
基于CCA的坡面尺度生物结皮空间分布
2015-01-18 07:39卜崇峰杨永胜张晓萍石志华
生态学报 2015年16期
张 朋,卜崇峰,,*,杨永胜,叶 菁,张晓萍,石志华
1 西北农林科技大学水土保持研究所,杨凌 712100
2 中国科学院水利部水土保持研究所,杨凌 712100
3 西北农林科技大学资源环境学院,杨凌 712100
基于CCA的坡面尺度生物结皮空间分布
张 朋1,卜崇峰1,2,*,杨永胜2,叶 菁2,张晓萍2,石志华3
1 西北农林科技大学水土保持研究所,杨凌 712100
2 中国科学院水利部水土保持研究所,杨凌 712100
3 西北农林科技大学资源环境学院,杨凌 712100
生物结皮是干旱半干旱地区普遍存在的活地被物,在该区生态系统中具有重要的生态功能。探讨生物结皮的空间分布规律是科学管理该资源的理论基础。选择黄土高原水蚀风蚀交错区六道沟小流域内的典型坡面,通过全面调查并应用GS+和CANOCO统计软件进行分析,探讨了坡面尺度上生物结皮的空间分布特征及其影响因子。结果表明:(1)生物结皮的分布具有明显的空间分异性。沙土区生物结皮以大面积连续分布为主,平均结皮盖度在30%以上;在黄土区则以零星分布为主,结皮盖度大都在20%以下,主要分布在坡的边缘和末端。而生物结皮厚度和抗剪强度的空间变异不大,说明其主要与生物结皮的发育年限有关。(2)典范对应分析(CCA)表明:生物结皮的空间分布与土壤、植被、地表湿度、坡度坡向等有密切关系。其中,土壤类型和生物结皮的空间分布关系最大,可以解释生物结皮空间变异的20%。其次是植被群落类型和地形湿度指数,沙蒿(ArtemisiadesertorumSpreng. Syst. Veg.)地和小叶杨(PopulussimoniiCarr.)林地是其最理想的生长环境;其他如坡度、坡向、太阳辐射等也都对生物结皮的分布有一定的影响。总体上,生物结皮具有明显的地形、土壤和植被群落的选择性,偏向于生长在较湿润的沙生植被群落当中。
生物结皮(BSCs);典范对应分析(CCA);水蚀风蚀交错区;坡面尺度;空间分布
生物结皮是由微生物、藻类、地衣、苔藓等生物及其代谢产物与土壤颗粒胶结而成的复合体,是干旱半干旱地区广泛存在的地被物[1],在这些地区具有重要的生态功能[2- 4],同时具有良好的水土保持作用。研究其空间分布特征规律和生态功能地位是有效管理和利用该资源的基础。然而,生物结皮的自然发育十分缓慢[5- 6],同时容易受到环境因素和干扰活动的影响[2],因此研究生物结皮的空间分布规律,对于生物结皮资源的野外调查、培育管护,保证其生态功能和水土保持效益的有效发挥是十分必要的。
为此,本研究选取取黄土高原北部水蚀风蚀交错区的六道沟小流域的一处典型坡面,通过全面调查和测算分析,应用GIS、GS+和CANOCO等软件,研究了生物结皮的空间特征,并通过典范对应分析(CCA)探讨土壤类型、植被类型和地形因素等与生物结皮的关系。旨在为野外生物结皮资源空间分布调查和生态功能研究提供借鉴。
1 研究区概况
研究区位于陕西省神木县以西14km处的六道沟小流域(E110°21′—110°23′,N38°46′—N38°51′),海拔1094—1274m,流域面积6.89km2,主沟道南北走向,长4.21km,属窟野河二级支流。地处黄土高原向毛乌素沙地、森林草原向干旱草原的过渡地带,属水蚀作用向风蚀作用过渡的典型水蚀风蚀交错带,年侵蚀模数达10000t/km2[7]。气候属中温带半干旱气候,冬春季干旱少雨多风沙,土壤风蚀严重,夏秋多雨且暴雨频繁,水蚀强烈。年均气温7—9℃,盛行风向为西北风,年平均风速为2.2m/s;年均降雨量400mm,其中6—9月份的降雨量约占全年的70%—80%[8]。