
长白山阔叶红松林中影响乔木幼苗存活的关键因素
2015-01-19 07:03王均伟赵秀海唐景毅
生态学报 2015年19期
肖 翠, 刘 帅, 黄 珍, 樊 莹, 王均伟, 赵秀海,*, 唐景毅
1 北京林业大学省部共建森林培育与保护教育部重点实验室, 北京 100083 2 吉林省白河林业局, 吉林 133600
长白山阔叶红松林中影响乔木幼苗存活的关键因素
肖 翠1, 刘 帅1, 黄 珍1, 樊 莹1, 王均伟1, 赵秀海1,*, 唐景毅2
1 北京林业大学省部共建森林培育与保护教育部重点实验室, 北京 100083 2 吉林省白河林业局, 吉林 133600
应用广义线性混合模型,对长白山阔叶红松林中影响幼苗存活的生物因素和非生物因素进行分析。结果表明:(1)在群落水平上,幼苗存活率和生物因素中同种幼苗邻居显著负相关,说明在阔叶红松林群落中存在负密度制约效应。(2)生物因子和非生物因子对不同年龄阶段的幼苗存活率影响不同。对于1年生的幼苗,幼苗存活率与异种大树邻居呈显著负相关,与同种大树邻居呈显著正相关;对于2—3年生的幼苗,其存活率和同种幼苗邻居、同种大树邻居均呈显著负相关,和非生物因子相关不显著;对于4年生以上的幼苗,其存活率和土壤主成分分析的PC1(低的有机质、速效钾、速效氮等比较贫瘠的土壤)显著负相关。(3)种子的传播方式不同,幼苗存活率的影响因子也不同。对于风传播的物种,存活率与同种幼苗邻居密度显著正相关。对于重力传播的物种,幼苗存活率与土壤PC3(高的全氮和速效氮,含氮较高的土壤)、异种大树邻居、草本密度呈负相关,与林冠开阔度和草本盖度成正相关。(4)对于不同的物种,影响幼苗存活的因素也不同。紫椴的幼苗存活率与土壤PC3、异种大树邻居、草本密度呈显著负相关,与土壤主成分PC2(高的有机质和全氮等养分比较好的土壤)呈显著正相关。
长白山阔叶红松林; 广义线性混合模型; 生物因素; 非生物因素; 幼苗存活
在树木生活史周期中,幼苗阶段是个体生长最为脆弱、对环境变化最为敏感的时期,也是个体数量变化最大的时期[1]。树木幼苗的存活状况间接影响森林群落的物种组成、结构以及稳定性。影响幼苗存活的因素很多,如光照、土壤水分、小尺度环境干扰、坡向、坡度、林冠盖度、枯枝落叶层等各种环境因子[2-3],以及林分条件、物种组成、草食动物啃食、菌类感染、幼苗与母树之间的距离等生物因子[4-6,1]。然而,以往的研究多集中在对单个种群幼苗存活问题的探讨上[7-8],对整个森林群落幼苗存活问题的研究还相对较少。
近年来,随着森林固定监测大样地的发展[9],长期连续定位监测森林群落的动态变化得以实现[10]。幼苗监测作为森林样地监测的一个重要方面,越来越受到重视[11-14]。充分结合和综合利用大样地中的监测数据,能够帮助了解幼苗的动态变化规律以及影响其存活的原因[15-17]。长白山阔叶红松林是典型的温带森林群落,具有丰富的物种多样性,是北半球同纬度地区树种最为丰富的森林类型。针对长白山阔叶红松林的研究一直是我国温带森林生态系统研究的热点[18-19],对群落幼苗存活规律的研究将进一步完善我们对阔叶红松林物种共存的认识。有关阔叶红松林中幼苗的研究已有一些[20-21],而综合探讨生物和非生物等多种因素对幼苗存活影响的研究还比较有限。
综上所述,本文以长白山阔叶红松林永久监测样地为研究平台,以乔木幼苗为研究对象,研究影响幼苗存活的生物和非生物因素,拟回答以下几个问题:(1)哪些因素影响阔叶红松林幼苗的存活?(2)负密度制约现象在温带森林群落中是否普遍存在?(3)生物因素和非生物因素对不同年龄阶段、不同传播方式、不同物种的幼苗存活率影响是否一样?(4)生物因素中的草本植物对幼苗存活是否有显著影响?
