
一株从片烟中分离的果胶酶菌株培养条件优化
2015-01-18 07:12付相敏王金华包可翔
湖北工业大学学报 2015年5期
何 伟,付相敏,王金华,包可翔
(1福建中烟工业有限责任公司技术中心,福建 厦门361022;2湖北工业大学轻工学部,湖北 武汉430068)
不同等级烟叶中纤维素和果胶等化学成分的含量有所差异[1],其中果胶在各等级烟叶中所占比例约为6%~20%[2]。烟草制品中所含的果胶质,不仅影响烟叶的燃烧性,而且在燃吸过程中容易产生甲醇、甲醛、甲酸和焦油等有害物质[3]。另外,烟叶中所含果胶在自然醇化发酵[4]过程中会产生乙酸,使得烟叶燃吸时有辛辣味和刺激味,产生一种呛咳感[5]。研究表明,烟叶中果胶含量越高,味觉、嗅觉和评吸总分越差[6],且烟叶燃吸产生的甲醛及乙酸会产生呛咳、辛辣等刺激性气味,不利于吸食[7]。
烟叶表面存在大量微生物,这些微生物的活动在烟叶醇化发酵过程中起到了重要作用[8],其实质是微生物发酵产酶降解烟叶中的有害成分。邓国宾等[9]从烟叶中筛选出一株果胶酶高产菌株黑曲霉DPE-005,其产生的果胶酶可使烟叶中果胶质降解高达18.50%;杨慧芳[10]从烟梗中筛选到一株青霉菌JXY-17,将其发酵粗酶液喷洒于烟梗,烟梗中所含果胶质降低了17.53%。果胶酶有降解烟叶中果胶的作用,从而降低了燃吸时产生甲醇等有害成分的含量[11],进而提高了吸味品质。于建军等[12]利用果胶酶处理烟叶,使烟叶在燃吸时降低了有害成分的含量,改善了吸味口感。目前,多采用商品酶制剂降解烟叶中的果胶,但其价格较高,在实际应用中难以控制成本[13]。果胶酶是一种诱导酶,烟叶中的果胶为菌株生产果胶酶提供了诱导环境,有利于产酶。如钱卫等[14]在培养基中添加果胶,果胶酶的产量有明显提高;尤华等[15]发现在果胶诱导物存在的条件下,才能产生果胶酶。
本实验室从烟叶表面筛选到一株果胶酶高产菌,经16SrRNA序列鉴定为克雷白氏杆菌(Klebsiella sp.)。为了进一步提高该菌株果胶酶产量,降低生产成本,本研究选择烟叶为碳源,无机氮化物为氮源,并对培养基的碳源、氮源及初始pH进行了响应面优化,为果胶酶的发酵提供理论依据。
1 材料与方法
1.1 材料
1.1.1 样品 片烟,由福建中烟工业有限责任公司提供。