流域内的主要植被有沙蒿(ArtemisiadesertorumSpreng)、柠条(CaraganakorshinskiiKom)、长芒草(StipabungeanaTrin)、苜蓿(MedicagosativaL.)、达乌里胡枝子(Lespedezadavurica(Laxm.)Schindl.)、阿尔泰狗娃花(Heteropappushispidus(Thunb.) Less.)等。流域东侧主要为黄土覆盖,占流域面积的86.5%;西侧多为固定沙丘,占流域面积的13.5%[9]。
2 研究方法
2.1 野外调查与制图
本研究在流域内选定了一处撂荒30a以上的典型的梁峁坡面(图1红色廓线所圈区域),于2014年6—10月开展了生物结皮的空间分布的系统调查工作。其中于6、7月份对生物结皮的盖度、厚度和抗剪强度进行了调查。为了精确制图,调查方式采用无样地取样的方式,利用etrex HD型手持GPS对野外每个调查点进行定位,利用方格网法调查该点周围半径10m左右范围内生物结皮的盖度;厚度(mm)用游标卡尺进行原位测定,重复3—5次;抗剪强度(kPa)用袖珍剪力仪(BWT2XZJL)原位测定,重复3—5次;利用ArcGIS 10.1在室内完成生物结皮盖度、厚度和抗剪强度的空间分布制图。于8月中旬到9月上旬对植被类型和植被盖度进行调查,调查的方式仍然采用无样地取样的方式,并利用ArcGIS 10.1在室内完成21种植被的空间分布制图。于10月上中旬在调查区内选取多处典型样地,对样地内的土壤进行采样分析。小心刮去土壤表皮后用环刀取表层0—5cm土层,带回室内105℃下烘24h,测算土壤容重(g/cm3)和质量含水率(%),并利用ArcGIS 10.1采用回归克里格的方法完成了对土壤容重和含水率稀疏数据点的插值和制图;其中,土壤含水率的测定有15d左右的时间梯度,故对土壤表层含水率首先利用除趋势对其进行校正,消除日间蒸发的影响,然后再进行回归克里格插值,得到土壤表层含水率的空间分布图。利用ArcGIS 10.1中的阴影工具结合调查区DEM数据生成10月1日8:00、10:00、12:00、14:00、16:00的太阳高度角的空间分布,来模拟太阳辐射的状况。选择10月1日是因为当地的结皮优势种苔藓的集中繁殖季节在9月到11月期间,考虑到太阳辐射对生物结皮繁殖可能有影响,以10月1日作为这期间的平均太阳辐射具有一定的典型性和代表性。
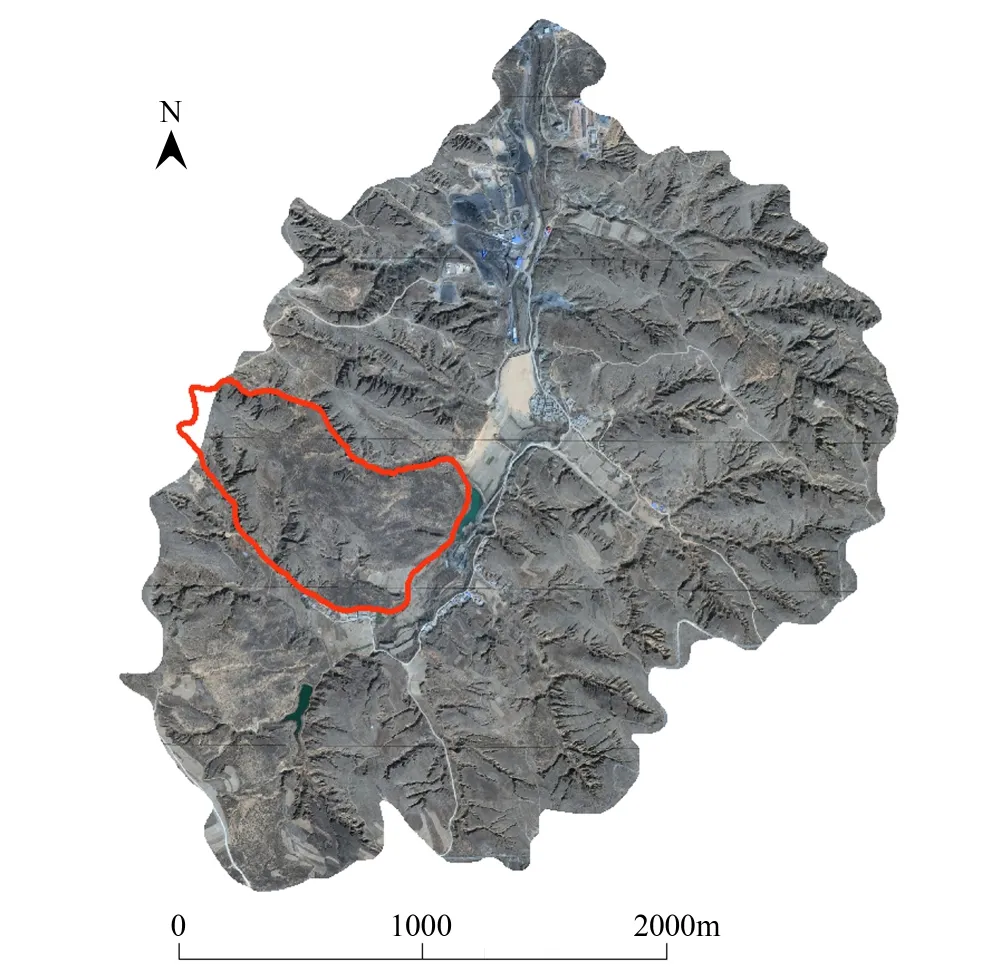
图1 六道沟小流域遥感图Fig.1 Remote sensing image of Liudaogou small watershed
2.2 统计分析
基于六道沟流域的DEM等基础图件,结合上述对生物结皮、植被、土壤的专题图件,利用ArcGIS 10.1软件的空间分析功能,通过重新构建的20m×20m格网对上述底图进行重采样提取,得到1342个样方数据和41个样方指标,形成包括15个环境指标和结皮厚度与抗剪强度构成的环境数据矩阵,以及包括结皮盖度在内的24个物种的盖度构成的物种数据矩阵。利用CANOCO 4.5软件对提取的数据进行CCA分析,得到生物结皮盖度、厚度、抗剪强度和地形、土壤、模拟太阳辐射等环境因子和植被群落之间的相关和对应关系的CCA排序结果和排序图,并结合空间分布图进行解读。
典范对应分析(CCA)是一种非线性多元直接梯度分析方法,它把对应分析与多元回归结合起来,每一步计算结果都与环境因子进行回归,进而详细地研究物种与环境的关系[10]。由于CCA排序是一种基于单峰模型的排序,对于物种的分布有一定的要求,需要先对物种数据进行除趋势对应分析(DCA),排序结果梯度最长的排序轴的长度小于3选择线性模型比较合适,大于4选择单峰模型,介于3—4之间两种模型均可[11]。为此,本文对重采样提取之后得到的样本属性数据首先进行了DCA排序,发现第一排序轴的长度为4.127,适合用基于单峰模型的CCA排序分析,分析结果包括统计数据结果(表2)和CCA二维排序图[12](图5)。