1 研究地概况与研究方法
1.1 研究地概况
研究区域位于长白山国家级自然保护区阔叶红松林内。本区地带性气候为温带湿润的季风气候,冬季寒冷干燥,夏季温暖湿润。研究区域立地类型为平缓的坡地,属典型的温带大陆性山地气候[22]。该地区年均气温0.6 ℃,年均降水量707 mm,土壤类型为山地暗棕色森林土[23]。
1.2 研究方法
1.2.1 样地设置
在2007年,参照CTFS (Center for Tropical Forest Science)样地建设标准,选择典型的原始阔叶红松林,应用全站仪建立了面积1 hm2(100 m×100 m)的固定监测样地。样地中心坐标为N42°20.211′,E128°05.705′,海拔为784 m。
1.2.2 乔木调查
记录样地内所有胸径(Diameter of Breast Height)大于等于1 cm的木本植物(包括乔木,灌木),记录其种类,测量其胸径和坐标,并对其定位及挂牌标记[24]。
1.2.3 幼苗及草本植物调查
在样地内均匀设置了50个种子收集器,在每个种子收集器旁边分别设置2个面积1 m×1 m的幼苗调查样方,共计100个幼苗样方,具体位置如图1所示。将样方内胸径小于1 cm的木本植物作为幼苗,调查其种类,并挂牌标记。同时测量样方内草本植物种类、高度、密度和盖度。其中,盖度用投影法测量。即1 m×1 m样方内草本植物投影面积占整个样方面积的比值。每年7—8月复查1次,迄今为止共完成了7次调查。

图1 幼苗样方示意图Fig.1 The sketch mapofseed trap and seedling grid空心方框代表草本和幼苗调查样方,黑色方框代表种子收集器
1.2.4 幼苗年龄的确定
乔木幼苗的年龄的确定是根据长期定位监测和主茎上的芽痕等因素综合判断的。1年生幼苗有子叶,无脱落芽痕;2—3年生幼苗存在1—2圈芽痕,每个芽痕之间有生长量。4年生以上的幼苗存在3圈以上芽痕。
1.2.5 环境因子测量
以样地内20 m×20 m为尺度测量环境因子,包括土壤有机质、全氮、有效氮、有效磷、有效钾、土壤pH值、林冠开阔度。具体调查方法是:在每个20 m×20 m的样方内,随机各取2个500 g土壤表层(0—20 cm)的土壤样品,带回分析。室内分析依据《土壤农业化学分析方法》的步骤进行。两个样方的平均值为每个样方测量结果的最终值。林冠开阔度指林地被树冠垂直投影所覆盖的比例。其测量采用林冠分析仪(WinSCANOPY,Canada)在每个样方中距离地面1m处随机拍摄2张半球状照片,从半球状照片中获得。林冠开阔度也取平均值。在每个样地所处地带地势较为平缓,没有明显的沟、山脊和谷地等特殊地形,故不考虑地形的影响。影响每个20 m×20 m样方内的幼苗存活的环境因子一样。
为了减少多元共线性现象和降低表述土壤因素的变量,参照Bai等对环境因子的处理方法,我们对土壤的有机质、全氮、有效氮、有效磷、有效钾5个代表土壤养分的化学变量进行主成分分析。PC1代表低的有机质、速效钾、速效氮等比较贫瘠的土壤,PC2代表高的有机质和全氮等养分比较好的土壤,PC3代表高的全氮和速效氮,含氮较高的土壤。具体结果见表1。