1.1.2 菌种 克雷白氏杆菌(Klebsiella sp.),由湖北工业大学轻工学部实验室分离及保藏。
1.1.3 培养基 种子培养基(质量分数):蛋白胨1.0%,酵母粉0.5%,NaCl 0.5%,葡萄糖2.0%。发酵培养基质量分数):果胶1.0%,酵母粉0.5%,硫酸铵 0.2%,K2HPO40.2%,NaCl 0.03%,Mg-SO4·7H2O 0.05%,琼脂2.0%,pH 7.0。
1.1.4 试剂及仪器 果胶,美国Sigma-Aldrich公司;D-半乳糖醛酸,97%,美国Sigma-Aldrich公司;DNS试 剂;0.05MpH10的 Na2PO4-NaOH 缓 冲液;可见分光光度计,上海尤尼柯仪器有限公司;恒温培养箱,上海智成分析仪器制造有限公司。
1.2 方法
1.2.1 发酵培养基优化 分别以果胶、橘皮粉、葡萄糖和烟末作为碳源进行发酵产酶,考察不同碳源种类对果胶酶活的影响;将碳源浓度分别设为0、1%、2%、3%、4%、5%进行发酵产酶,考察不同碳源浓度对果胶酶活力的影响;分别以硫酸铵、氯化铵、蛋白胨、胰蛋白胨作为氮源进行发酵产酶,并提取粗酶液,测定其果胶酶活;将氮源浓度分别设为0、0.1%、0.2%、0.3%、0.4%、0.5%进行发酵产酶,考察不同氮源浓度对果胶酶活力的影响;将菌株接种到初始pH 为4、5、6、7、8、9、10的发酵培养基中进行发酵产酶,考察不同初始pH发酵对果胶酶活力的影响。
1.2.2 响应面试验设计 以发酵液粗酶液酶活力为响应值,以单因素实验得到的显著因子起始pH(A)、烟末、硫酸铵为自变量。根据 Central Composite实验确定中心点,各因子实际值及自变量水平见表1。利用Design-Expert8.0软件进行响应面设计及分析。
1.2.3 粗酶液的制备 将筛选得到的产果胶酶菌株Tg1在LB固体培养基上活化2~3代,挑取单菌落接种到液体发酵培养基中,于30℃、220r/min摇床培养72h。发酵液于8 000r/min 4℃下离心10 min,上清液即为粗酶液。