3 结果与分析
3.1 生物结皮的空间分布特征
3.1.1 生物结皮盖度的空间特征
生物结皮的空间分布具有明显的分异性和连续性。图2展示了生物结皮盖度的空间分布,结合土壤类型的空间分布,直观上可以看出,生物结皮主要分布在沙土区,呈现成片的连续分布,统计分析结果(表1)表明:结皮平均盖度在30%以上;而在黄土区,生物结皮则呈现离散的零星分布,盖度大都在20%以下,且主要分布在坡的边缘和末端。地统计软件GS+ 9.0分析结果(图3)表明:沙土区生物结皮的空间分布具有明显的空间自相关性(C/(C0+C)=0.898),在假设各向同性的前提下,对其空间变异的半方差函数用指数模型拟合所得最大空间自相关距离(变程h)为 90—100m。从结皮盖度的空间分布图上看,沙土区生物结皮的空间分布具有明显的发源地(图2结皮蓝色框线区域),大都处在沙土和黄土的交界处的低洼处,其土壤水分和养分状况都极有利于生物结皮的发育。且经过30多年的发育,其在发源地的盖度可以达到90%以上。
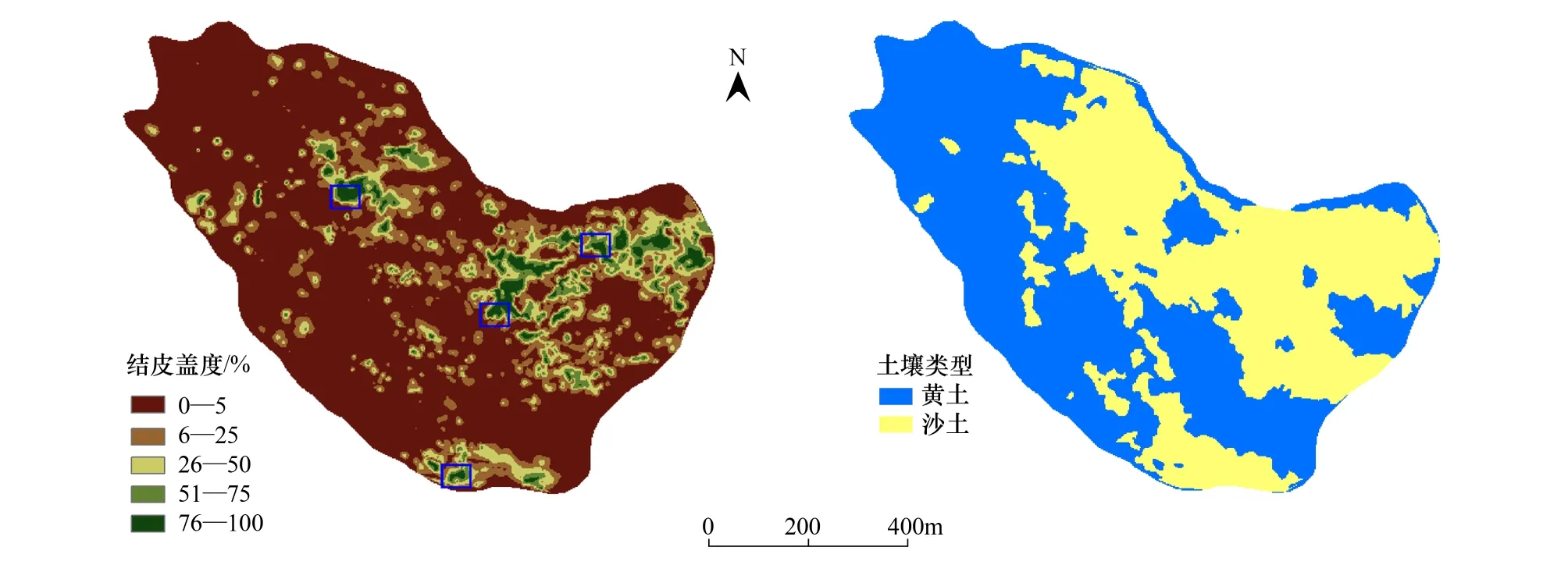
图2 生物结皮盖度和土壤类型的空间分布Fig.2 Distribution of coverage of BSCs and soil type
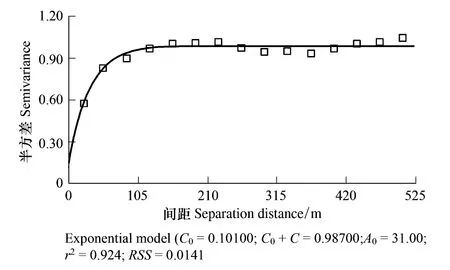
图3 生物结皮空间自相关的半方差函数模型 Fig. 3 Model of semivariance function of BSCs′ spatial autocorrelation
3.1.2 生物结皮厚度和抗剪强度的空间特征
调查分析结果表明:生物结皮的厚度和抗剪强度具有比较明显空间变异性,但总体差异不大。图4展示了生物结皮厚度和抗剪强度的空间分布特征。可以看出生物结皮的厚度大都在12—14mm,抗剪强度大都在40—60kPa;在发源地厚度可以达到20mm以上,抗剪强度可以达到70kPa以上。对沙土区和黄土区的厚度和抗剪强度的统计分析结果(表1)表明:沙土区和黄土区生物结皮的平均厚度分别为13.1 mm和12.8 mm,平均抗剪强度分别为49.6kPa和47.6kPa。t检验结果表明:黄土区生物结皮的厚度和沙土区的差异并不明显(P>0.05),平均的抗剪强度也差异不大(P>0.05)。
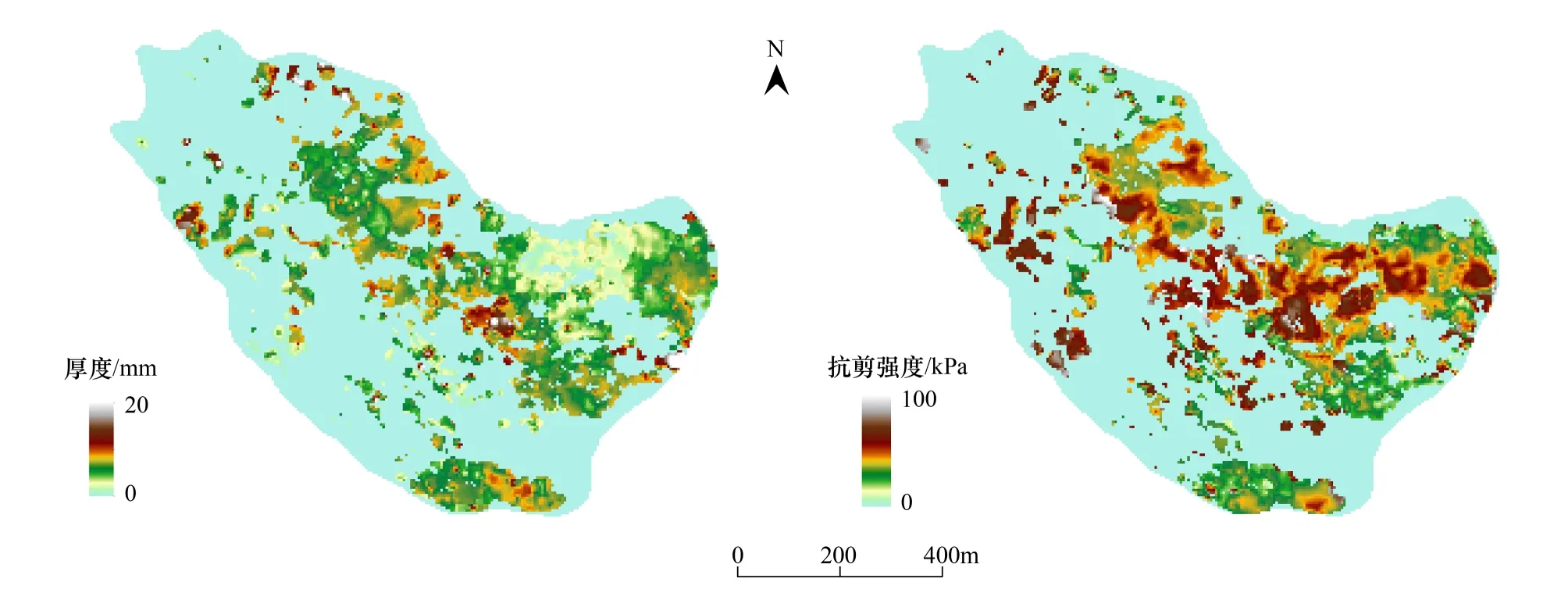
图4 生物结皮厚度、抗剪强度的空间分布Fig.4 Distribution of thickness and shear strength of BSCs
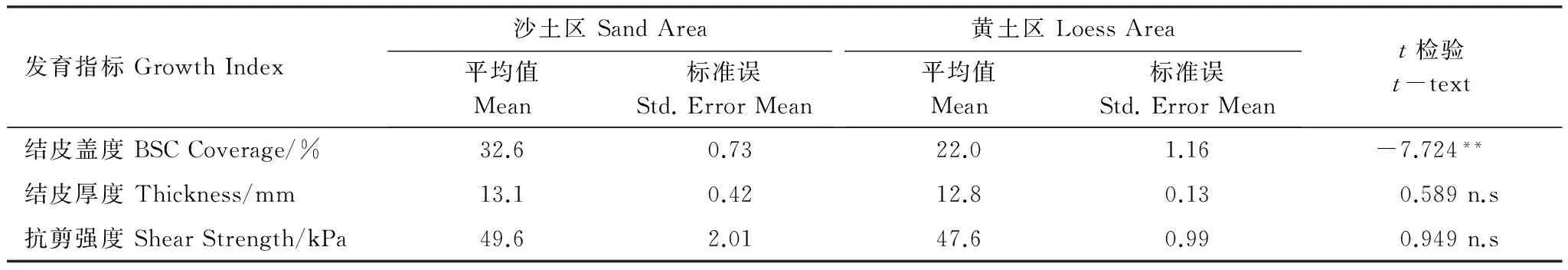
表1 沙土区和黄土区生物结皮发育特征对比结果
3.2 生物结皮的空间分布与环境因子的关系
3.2.1 CCA结果的整体解读
CCA分析结果(表2)显示:包括生物结皮在内的物种的空间分布与第一排序轴(水平轴)的关系最大(r=0.910),可以解释物种与环境关系的60.7%,往后与各其他排序轴的相关性依次减小,4个排序轴总共可以解释物种与环境关系的87.8%,蒙特卡洛置换检验通过(P=0.002),但只能解释物种变异的22.2%,所以对于物种的空间分布的预测能力较差,但对物种和环境因子之间的关系的还原能力很强,即对物种与环境之间的关系的解读是很有意义的。