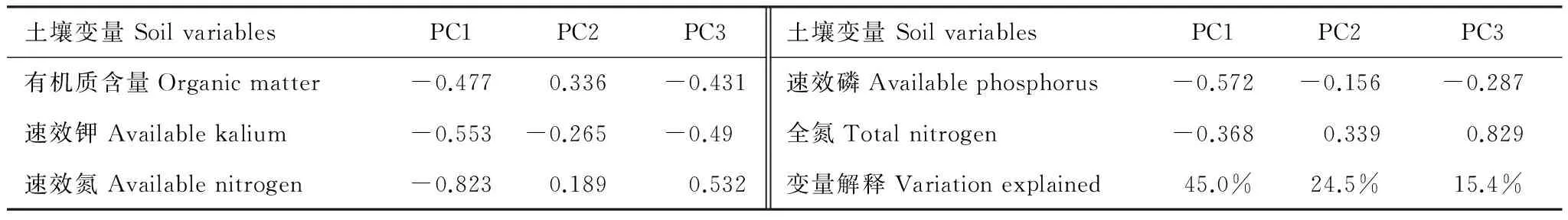
表1 土壤养分的主成分分析
1.2.6 数据分析方法
选取2012年和2013年的幼苗调查数据,应用广义线性混合模型分析幼苗存活与生物因子和非生物因子之间的关联。广义线性混合模型中包含固定效应和随机效应两部分自变量,将生物因子和非生物因子作为固定效应并都经过标准化处理,小样方号和树种被用作随机效应[10]。为了比较生物因子和非生物因子对幼苗存活的重要性,共建立了以下3个备选模型:(1)生物模型:模型中仅包含大树效应、幼苗效应和草本植物效应。大树效应是指以幼苗样方为中心直径10 m范围内的同种大树胸高断面积和异种大树胸高断面积[12]。不同研究选取的尺度不同,大树效应的影响也不同。长白山地区的研究大多数选择20 m范围,这主要是考虑林冠的影响。而本文试验地面积较小,面积仅有1 hm2,同时兼顾大树的地上林冠和地下根系两方面对幼苗的影响,故选取10 m范围[12]。幼苗样方10 m范围内的大树均位于样地内,即不存在边缘效应。幼苗效应是指幼苗所在幼苗调查样方内的同种幼苗密度和异种幼苗密度,草本植物效应是指幼苗调查样方草本植物密度和盖度;(2)非生物模型:模型中仅包含环境因子,即土壤PC1、土壤PC2、土壤PC3、土壤pH值和林冠开阔度。(3)生物+非生物模型。用Akaike′s Information Criterion (AIC)比较模型,不同的模型有不同的AIC值,AIC差值小于2的看作无差别[25]。将幼苗分成以下几个类别进行上述模型检验,(1)群落水平:包括所有乔木幼苗;(2)不同的年龄:1a、2—3a和大于等于4a的幼苗;(3)不同的种子扩散方式:重力传播和风传播。长白山地区靠动物传播种子的物种主要是红松,但因为调查中2012年至2013年存活下来的红松幼苗仅有5株,没有达到统计分析的数量,故本文没有分析动物传播方式;(4)物种水平:2012—2013年调查数据中存活数量最多的前5个树种。为了比较各个参数对幼苗存活的贡献,计算每个参数的优势比率(Odds ratio),优势比率大于1(95%的置信区间)表明与幼苗存活正相关,优势比率小于1(95%的置信区间)表明与幼苗存活负相关[20]。
采用R软件的‘lme4’[26]资源包进行了幼苗存活关键因素的分析。
2 结果
2.1 群落水平的幼苗存活
2012年共调查到1288株乔木幼苗,2013年调查到965棵乔木幼苗,从2012年至2013年仍然存活的幼苗有549棵,占2012年幼苗总量的42.6%。幼苗存活模型中用到的参数见表2。
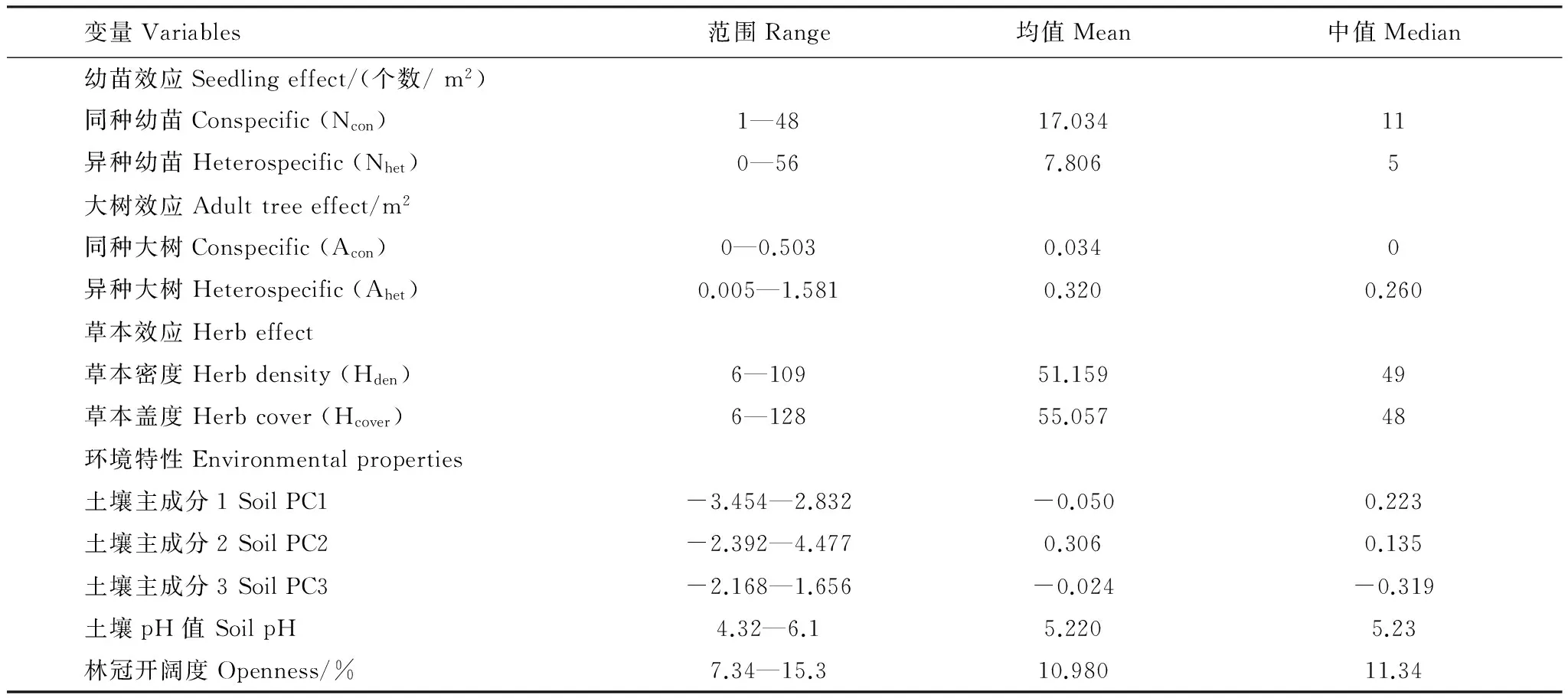
表2 幼苗存活模型中用到的参数
文章中共用了3个模型:生物模型,非生物模型以及生物+非生物模型。通过表3中的ΔAIC值(ΔAIC=AIC-min(AIC))可以判断模型的好坏。ΔAIC值越小的模型越好。
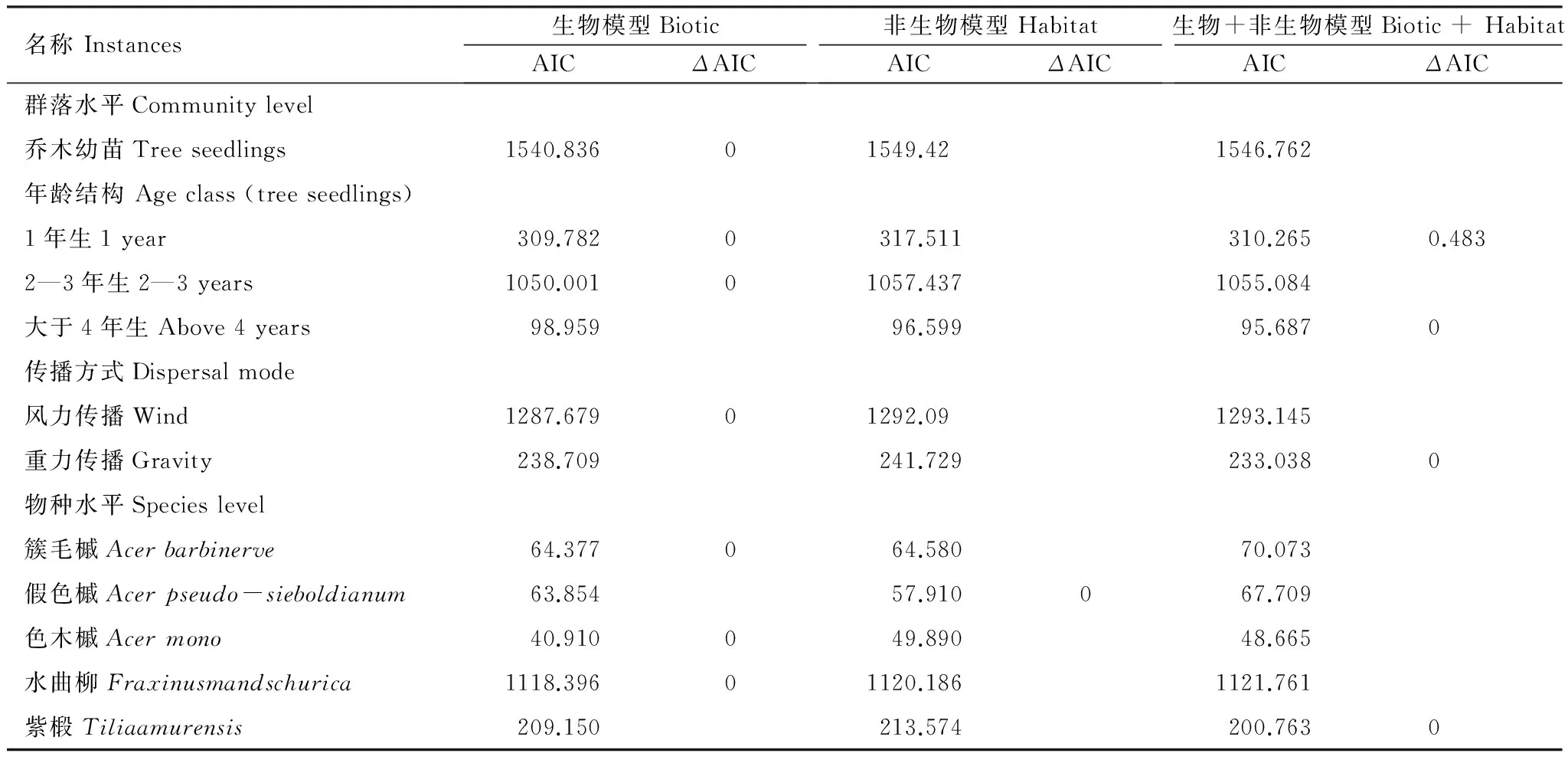