1.2.4 果胶酶活力测定 采用DNS显色法测定果胶酶活性。在刻度为20mL试管中加入1.0mL 0.1%果胶溶液 (0.05MpH10Na2HPO4缓冲液配制),60℃水浴预热5min,加入1.0mL粗酶液,50℃水浴反应60min后,加入2mL DNS溶液,沸水浴10min,取出后浸入自来水中冷却至室温,定容至10mL。以灭活酶液为对照,在540nm下测定吸光度,根据半乳糖醛酸标准曲线计算还原糖含量。
在上述反应条件下,每小时水解果胶底物产生1μmol半乳糖醛酸的酶量为1个酶活性单位,以U/mL表示。
2 结果与分析
2.1 不同碳源对酶活力的影响
实验分别以葡萄糖、烟末、果胶和橘皮粉为碳源,考察不同碳源对酶活力的影响,如图1所示,果胶酶活力由高到低依次为烟末、果胶、橘皮粉和葡萄糖,对应果胶酶活力分别为10.20、9.56、9.19、7.53 U/mL。由于菌种来源于烟末,烟末中所含果胶可以诱导果胶酶的合成,因此所产酶活力最高;葡萄糖作为碳源可以促进菌体生长,但无法诱导果胶酶的合成;以果胶为碳源有利于诱导果胶酶的合成,但营养单一,不利于菌体生长。橘皮粉含有一定量的果胶,但由于该菌株来源于烟末,不能较好地利用橘皮粉所含营养成分。
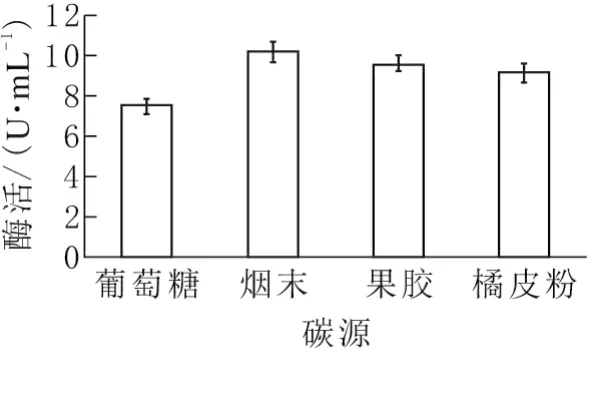
图1 不同碳源对发酵产酶的影响
2.2 碳源浓度对酶活力的影响
分别以碳源浓度0、1%、2%、3%、4%、5%、6%进行实验,考察碳源浓度对酶活力的影响。如图2所示,果胶酶活力随烟末浓度的增大呈先上升后下降的趋势。当烟末浓度为3%时果胶酶活力最高,为10.20U/mL。烟末浓度继续增加,酶活反而呈下降趋势,说明过高的烟末浓度对产果胶酶不利。
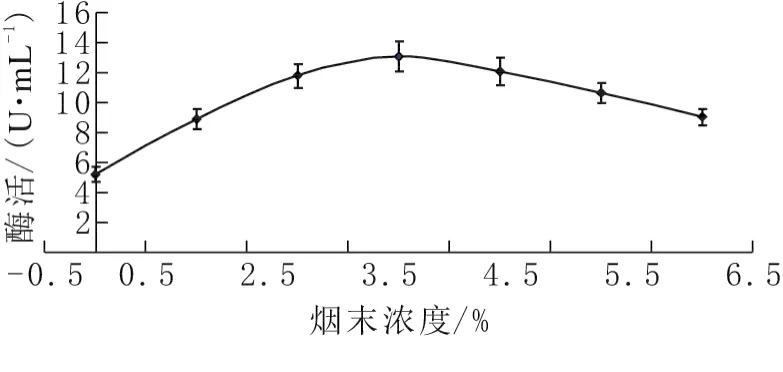
图2 碳源浓度对发酵产酶的影响
2.3 不同氮源对酶活力的影响
以3%烟末作为碳源,氯化铵、硫酸铵、蛋白胨和胰蛋白胨为氮源,考察不同氮源对酶活力的影响。如图3所示,果胶酶活力由高到低依次为硫酸铵、胰蛋白胨、蛋白胨和氯化铵,对应果胶酶活力分别为9.92、8.80、6.32、6.36U/mL,可知硫酸铵和胰蛋白胨作为氮源时效果较好,胰蛋白胨中含有多种有机成分供菌体生长及产酶,而硫酸铵相对廉价,且果胶酶活最高,因此选用硫酸铵作为氮源进行后续实验。
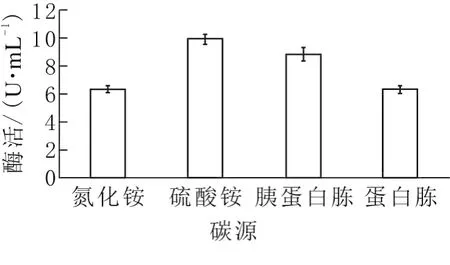