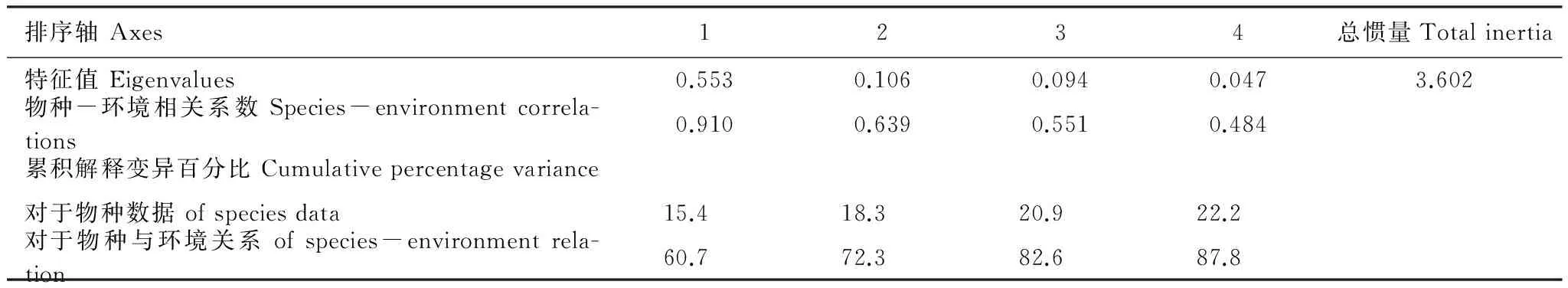
表2 植被与环境的典范对应分析(CCA)分析结果
通过排序图(图5)可以看出:植被群落大概可以分成沙生、黄土生、阴生、阳生四种灌木-草本群落类型和一个小叶杨林地群落类型,主要包括沙蒿群落、沙柳+沙蒿群落、柠条+长芒草群落、猪毛蒿群落、狗娃花+狗尾草群落、硬质早熟禾群落、小叶杨群落等。通过4个排序轴也可以看出环境梯度的变化,第一排序轴(水平轴)与土壤类型、容重关系很大;沿轴的方向,左侧为沙土环境,对应以沙蒿、沙柳、小叶杨为主的沙生植物群落。右侧为黄土环境,对应以柠条、长芒草为主的黄土生植物群落;纵轴与坡度、坡向和太阳辐射关系较大;沿着纵轴方向,上部主要为阴坡陡坡环境,对应早熟禾、硬质早熟禾、铁杆蒿植物群落;下部主要为阳坡缓坡环境,对应狗娃花、狗尾草等植物群落。
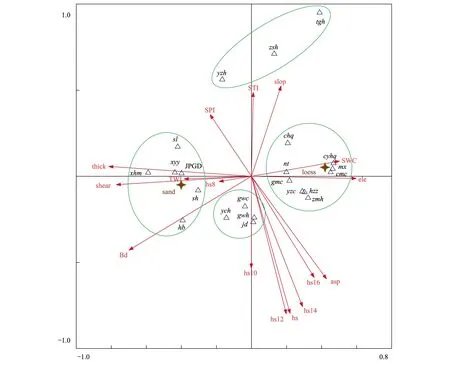
图5 生物结皮、植物群落与环境因子关系的CCA排序图Fig.5 CCA ordination plot of the relationship of BSCs, vegetation communities and environment factors图中箭头代表环境因子,ele:海拔;SWC:土壤含水率;asp:坡向;slop:坡度;Bd土壤容重;hs8—hs12:白天8:00—12:00期间太阳高度角;TWI:地形湿度指数;SPI:汇流动力指数;STI:沉积物运移指数; JPGD:结皮盖度;thick:结皮厚度;shear:结皮抗剪强度;星状质心代表黄土(loess)和沙土(sand)两种土壤类型;sh:沙蒿, tgh:铁杆蒿, ych:茵陈蒿, zmh:猪毛蒿, cl:刺藜, gwh:狗娃花, chq:草木犀状黄芪, cyhq:糙叶黄芪, hb:花棒, hzz:达乌里胡枝子, mx:紫花苜蓿, nt:柠条, jd:棘豆, cmc:长芒草, gmc:冠芒草, gwc:狗尾草, xhm:小画眉, yzc:隐子草, yzh:硬质早熟禾, zsh:早熟禾, sl:沙柳, xyy:小叶杨
3.2.2 生物结皮与地形因子之间的关系
CCA分析的排序图展示了生物结皮盖度、厚度和抗剪强度与坡度、坡向、海拔、地形湿度指数(TWI)、汇流动力指数(SPI)和沉积物运移指数(STI)等地形因子之间的关系。地形湿度指数TWI、汇流动力指数SPI和沉积物运移指数STI是表征特定景观中水分和沉积物运移的有用的综合地形变量[13]。可以看出:生物结皮的盖度、厚度、抗剪强度三者具有良好的一致性,与坡度、坡向、海拔均呈一定程度的负相关;与TWI呈明显的正相关;与SPI存在比较弱的正相关;与STI几乎无关。
此外,由于地形的关系,以平均太阳高度角表征的太阳辐射也对结皮的空间分布有一定影响。从排序图中可以看出:生物结皮的盖度与8:00时的太阳高度角呈较强的正相关,或者说生物结皮比较倾向于分布在10月初8:00时太阳高度角比较大的地方,而与10:00的太阳高度角几乎无关,此后与12:00和14:00、16:00的太阳高度角以及白天平均的太阳高度角hs呈负相关。
3.2.3 生物结皮与土壤因子之间的关系
从CCA排序图中可以看出:代表生物结皮盖度的质心(JPGD)距离沙土质心(Sand)很近,而距离黄土质心(Loess)较远,说明生物结皮集中偏向于在沙土环境中生长,与实际调查过程中的情况相符。仅以土壤类型进入回归分析的结果表明,土壤类型的差异可以解释生物结皮空间变异的20%(R2=0.199)。另外,从排序图中也可以看出,结皮厚度和抗剪强度则与土壤容重呈现出较明显的正相关。
3.2.4 生物结皮与植物群落之间的关系