表3 个体水平幼苗存活模型中AIC and ΔAIC值
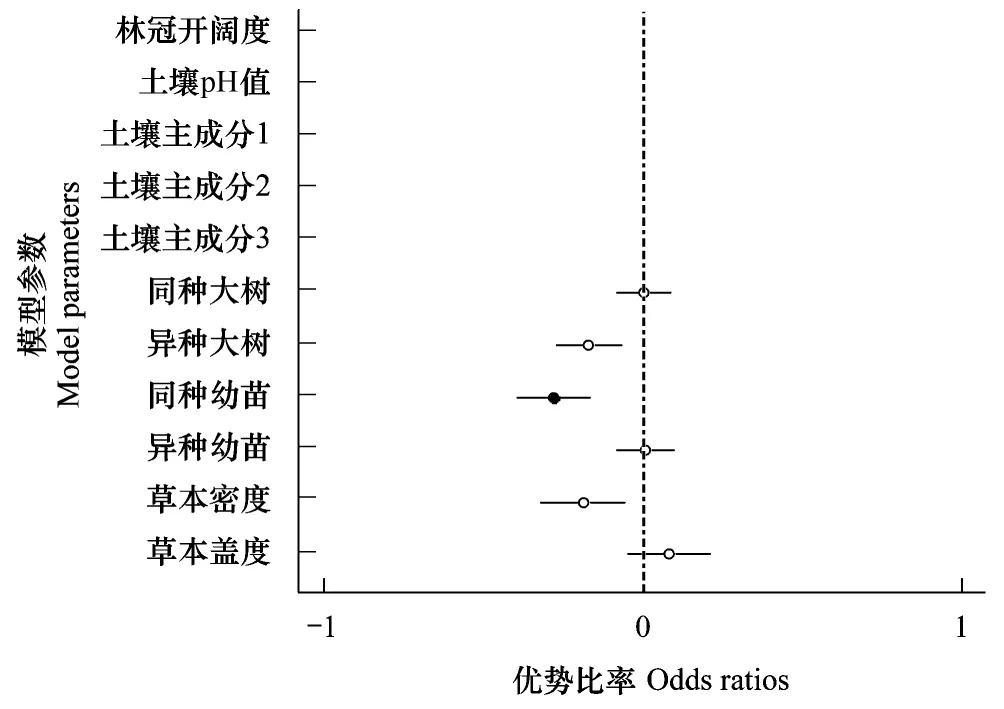
图2 群落水平上幼苗存活的优势比率分析 Fig.2 Odds ratios of a tree seedling survival for community level圆圈代表每个参数的优势比率,实心点表示显著相关,空心点表示相关不显著
群落水平上,最好的模型为生物模型(表3),生物邻居对幼苗的存活率影响更大。由图2的优势比率可以看出,在群落水平上,同种幼苗密度和幼苗存活率呈负相关(odds ratio=-0.28,P=0.013),即同种幼苗密度越大的地方,幼苗存活率越低。
2.2 不同年龄的幼苗存活
不同年龄阶段影响幼苗存活的最优模型不同(图3)。2—3年生幼苗最好的模型是生物模型,1年生幼苗和4年生以上幼苗最优模型是生物+非生物模型。1年生幼苗存活率与同种大树胸高断面积呈显著正相关(odds ratio=0.52,P=0.013),与异种大树胸高断面积呈显著负相关(odds ratio=-0.37,P=0.010)。2—3年生的幼苗与同种大树胸高断面积呈显著负相关(odds ratio=-0.36,P=0.005),与同种幼苗密度呈显著负相关(odds ratio=-0.35,P=0.011)。对于大于等于4a的幼苗,其存活率与土壤主成分分析的PC1呈显著负相关(odds ratio=-1.01,P=0.029)。
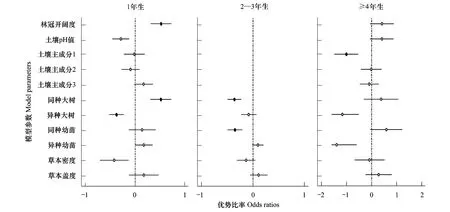
图3 不同年龄结构幼苗存活的优势比率Fig.3 Odds ratios of tree seedling survival for three different age classes圆圈代表每个参数的优势比率,实心点表示显著相关,空心点表示相关不显著
2.3 不同种子传播类型的幼苗存活
表3可以看出,种子风力传播的幼苗存活最优模型是生物模型,而重力传播的幼苗存活的最优模型是生物+非生物模型。
从图4中可以看出,种子风力传播的幼苗存活率与同种幼苗密度呈显著负相关(odds ratio= -0.31,P=0.006)。种子重力传播的幼苗存活率与林冠开阔度呈显著正相关(odds ratio=0.49,P=0.049),与草本盖度(odds ratio=0.89,P=0.038)呈正相关,与土壤PC3呈显著负相关(odds ratio=-0.66,P=0.006),与异种大树胸高断面积呈显著负相关(odds ratio=-0.60,P=0.001),与草本密度呈显著负相关(odds ratio=-0.98,P=0.010)。
2.4 物种水平的幼苗存活
在物种水平上,由表3可以看出,簇毛槭(Acerbarbinerve)、色木槭(Acermono)、水曲柳(Fraxinusmandschurica)的最优模型均为生物模型,假色槭(Acerpseudo-sieboldianum)的最优模型为非生物模型,紫椴(Tiliaamurensis)的最优模型为生物+非生物模型。
