低分子产物可被微生物降解[17]。
王谦等[5]用紫外线照射糙皮侧耳的原生质体筛选高效分解秸秆的菌株,采用该菌株处理秸秆其木质素和纤维素的降解率分别为出发菌株的1.75倍和1.71倍。张杰[13]采用射线照射白腐真菌处理秸秆,在发酵20~30 d时,出发菌株与诱变菌株发酵秸秆,其酸不溶木质素(ADL)的降解明显提高;在发酵的第30 d,其中的3个诱变菌株处理秸秆使ADL的降解率提高0.88~1.23倍。邱爱莲等[18]采用He-Ne激光对白腐真菌L1原生质体进行诱变,选育出一菌株Lx,其木质素降解率比出发菌株提高50%。本试验结果表明,诱变菌株89号处理秸秆后其木质素含量降低了19.89%,NDF和ADF含量分别降低5.28%和5.16%,木质素降解率较对照组提高了51.76%。其它4株诱变菌株的木质素、NDF和ADF含量变化较小。本结果还表明,不同菌株处理的秸秆纤维含量大小与酶活性高低变化相对应。秸秆的细胞壁结构主要由纤维素、半纤维素和木质素构成,而木质素与半纤维素以其坚固的酯键相结合,并将纤维素包裹在其中。木质素的分解意味着坚固的细胞壁结构被破坏,这样就可以释放动物可以消化利用的纤维素,提高秸秆营养物质的消化利用率。以上研究结果表明,在秸秆发酵的15~20 d,木质素分解酶的活性远高于纤维素酶活性,木质素的降解率也大于纤维素的分解率(由表1及表5计算可知),这有利于降解木质素而保留纤维素的秸秆处理模式。
4 结论
通过紫外诱变育种和筛选试验,综合生长速度、产酶活性及木质素降解能力得到的P89诱变菌株,木质素过氧化酶和漆酶活性分别为出发菌的2.03倍和1.89倍,菌丝生长速度提高18.21%,木质素含量降低19.89%。菌株P89为本研究诱变筛选获得的开发秸秆资源优秀菌株。农业科学,2011,39(8):4598-4599.
猜你喜欢
陕西农业科学(2022年2期)2022-03-28 02:41:54
陕西农业科学(2022年2期)2022-03-28 02:41:54
纺织科技进展(2021年3期)2021-06-09 08:07:14
浙江农业学报(2021年5期)2021-05-31 07:29:08
陶瓷学报(2021年1期)2021-04-13 01:33:02
上海包装(2019年8期)2019-11-11 12:16:14
上海农业学报(2017年4期)2017-04-10 12:40:22
天津造纸(2016年1期)2017-01-15 14:03:29
广东饲料(2016年5期)2016-12-01 03:43:21
中国造纸学报(2015年1期)2015-12-16 19:35:25