根据野外调查的结果和CCA排序图来看,生物结皮主要分布在以沙蒿、硬质早熟禾、小画眉为主的沙生阴生草本植物群落和以小叶杨为优势种和建群种的林地群落中,其盖度一般都在30%—40%以上。
对于生物结皮与植被盖度之间的关系,野外调查发现,生物结皮的盖度与沙蒿的盖度有明显的同步趋势,与柠条和长芒草的盖度呈相反趋势。在除去与结皮盖度关系最密切的土壤类型因素之后,通过对物种数据进行偏相关分析发现:结皮盖度与各植物种盖度的相关性均未达到统计学上的显著性,但仍能在一定程度上展示出生物结皮盖度与植被盖度的相关关系,在统计学上属于弱相关。其中与沙蒿、小叶杨、刺藜、小画眉呈一定的正相关关系(相关系数分别为:0.19、0.17、0.25、0.22),与柠条、长芒草、狗娃花呈一定的负相关(相关系数分别为:-0.24、-0.13、-0.13)。
此外,群落的类型和物种组成也对其无机环境类型有代表和指示作用,群落中的某些微小种和偶见种也不能忽略。例如,本研究发现冠芒草极明显地偏向于生长在黄土区内比较湿润的片沙部位,而且基本上有冠芒草的地方往往有发育很好的生物结皮。
4 讨论
4.1 生物结皮厚度与抗剪强度的低空间变异原因探讨
生物结皮的厚度和抗剪强度存在一定的空间变异,但整体差异不大。原因可能来自3个方面:(1)生物结皮的发育厚度可能最主要的还是受发育年限的影响[14],本研究所选调查区撂荒年限都在30a以上,各部分的生物结皮都经过了充分发育,理论上都应该达到了稳定的发育厚度,导致不同部位的生物结皮差异不大;(2)测量中带来的不可避免的误差,由于生物结皮的厚度是生物结皮与下层土壤自然分离情况下的厚度[15],然而实际中不可能保证生物结皮与下层土壤的绝对分离,而且分离面也不可能绝对坚硬平整,而生物结皮厚度本身的变化范围就很小,所以采样测量误差对结皮厚度的影响是比较大且难以控制的;(3)生物结皮厚度的局部变异本身就比较大,甚至会呈现出纯块金效应。生物结皮的抗剪强度同时也会受到多种实际情况的影响,尽管本研究尽量选取那些比较完整而典型的结皮进行测量,但把袖珍剪力仪压入结皮层的过程中很难保证结皮层不被破坏,而且结皮层的干湿程度也会对测量结果造成较大影响。
4.2 地形对生物结皮分布的影响
地形(包括坡度、坡向、海拔等)是影响生物结皮发育和空间分布的一个重要因素,一方面地形会在一定程度上决定与生物结皮发育直接相关的光温水条件的空间分布,另一方面对于生物结皮的繁殖扩张方向也起着一定的导向作用。前者决定了生物结皮的空间变异性,后者决定了生物结皮空间分布的连续性。
本研究中,结皮盖度与坡度、坡向、海拔均呈一定程度的负相关,说明生物结皮倾向于生长在低海拔坡度较缓的阴坡;与TWI呈明显的正相关,说明生物结皮倾向于生长在土壤长期稳定湿度比较高的地方;与SPI存在比较弱的正相关,原因可能是生物结皮的繁殖扩散方向与径流分布存在一定程度的一致性和连续性;与STI几乎无关,可能是因为STI与坡度呈较强的正相关,而生物结皮倾向于生长在坡度较缓的地方,说明SPI比STI更适合预测生物结皮空间分布。
根据结皮盖度与太阳高度角之间的关系,结合其他学者有关研究结论[16- 18],推测可能与生物结皮在一天中的活动时间有关,8:00时的露水凝结比较多,结皮生物开始活跃,并进行光合作用,所以生物结皮更倾向于8:00太阳高度角比较大的地方,以获取更多的太阳能。到10:00时候露水已经蒸发殆尽,结皮活动强度减弱,太阳高度角与结皮分布关系不大了,到了下午来看生物结皮则更倾向于生长在阴坡。
4.3 土壤对生物结皮分布的影响
土壤是影响生物结皮发育和空间分布的最直接、最关键的因子之一,土壤类型甚至在一定程度上决定了调查区域内的群落形态和景观生态格局。虽然沙土是生物结皮发育的良好基质,但并不是越沙越好,纯粹的沙粒上面甚至无法生长结皮。实际调查研究发现沙土与黄土按照一定比例混合效果更佳,从排序图上也可以看出,生物结皮在容重方向上有一个最适的范围,并不是越大越好。