图3 不同氮源对发酵产酶的影响
2.4 氮源浓度对酶活力的影响
分别以氮源浓度0、0.1%、0.2%、0.3%、0.4%、0.5%、0.6%进行实验,考察氮源浓度对酶活力的影响。如图4所示,果胶酶活力随烟末浓度的增大呈先上升后下降的趋势。硫酸铵浓度过低或过高,则碳氮比过高或过低。不能为菌体生长和产酶提供合适的碳氮比;当浓度为0.3%时,果胶酶活力最高,为14.88U/mL,因此选取该浓度进行后续实验。
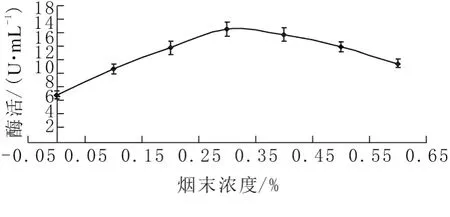
图4 氮源浓度对发酵产酶的影响
2.5 不同起始pH对酶活的影响
分别以浓度3%烟末、浓度0.3%硫酸铵为碳源和氮源,起始pH 分别为5、6、7、8、9的条件下,考察不同起始pH对果胶酶活力的影响。如图5所示,起始pH<7时,果胶酶活力随pH增加而变大;起始pH>7时,果胶酶活力随pH增加而变小;当起始pH为7时,果胶酶活力最高为8.71U/mL。发酵液起始pH偏酸(<7)或偏碱(>7)对酶活均不利。pH值影响培养基中一些成分的解离度,同时对酶的稳定性有影响。实验表明该菌株适合在中性pH环境下产果胶酶。
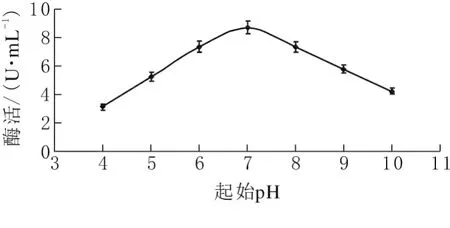
图5 不同起始pH对发酵产酶的影响
2.6 对影响产酶的因素运用RSM法进行优化
用Design-Expert8.0软件设计试验,试验结果见表1。对Central Composite试验设计进行方差分析(表2),利用 Design-Expert 8.0对数据进行三元二次回归方程得到果胶酶活力Y对编码自变量起始pH(A)、烟末(B)、硫酸铵(C)的二次多项回归方程为:酶活力(Y)=-18.54756+6.13519×A+3.73272×B+16.90083×C-0.029722×A×B+0.058333×A×C-1.53056×B×C-0.41439×A2-0.40578×B2-15.88333×C2。由表2中回归模型方差分析(ANOVA)可以看出,FM=26.40,P=0.0001,表明该模型显著;F失拟=1.72,失拟项P=0.3007>0.05,表明失拟不显著;A,B,C,BC,A2,B2,C2项中P <0.05,表明影响显著;回归模型 的 调 整 确 定 系 数 (AdjR-squared)Adj R2=0.954 6,即该模型能解释95.46%响应值的变化,表明该回归模型拟合程度较好,误差小,可用此模型优化果胶酶发酵条件。