而对以上5个样地中最常见的幼苗存活的所有生物和非生物因素进行分析,结果显示:簇毛槭、假色槭、色木槭、水曲柳4个物种与文章中所测的所有生物因素及非生物因素均无显著相关性(图5)。而只有常见种之一的紫椴与土壤主成分PC2、PC3、异种大树胸高断面积、草本密度有相关性(图5)。紫椴幼苗存活率与土壤主成分PC2(odds ratio=0.99,P=0.010)呈正相关。与土壤主成分PC3(odds ratio=-0.78,P=0.003)、异种大树邻居(odds ratio=-0.60,P=0.004)、草本密度(odds ratio=-1.11,P=0.007)均呈负相关。
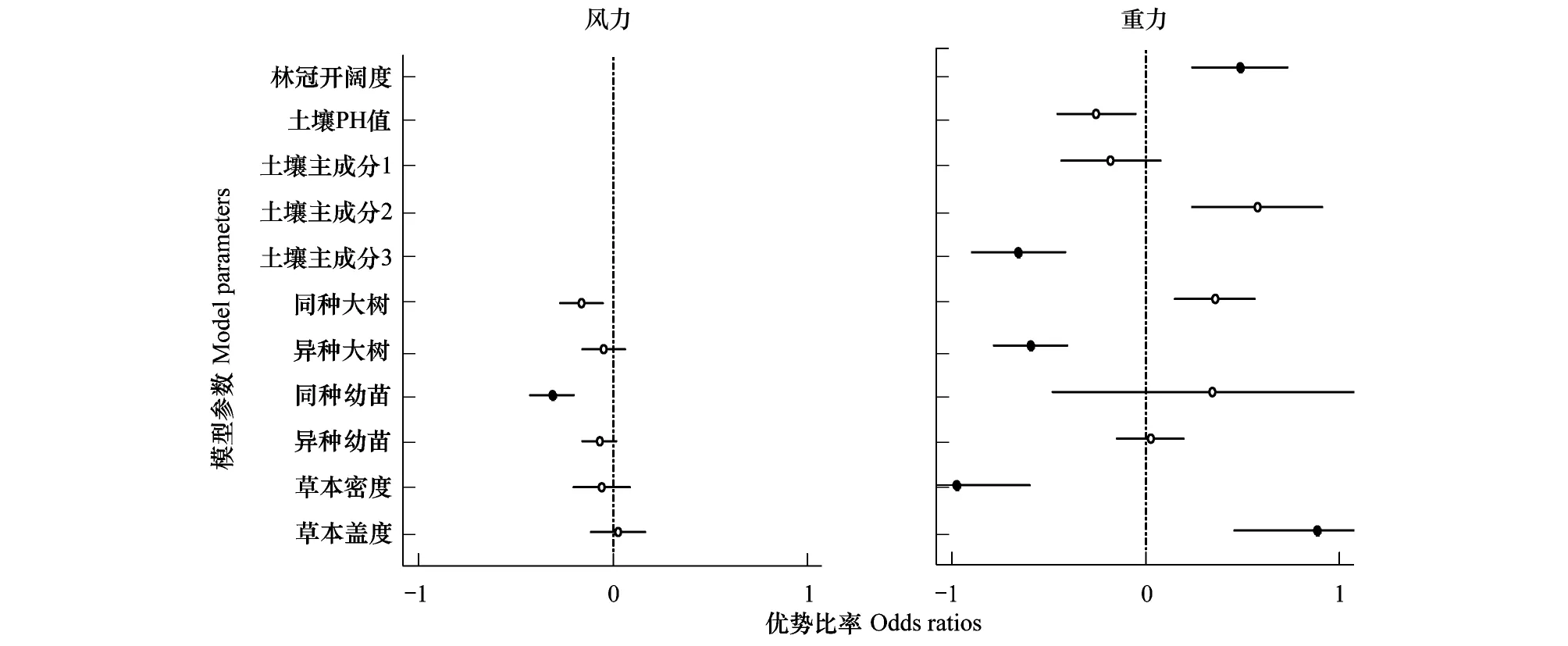
图4 不同种子传播方式幼苗存活的优势比率Fig.4 Odds ratios of tree seedling survival for two dispersalmode groups圆圈代表每个参数的优势比率,实心点表示显著相关,空心点表示相关不显著
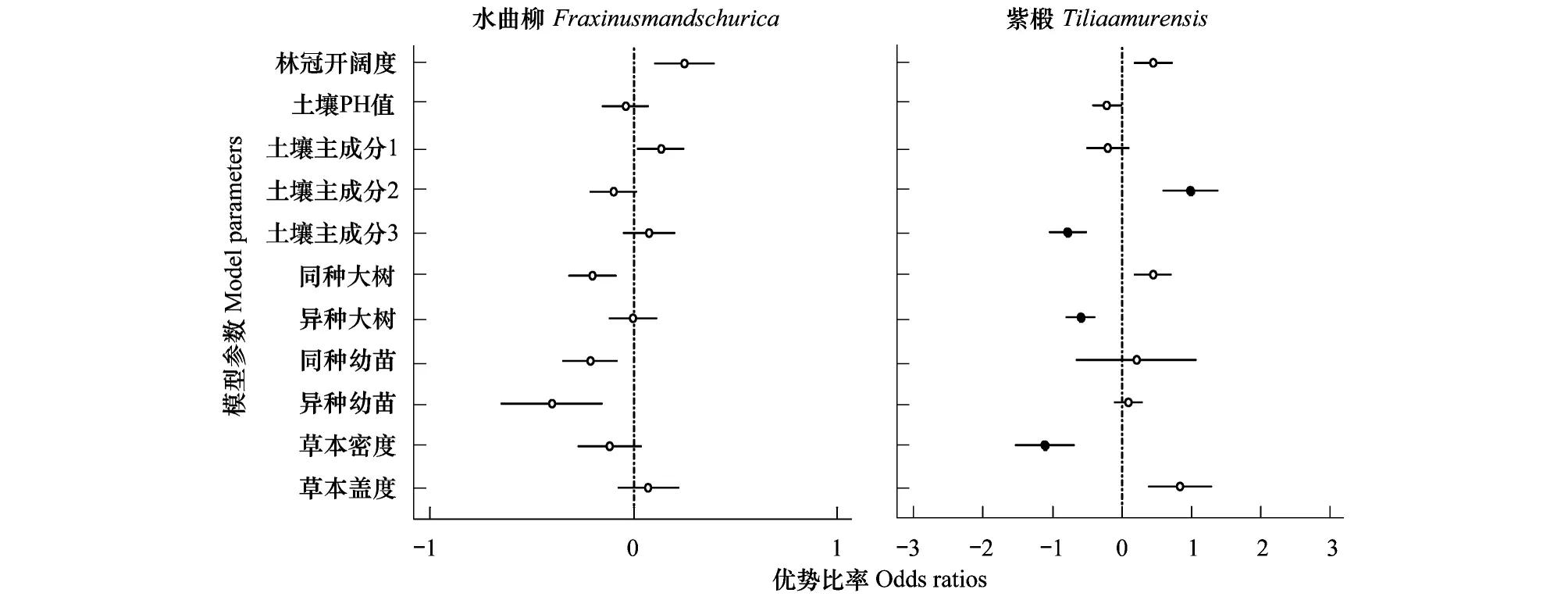
图5 物种水平幼苗存活的优势比率Fig.5 Odds ratios of seedling survival of two common species 圆圈代表每个参数的优势比率,实心点表示显著相关,空心点表示相关不显著
3 结论与讨论
为了探讨影响温带阔叶红松林幼苗存活的因素,从群落水平、不同年龄阶段、不同种子传播方式和物种水平4个方面检验了生物因素和非生物环境对幼苗存活率的影响。
3.1 影响幼苗存活的因素是多样的
幼苗存活受生物邻居和非生物环境共同影响[20],但两者对不同状态下的幼苗影响的程度不同。本文的研究中群落水平上幼苗的存活受生物邻居的影响要大于环境因子的影响(表3)。同种幼苗密度与幼苗存活呈显著的负相关(图2),许多热带、亚热带的研究也得到了相同的结论[27-29]。随着幼苗的生长,影响幼苗存活的因子也在发生变化。4年生以下的幼苗存活受生物因子影响较大(图3),而4年生及以上幼苗存活则受环境因子影响较大(图3)。即随着幼苗年龄的增长,生物邻居对幼苗的影响程度逐渐变小,生境对其生长的影响程度反而增大。这可能由于幼苗对周围环境中与生物邻居之间竞争力增强,对周围环境中真菌感染[30],食草动物和病原菌的抗性增强,而不断生长过程中,需要更多的养分、水分等,反而更依赖于生境环境。种子风力传播的幼苗存活更易受生物因子的影响(图4),而重力传播的物种则受到生物和非生物因素的双重作用(图4),具体原因还有待进一步探讨。不同物种的幼苗存活的影响因子明显不同,紫椴幼苗同时受生物和非生物多种因素显著影响,地上部分和地下部分同时对紫椴的生长起作用[31]。而水曲柳幼苗则基本不受任何因素影响,这与物种本身的生理特性有关,水曲柳有更强的适应能力,生长迅速,成林快,适生于东北东部山地的中生、潮湿和湿生立地[32]。
3.2 负密度制约效应在温带森林中被进一步验证