需要指出的是,土壤对生物结皮的影响是多方面的[19],土壤表层含水率对生物结皮的影响比较直接,但含水率也受制于土壤其他方面的理化性质,如容重、孔隙度等;容重只是土壤孔隙状况、机械组成和土壤结构的间接反映,而非直接因素,更为本质的直接因素如土壤PH值、各种土壤养分元素的含量及其状态、土壤结构和孔隙度等对生物结皮发育的影响则需要进一步研究。
4.4 植被群落对生物结皮分布的影响
植物群落与生物结皮之间的关系和相互作用机制比较复杂。有研究表明,生物结皮对维管植物群落的种子库有一定影响[20- 22],如提高种子库的储量、阻碍某些物种种子的萌发等,但也不尽然,和结皮类型以及物种的生活型有关。反过来,维管植物对生物结皮也有一定影响,比如优势种和建群种的冠层结构、种群密度、根系深度、植物的枯落物和分泌物等,都会对生物结皮的发育产生影响。长期的相互作用和自然选择会使得各物种的生态位出现位移和分离,从而形成一种相对稳定的群落有机结构。
需要说明的是,生物结皮与植物群落之间的关系很多情况下未必是直接的或因果的关系,更多的可能是平行的相关关系,生物结皮喜好在沙土环境发育,而沙蒿又是典型的沙生植物,所以两者表现出明显相关性;除去土壤基质影响后的偏相关分析结果也表明,生物结皮的空间分布与植被群落类型之间的弱相关也就证明了这一点。
4.5 存在的问题与展望
典范对应分析(CCA)是20世纪80年代发展起来的一种统计分析方法[10],之后被多次应用到生态学研究当中[23],能够直观而详细地揭示植被群落和环境之间的关系。本研究尝试将CCA方法引入到生物结皮资源分布调查,并根据实际情况,创新地采用了无样地调查和取样的方式,结合GIS、地统计分析等手段,初步探讨了黄土高原北部水蚀风蚀交错区生物结皮的空间分布特征及规律,收到了比较满意的效果。但也发现存在的一些问题,野外调查的数据经过室内转绘成图,基于图数据进行的提取和重采样与实测值相比往往偏低,这是图像插值算法导致的结果。本文大部分分布图都是采用自然近邻法插值制成的,能很好地还原结皮和植被的空间分布,但对其总体盖度却低估了10%左右,因此这种方法与遥感图像的解译结果会存在一定的系统偏差;加密采点精确制成的图像与遥感图像可以高度吻合,但工作量也很大,仅限于小尺度的空间分布调查研究。鉴于此,本研究下一步将尝试利用地面调查结合遥感影像解译,研究流域或区域尺度下的生物结皮空间分布规律及其影响因子,为大尺度下生物结皮的生态功能和防风固沙效应,以及对干旱半干旱地区碳汇方面的影响研究奠定基础[4, 23]。
另外,CCA是以CA/RA为基础的[24],其最大缺点是第二排序轴在许多情况下都是第一轴的二次变形,即存在弓形效应[25]。因此,Braak[26]提出了除趋势典范对应分析(DCCA),克服了弓形效应,其计算结果优于CCA,在弓形效应明显的情况下,可以考虑用DCCA进行分析。
5 结论
(1)生物结皮在小流域坡面尺度上的空间分布具有明显的分异性和一定的连续性。沙土区和黄土区生物结皮的发育和分布规律明显不同,表现为:
1)生物结皮在沙土上广泛发育,分布连续成片,平均盖度在30%以上;在黄土区则呈零星分布,主要分布在黄土坡的边缘和末端,盖度大都在20%以下,规模十分有限;在沙土与黄土交界的低洼处发育最好,盖度可以达到90%以上。
2)生物结皮的厚度和抗剪强度的空间变异相对较小。厚度大都在12—14mm范围内,抗剪强度大都在40—60kPa范围内;在沙土与黄土交界的低洼处厚度可以达到20mm以上,抗剪强度可以达到70kPa以上。