表2列出了3个因素在Central Composite试验设计下影响果胶酶活力的回归方程系数及其显著性分析[16]。结果表明,起始pH、烟末、硫酸铵的一次项及其二次项以及烟末和硫酸铵的交互作用对酶活力有显著影响(P<0.05)。其他两两交互效应不显著。
由图6可知,当起始pH值固定时,硫酸铵取某一定值,烟末用量从0到6%范围内增大时曲线较陡,在烟末浓度约为3%果胶酶活达到最大值后开始下降。且硫酸铵与烟末之间交互影响显著。对上述回归模型方程进行典型性分析表明,该模型具有稳定点,即果胶酶活力最大值。
模型给出的最有组合为pH7.20,烟末3.30%,硫酸铵0.35%,在此最优条件下模型预测酶活为15.12U/mL。为了验证模型的准确性,设计三组试验在最优条件下摇瓶发酵产酶,结果平均值15.07 U/mL。实验结果说明回归模型可信度较高,实验值与预测值基本符合,能够较好地预测试验结果。调整系数R2=0.9546
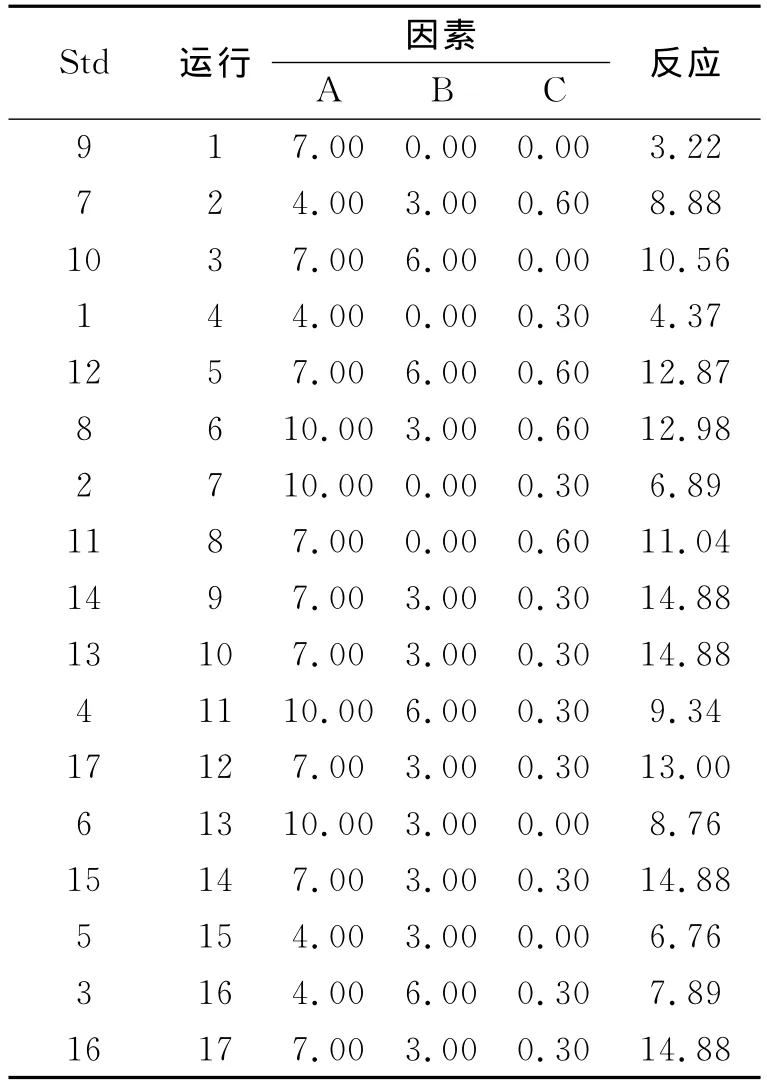
表1 Central Composite试验设计及结果
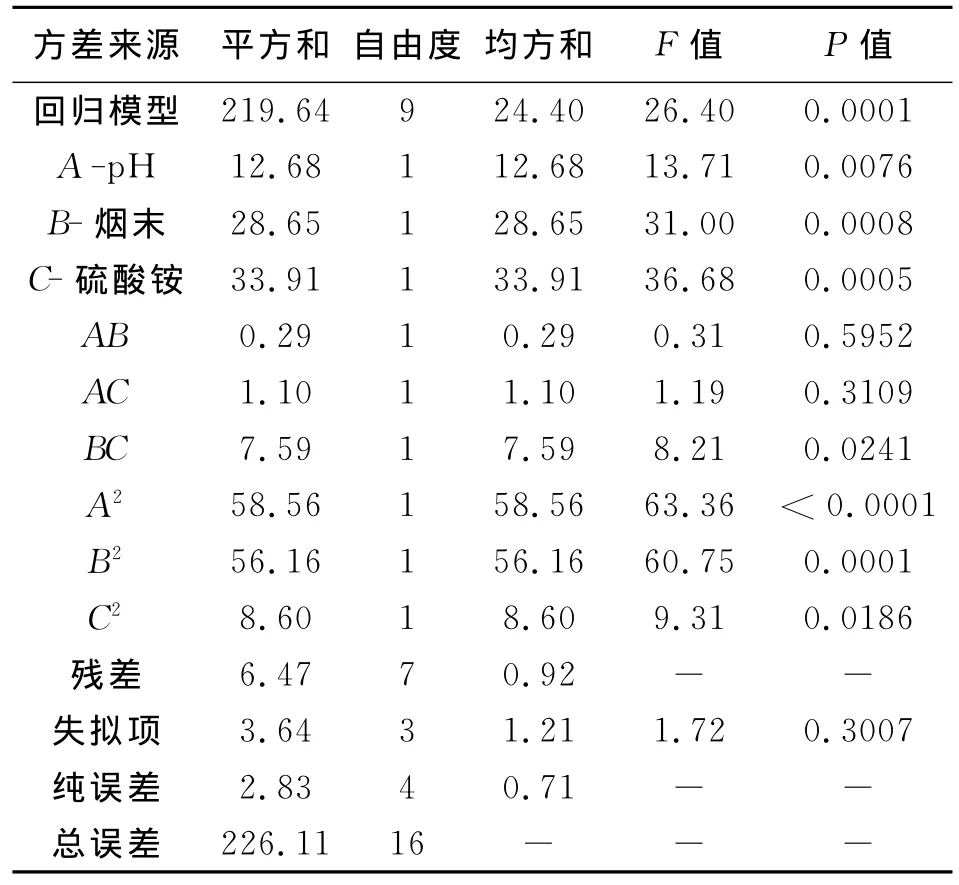
表2 回归方程系数及显著性检验
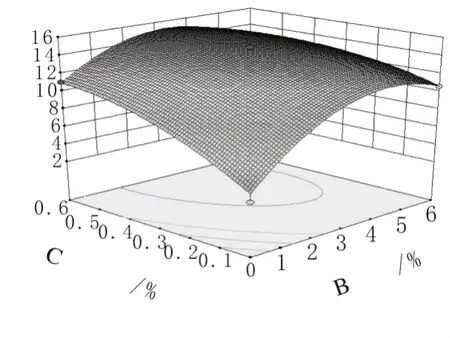
图6 烟末与硫酸铵的交互作用
3 结论
从烟叶表面筛选得到一株果胶酶高产菌株克雷白氏杆菌,最初测得其所产果胶酶活力为10.12U/mL。通过单因素实验确定最佳发酵产酶条件为pH7、碳源为3.00%烟末、氮源为0.30%硫酸铵;在单因素实验的基础上,运用响应面分析对发酵条件进一步优化,结果表明:起始pH7.2,烟末3.30%,硫酸铵0.35%为最佳发酵条件。经摇瓶发酵验证,在该条件下,果胶酶活力为15.07U/mL。与优化前相比,酶活提高约49%。
[1] 闫克玉,闫洪洋,李兴波,等.烤烟烟叶细胞壁物质的对比分析[J].烟草科技,2005(10):6-11.