本研究发现群落水平总体幼苗存活率(图2)、2—3年生幼苗存活率(图3)和种子风力传播幼苗存活率(图4)都与同种幼苗密度或同种大树胸高断面积呈显著负相关,说明在该森林群落中存在负密度制约效应。这与白雪娇等在长白山的研究、祝燕对负密度制约的研究结果相一致[17,20,33]。负密度制约假说主要描述由于资源竞争、有害生物侵害(比如病原微生物、食草动物捕食)等,同种个体之间发生的相互损害行为;它主要强调同种个体之间的相互作用,解释自然群落物种共存的机理。负密度制约假说在多种森林群落中得到验证,Carson等在2008综合分析了53项有关负密度制约的检验,其中81.2%的研究在热带森林,3.8%的研究在亚热带森林,15%的研究在温带森林[34]。尽管生态学家们仍然怀疑负密度制约效应调节群落物种共存的重要性,但是目前的研究还没有发现否定负密度制约假说的充分证据。本文的研究验证了负密度制约效应在温带森林中存在。
3.3 草本效应影响幼苗的存活率
以往对于大样地中草本的研究主要集中在多样性和季节动态、形成格局及影响因素等方面[35-37],也有文章分析草本格局的影响因素中提到乔木层通过影响光照和降水及枯落物的含量而间接影响草本层的多样性格局[37]。而至今在分析影响乔木幼苗存活率的因素中没有提及草本植物的影响。本文将幼苗样方中的草本密度和草本盖度引入幼苗存活率的影响因素中来。分析结果显示,草本植物对某些特定的幼苗存活率有影响,比如草本密度与紫椴幼苗存活率呈负相关(图5),草本密度与重力传播物种的幼苗存活率也存在负相关关系(图4,b)。因为本研究的数据量有限,草本植物对某些幼苗存活率的影响没有检测出来。有待以后加大数据量,更为准确的监测草本植物对幼苗存活率的影响程度。
[1] Wright S J, Muller-Landau H C, Calderon O, Hernandez A. Annual and spatial variation in seedfall and seedling recruitment in a neotropical forest. Ecology, 2005, 86(4): 848-860.
[2] Mason W L, Edwards C, Hale S E. Survival and early seedling growth of conifers with different shade tolerance in a Sitka spruce spacing trial and relationship to understory light climate. Silva Fennica, 2004, 38(4): 357-370.
[3] Poorter L, Rose S A. Light-dependent changes in the relationship between seed mass and seedling traits: a meta-analysis for rain forest tree species. Oecologia, 2005, 142(3): 378-387.