(2)在与生物结皮分布和发育有关的环境因子中,土壤是影响生物结皮空间分布和发育的最主要的因子,土壤类型的基本格局奠定了生物结皮空间分布整体格局的基础,地形和植被因子对生物结皮的影响也十分显著,是导致生物结皮发育和空间分布细部差异的重要原因。具体来说:
1)地形因子中,生物结皮的盖度与坡度、坡向、海拔呈明显的负相关关系,与TWI呈较强的正相关关系,说明生物结皮偏向于生长在湿度较高的低海拔处的背阴缓坡,与SPI呈弱的正相关关系,说明生物结皮的的繁殖扩散与水流方向和分布有一定关系。
2)植被因子中,植物群落类型与生物结皮的分布和发育有明显的相关性,生物结皮偏向于选择在沙蒿、小叶杨、刺藜、小画眉、硬质早熟禾等为优势种的沙生和阴生植物群落中发育。在黄土区的片沙部位,常与冠芒草共生。
本研究在坡面尺度上探讨生物结皮的空间分布与环境因子之间的关系,为更大尺度生物结皮的空间分布资源调查提供了参考和借鉴。
[1] Belnap J. The world at your feet: desert biological soil crusts. Frontiers in Ecology and the Enviroment, 2003, 1(5): 181- 189.
[2] Belnap J, Gardner J S. soil microstructure in soil of the colorado plateau-the role of the cyanobacterium microcoleus-vaginatus. Great Basin Naturalist, 1993, 53(1): 40- 47.
[3] 李新荣, 张景光, 王新平, 刘立超, 肖洪浪. 干旱沙漠区土壤微生物结皮及其对固沙植被影响的研究. 植物学报, 2000, 42(9): 965-970.
[4] 杨晓晖, 张克斌, 赵云杰. 生物土壤结皮——荒漠化地区研究的热点问题. 生态学报, 2001, 21(3): 474- 480.
[5] 闫德仁, 薛英英, 韩凤杰, 赵春光, 郝凯. 沙漠生物土壤结皮国外研究概况. 内蒙古林业科技, 2007, 33(1): 39- 50.
[6] 闰德仁, 薛英英, 赵春光. 沙漠生物结皮国内研究现状. 内蒙古林业科技, 2007, 33(3): 28- 38.
[7] 成向荣,黄明斌,邵明安. 神木水蚀风蚀交错带主要人工植物细根垂直分布研究. 西北植物学报,2007, 27(2): 321- 327.
[8] 李勉, 李占斌, 刘普灵, 崔灵周, 李雅琦. 黄土高原水蚀风蚀交错带土壤侵蚀坡向分异特征. 水土保持学报, 2004, 18(1): 63- 65.
[9] 贾恒义, 雍绍萍, 王富乾. 神木地区的土壤资源 // 中国科学院西北水土保持研究所集刊. 西安: 陕西科学技术出版社, 1993, 18(2): 36- 46.
[10] Braak C J F Ter. Canonical correspondence analysis: A new eigenvector method for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167- 1179.
[11] 王同智, 薛焱, 包玉英, 屈永安, 齐云龙, 王海娟, 董明月. 不同复垦方式煤矿排土场植物群落与土壤因子的关系. 西北植物学报, 2014, 34(3): 587- 594.
[12] Leps J, Smilauer P. Multivariate Analysis of Ecological Data using CANOCO. Cambridge: Cambridge University Press, 2003: 50- 51.