[2] 李 宁,汪长国,曾代龙,等.蜡样芽孢杆菌 (Bacillus cereus)筛选鉴定及在雪茄烟叶发酵中的应用研究[J].中国烟草学报,2012,18(02):65-69.
[3] 贺兆伟,奚家勤,邓宾玲,等.烟叶中高产果胶酶菌株的筛选及酶学性质[J].烟草科技,2013(05):020.
[4] 颜克亮,武 怡,曾晓鹰,等.基于提质减害的烟叶醇化技术研究进展[J].湖北农业科学,2011,50(03):450-453.
[5] 张 军,曹 茜,李新生,等.再造烟叶原料烟梗中果胶的去除[J].烟草科技,2011(09):5-11.
[6] 陈红丽,任晓红,杨永锋,等.四川烤烟烟叶细胞壁物质含量与外观质量,感官质量的关系[J].烟草科技,2011(01):9-12.
[7] 王志勇,邵 岩,周清明,等.果胶及其对烟草生理和品质性状的影响[J].中国农业科技导报,2013,15(01):130-135.
[8] 赵铭钦,李晓强.烟叶微生物发酵机理及生物制剂应用研究进展[J].甘肃农业大学学报,2008,42(06):84-91.
[9] 邓国宾,李成斌,李雪梅,等.烟叶果胶质分解菌的选育[J].生物技术,2003,13(03):36-38.
[10]杨慧芳.降解烟梗果胶质微生物筛选及产果胶酶的研究[D].江南大学,2012.
[11]代同成,范坚强,郑湖南,等.片烟产果胶酶细菌的鉴定及酶活测定[J].微生物学通报,2011,38(06):816-824.
[12]于建军,马海燕,杨寒文,等.利用果胶酶降解烟叶中果胶的研究[J].江西农业学报,2009,21(03):136-138.
[13]汪天虹,王春卉,高培基.纤维素酶纤维素吸附区的结构与功能[J].生物工程进展,2000,20(02):37-40.
[14]钱 卫,田 敏,李丽莉,等.烤烟叶面微生物5种水解酶的产生,温度稳定性及其在烟叶人工陈化中的应用[J].山东大学学报:理学版,2006,41(05):155-160.
[15]尤 华,陆兆新,冯红霞.曲霉液体发酵产原果胶酶的条件优化研究[J].微生物学通报,2003,30(01):26-30.
[16]刘春泉,刘玉花,李建军,等.响应曲面法优化速冻紫甘薯的烫漂工艺[J].江苏农业学报,2009,25(04):880-884.
猜你喜欢
中国化肥信息(2022年6期)2023-01-06
中国化肥信息(2020年3期)2021-01-07
食品与机械(2020年8期)2020-09-18
中国化肥信息(2018年8期)2018-10-08
中成药(2018年7期)2018-08-04
山东工业技术(2016年15期)2016-12-01
中国酿造(2016年12期)2016-03-01
应用海洋学学报(2015年3期)2015-11-22
中国果菜(2015年2期)2015-03-11
中国酿造(2014年9期)2014-03-11