[4] Silvera K, Skillman J B, Dalling J W. Seed germination, seedling growth and habitat partitioning in two morphotypes of the tropical pioneer tree Tremamicrantha in a seasonal forest in Panama. Journal of Tropical Ecology, 2003, 19(1): 27-34.
[5] 彭闪江, 黄忠良, 彭少麟, 欧阳学军, 徐国良. 植物天然更新过程中种子和幼苗死亡的影响因素. 广西植物, 2004, 24(2): 113-121.
[6] Condit R, Ashton P, Bunyavejchewin S, Dattaraja HS, Davies S, Esufali S, Ewango C, Foster R, Gunatilleke I, Gunatilleke CV, Hall P, Harms KE, Hart T, Hernandez C, Hubbell S, Itoh A, Kiratiprayoon S, Lafrankie J, de Lao SL, Makana JR, Noor MN, Kassim AR, Russo S, Sukumar R, Samper C, Suresh HS, Tan S, Thomas S, Valencia R, Vallejo M, Villa G, Zillio T. The importance of demographic niches to tree diversity.Science, 2006, 313(5783):98-101.
[7] 刘庆. 林窗大小和位置对丽江云杉自然更新幼苗存活和生长的影响. 应用与环境生物学报, 2004, 10(3): 281-285.
[8] 盛海燕, 李伟成, 葛滢. 明党参幼苗存活与生长对光照强度的响应. 应用生态学报, 2006, 17(5): 783-788.
[9] 王斌, 黄俞淞, 李先琨, 向悟生, 丁涛, 黄甫昭, 陆树华, 韩文衡, 文淑均, 何兰军. 弄岗北热带喀斯特季节性雨林15ha监测样地的树种组成与空间分布. 生物多样性, 2014, 22(2): 141-156.
[10] 宾粤, 叶万辉, 曹洪麟, 黄忠良, 练琚愉. 鼎湖山南亚热带常绿阔叶林20公顷样地幼苗的分布. 生物多样性, 2011, 19(2): 127-133.
[11] Lambers J H R, Clark J S. Effects of dispersal, shrubs, and density-dependent mortality on seed and seedling distributions in temperate forests. Canadian Journal of Forest Research, 2003, 33(5): 783-795.
[12] Comita L S, Hubbell S P. Local neighborhood and species′shade tolerance influence survival in a diverse seedling bank. Ecology, 2009, 90(2): 328-334.
[13] Queenborough S A, Burslem D F R P, Garwood N C, Valencia R. Taxonomic scale-dependence of habitat niche partitioning and bioticneighbourhood on survival of tropical tree seedlings. Proceedings of theRoyal Society B:Biological Sciences, 2009, 276(1767): 4197-4205.
[14] Streng D R, Glitzenstein J S, Harcombe P A. Woody seedling dynamics in an east texas floodplain forest. Ecology Monograph, 1989, 59: 177-204.
[15] 巩合德, 杨国平, 鲁志云, 刘玉洪, 曹敏. 哀牢山常绿阔叶林乔木树种的幼苗组成及时空分布特征. 生物多样性, 2011, 19(2): 151-157.
[16] 汪殷华, 米湘成, 陈声文, 李铭红, 于明坚. 古田山常绿阔叶林主要树种2002—2007年间更新动态. 生物多样性, 2011, 19(2): 178-189.
[17] Zhang J, Hao Z Q, Sun I F, Song B, Ye J, Li B H, Wang X G. Density dependence on tree survival in an old-growth temperate forest in northeastern China. Annals of Forest Science, 2009, 66(2): 204-204.
[18] Yuan Z Q, Gazol A, Wang X G, Li B H, Lin F, Ye J, Bai X J, Hao Z Q. Scale specific determinants of tree diversity in an old growth temperate forest in China. Basic and Applied Ecology, 2011, 12(6): 488-495.
[19] Wang X G, Wiegand T,Wolf A, Howe R, Davies S J, Hao Z Q. Spatial patterns of tree species richness in two temperate forests. Journal of Ecology, 2011, 99(6): 1382-1393.
[20] Bai X J, Queenborough S A, Wang X G, Zhang J, Li B H,Yuan Z Q, Xing D L,Lin F, Ye J, Hao Z Q. Effects of local biotic neighbors and habitat heterogeneity on tree and shrub seedling survival in an old-growth temperate forest.Oecologia, 2012, 170: 755-765.
[21] 张健, 李步杭, 白雪娇, 原作强, 王绪高, 叶吉, 郝占庆. 长白山阔叶红松林乔木树种幼苗组成及其年际动态. 生物多样性, 2009, 17(4): 385-396.
[22] 李文华, 邓坤枚, 李飞. 长白山主要生态系统生物量生产量的研究. 森林生态系统研究, 1981, (2): 34-50.
[23] 许广山, 丁桂芳, 张玉华. 长白山北坡主要森林土壤有机质及其特征的初步研究. 森林生态系统研究, 1980, (1): 215-220.
[24] Condit R. Tropical Forest Census Plots: Methods and Results from BarroColorado Island, Panama and a Comparison with Other Plots. Berlin:Springer, 1998.
[25] Burnham K P, Anderson D R. Model Selection and Multi-modelInference: APractical Information-theoretic Approach. New York:Springer, 2002.
[26] Bates D, Maechler M, Bolker B. lme4: Linear mixed-effects models using S4 classes. R Package Version: 0. 999375-37edn, 2010.
[27] Metz M R, Sousa W P, Valencia R. Widespread density-dependent seedling mortality promotes species coexistence in a highly diverse Amazonian rain forest. Ecology, 2010, 91(12): 3675-3685.
[28] Queenborough S A,Burslem D F R P, Garwood NC, Valencia R. Neighborhood and community interactions determine the spatial pattern of tropical tree seedling survival. Ecology, 2007, 88(9): 2248-2258.
[29] Webb C O, Gilbert G S, Donoghue M J. Phylodiversity-dependent seedling mortality, size structure, and disease in a bornean rain forest. Ecology, 2006, 87(7): S123-S131.
[30] Masaki T, Nakashizuka T. Seedling demography of Swidacontroversa: effect of light and distance to conspecifics. Ecology, 2002, 83(12): 3497-3507.