[13] 连纲, 郭旭东, 傅伯杰, 虎陈霞. 黄土高原小流域土壤养分空间变异特征及预测. 生态学报, 2008, 28(3): 946-954.
[14] 赵允格, 许明祥, 王全九, 邵明安. 黄土丘陵区退耕地生物结皮理化性状初报. 应用生态学报, 2006, 17(8): 1429- 1434.
[15] 肖波, 赵允格, 邵明安. 陕北水蚀风蚀交错区两种生物结皮对土壤理化性质的影响. 生态学报, 2007, 27(11): 4662- 4670.
[16] 赵允格, 许明祥, Jayne Belnap. 生物结皮光合作用对光温水的响应及其对结皮空间分布格局的解译. 生态学报, 2010, 30(17): 4668- 4675.
[17] 贾艳, 白学良, 单飞彪, 白少伟, 詹洪瑞. 藓类结皮层人工培养试验和维持机制研究. 中国沙漠, 2012, 32(1): 54- 59.
[18] 张静, 张元明, 周晓兵, 张丙昌, 魏美丽, 生物结皮影响下沙漠土壤表面凝结水的形成与变化特征. 生态学报, 2009, 29(12): 6600- 6608.
[19] 吴玉环, 高谦, 于兴华. 生物土壤结皮的分布影响因子及其监测. 生态学杂志, 2003, 22(3): 38- 42.
[20] 苏延桂, 李新荣, 张景光, 杨丽雯. 生物土壤结皮对土壤种子库的影响. 中国沙漠, 2006, 26(6): 997- 1001.
[21] 苏延桂, 李新荣, 贾荣亮, 潘颜霞. 腾格里沙漠东南缘苔藓结皮对荒漠土壤种子库的影响. 应用生态学报, 2007, 18(3): 504- 508.
[22] 苏延桂, 李新荣, 陈应武, 谭会娟, 贾荣亮. 生物土壤结皮对荒漠土壤种子库和种子萌发的影响. 生态学报, 2007, 27(3): 938-946.
[23] 房世波, 冯凌, 刘华杰, 张新时, 刘建栋. 生物土壤结皮对全球气候变化的响应. 生态学报, 2008, 28(7): 3312- 3321.
[24] 张金屯. 数量生态学. 北京: 科学出版社, 2004: 157- 163.
[25] 贾晓妮, 程积民, 万惠娥. DCA、CCA 和 DCCA 三种排序方法在中国草地植被群落中的应用现状. 中国农学通报, 2007, 23(12): 391- 395.
[26] Braak C J F Ter. The analysis of vegetation-environment relationships by canonical correspondence analysis. Vegetation, 1987, 69(1): 69- 77.
The slope scale distribution regularity of biological soil crusts based on CCA
ZHANG Peng1, BU Chongfeng1,2,*, YANG Yongsheng2, YE Jing2, ZHANG Xiaoping2, SHI Zhihua3
1InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling712100,China2InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistryofWaterResources,Yangling712100,China3CollegeofNaturalResourcesandEnvironment,NorthwestA&FUniversity,Yangling712100,China
Biological soil crusts (BSCs) is a widespread ground living cover in arid and semi-arid areas, which serves an important ecological function in ecosystems of arid regions. To study its distribution regularity is the theoretical basis of scientific management and utilization of this resource. In view of this, this paper chose a typical slope in Liudaogou small watershed of Wind-Water Erosion Crisscross Region of the Loess Plateau, made a comprehensive survey and analyzed by using GS+ and the CANOCO statistical software, discussed the slope scale distribution characteristics of BSCs and its influencing factors. The results showed that: (1) the distribution of BSCs has an obvious spatial differentiation. In sand area, the BSCs mainly distributed widely and continuously, and their mean coverage is generally over 30%. While in loess area, the BSCs mainly shows a sporadic distribution pattern, and their coverage are generally below 20%; However, the spatial variation of the thickness and shear strength of BSCs are not significant, which indicates that it mainly depends on its development years. (2) Canonical correspondence analysis (CCA) showed that the distribution of BSCs has a close relation with soil, vegetation, ground surface humidity, slope and aspect ect. Among them, soil type has the closest relation with the distribution of BSCs, which can explain 20% of spatial variation of BSCs. The next is type of vegetation communities and topographic wetness index, andArtemisiadesertorumSpreng land andPopulussimoniiCarr woodland are its most ideal growth environment. Some other environment factors such as slope, aspect and solar radiation also have some effects on the distribution of BSCs. On the whole, BSCs has an obvious topography, soil and vegetation community selectivity, and tend to develop in humid desert vegetation communities.
biological soil crusts (BSCs); canonical correspondence analysis (CCA); wind-water erosion crisscross region; slope scale; distribution
国家自然科学基金项目(41071192,40701096);中国科学院西部之光人才项目;西北农林科技大学科研专项(2014YQ006)
2014- 12- 18;
2015- 03- 19
10.5846/stxb201412182515
*通讯作者Corresponding author.E-mail: buchongfeng@163.com
张朋,卜崇峰,杨永胜,叶菁,张晓萍,石志华.基于CCA的坡面尺度生物结皮空间分布.生态学报,2015,35(16):5412- 5420.
Zhang P, Bu C F, Yang Y S, Ye J, Zhang X P, Shi Z H.The slope scale distribution regularity of biological soil crusts based on CCA.Acta Ecologica Sinica,2015,35(16):5412- 5420.
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
草业学报(2021年6期)2021-06-28
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持通报(2020年6期)2020-03-10
岩土工程技术(2019年6期)2020-01-06
水土保持研究(2019年6期)2019-10-19
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
西南交通大学学报(2016年4期)2016-06-15
铁道科学与工程学报(2015年5期)2015-12-24
铁道科学与工程学报(2015年4期)2015-12-24