[31] 王军邦, 王政权, 胡秉民, 牛铮, 王长耀. 不同栽植方式下紫椴幼苗生物量分配及资源利用分析. 植物生态学报, 2002, 26(6): 677-683.
[32] 荆涛, 马万里, Kujansuu J, 罗菊春, 王广发, 孙波. 水曲柳萌芽更新的研究. 北京林业大学学报, 2002, 24(4): 12-15.
[33] 祝燕, 米湘成, 马克平. 植物群落物种共存机制: 负密度制约假说. 生物多样性, 2009, 17(6): 594-604.
[34] Carson W P, Anderson J, Leigh E G, Schnitzer S A. Challengesassociated with testing and falsifying the Janzen-Connell hypothesis: a review and critique// Carson WP, Stefan AS. Tropical Forest Community Ecology.Oxford, UK: Wiley-Blackwell,2008:210-241.
[35] 夏富才, 潘春芳, 赵秀海, 何海燕, 周海城. 长白山原始阔叶红松林林下草本植物多样性格局及其影响因素. 西北植物学报, 2012, 32(2): 370-376.
[36] 吴刚, 尹若波, 周永斌, 郝占庆, 梁秀英. 长白山红松阔叶林林隙动态变化对早春草本植物的影响. 生态学报, 1999, 19(5): 659-663.
[37] 郝占庆, 赵士洞, 陶大立. 长白山北坡阔叶红松林草本植物物种多样性及其季节动态. 生物多样性, 1994, 2(3): 125-132.
Effects of biotic and abiotic factors on tree seedling survival in a broad-leaved
Korean pine (Pinuskoraiensis) mixed forest on Changbai Mountain
XIAO Cui1, LIU Shuai1, HUANG Zhen1, FAN Ying1, WANG Junwei1, ZHAO Xiuhai1,*, TANG Jingyi2
1TheKeyLaboratoryforSilvicultureandConservationofMinistryofEducation,BeijingForestryUniversity,Beijing100083,China2JilinProvincialBaiheForestryBureau,Jilin133600,China
Seedling recruitment plays a key role in determining species composition and diversity. Exploring patterns of seedling dynamics can therefore provide crucial insights into the mechanisms that affect seedling recruitment. The broad-leaved Korean pine (Pinuskoraiensis) mixed forest is a temperate forest typical of northeast China. In 2007, we established a 1 hm2plot in a broad-leaved Korean pine mixed forest on Changbai Mountain, northeast China. We identified, tagged, and mapped each tree with diameter at breast height ≥1 cm. At the same time, we set up 100 seedling quadrats of 1m × 1m which were censused every July. In total, we collected data on 1288 adult trees and 965 seedlings in the 1 hm2plot between 2012 and 2013, and examined the importance of biotic and abiotic factors on seedling survival using a generalized mixed linear model. Biotic factors included presence of conspecific seedling neighbors and heterospecific seedling neighbors, presence of conspecific adult neighbors and heterospecific adult neighbors, and herb density and herb cover. Abiotic factors included soil organic matter, available soil phosphorus, available soil potassium, available soil nitrogen, total soil nitrogen, and canopy openness. Conspecific neighbors had a significant negative effect on seedling survival at the community level, indicating significant negative density-dependence effects. The effects of biotic and abiotic factors on seedlings differed with seedling age. Conspecific adult neighbors and canopy openness had significant positive effects on survival of, 1-year seedlings while heterospecific adult neighbors had a significant negative effect. In contrast, for 2-to 3-year-old seedlings, conspecific seedling neighbors and conspecific adult neighbors showed a significant negative effect on seedling survival, but abiotic factors had no significant effect. Moreover, for ≥4-year-old seedlings, soil PC axis 1 (associated with low organic matter available K, available N, and total N) showed a significant negative effect on survival. Factors that drove seedling survival varied with species dispersal-mode. For wind-dispersed species, conspecific seedling neighbors had a significant positive effect on seedling survival. For gravity-dispersed species, soil PC axis 3 (associated with high available N and total N, heterospecific adult neighbors, and herb density) had a significant negative effect on survival, whereas canopy openness showed a significant positive effect. The effects of biotic neighbors and habitat differed among species. For instance, soil PC axis 3, heterospecific adult neighbors, and herb density had significant negative effects on seedling survival ofTiliaamurensis, while soil PC axis 2 (associated with high organic matter and total N) showed a significant positive effect. Finally, herbs might influence seedling survival. Both herb density and herb cover impacted seedling survival by modifying local habitat. However, there was no significant effect of herbs on survival due to limited data. We conclude that both local biotic and abiotic factors play important roles in determining seedling recruitment in this temperate forest.
broad-leaved Korean pine (Pinuskoraiensis) mixed forest on Changbai Mountain; generalized linear mixed-effect model; biotic factors; environmental factors; seedling survival
国家“十二五”科技支撑课题(2012BAD22B0203, 2012BAC01B03); 北京市共建项目专项(天然林生物多样性保护技术与杨树抗逆机理研究2012)
2014-03-04; < class="emphasis_bold">网络出版日期:
日期:2014-12-04
10.5846/stxb201403040371
*通讯作者Corresponding author.E-mail: zhaoxh@bjfu.edu.cn
肖翠, 刘帅, 黄珍, 樊莹, 王均伟, 赵秀海, 唐景毅.长白山阔叶红松林中影响乔木幼苗存活的关键因素.生态学报,2015,35(19):6557-6565.
Xiao C, Liu S, Huang Z, Fan Y, Wang J W, Zhao X H, Tang J Y.Effects of biotic and abiotic factors on tree seedling survival in a broad-leaved Korean pine (Pinuskoraiensis) mixed forest on Changbai Mountain.Acta Ecologica Sinica,2015,35(19):6557-6565.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
小哥白尼(野生动物)(2021年9期)2022-01-17
天津医科大学学报(2021年1期)2021-12-05
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
中国林副特产(2020年5期)2020-10-09
林业调查规划(2020年3期)2020-06-03
防护林科技(2017年7期)2017-09-01
中国医学影像技术(2017年11期)2017-01-16
考试周刊(2016年63期)2016-08-15
