
云斑天牛幼虫空间分布格局的地统计学分析
2015-01-03 05:12胡瑜珩武海卫许志春
西北林学院学报 2015年5期
胡瑜珩,武海卫,许志春*
(1.北京林业大学 林木有害生物防治北京市重点实验室,北京100083;2.山东省林业科学研究院,山东 济南250014)
云斑天牛(Batocera horsfieldi)隶属于鞘翅目(Coleoptera)天牛科 (Cerambycidae),是我国重要的蛀干害虫[1]。国外主要在越南、日本、印度、缅甸等国家有分布;我国则已经遍及河北、北京、山东等大部分省份[2]。主要危害杨树(Populus)、白蜡(Fraxinus chinensis)、榆 (Ulmus pumila)、核 桃(Juglans regia)、苹 果 (Malus pumila)、梨(Pyrus)、和板栗(Castanea mollissima)等。其成虫危害新枝皮和嫩叶,幼虫蛀食枝干,造成树木生长势衰退,凋谢乃至死亡。目前,国内外对云斑天牛的研究比较广泛,在生物学、生态学以及药剂防治方面均有报道[3-6]。
了解昆虫的时空动态及种群数量的空间分布格局是阐释其爆发机制并制定相应的虫害防治策略的前提和基础[7]。地统计学自20世纪50年代形成之后,逐步应用于土壤、地理、生物、矿业、农业以及森林等诸多领域中[8-12]。如今,地统计学因为克服了昆虫种群空间格局动态研究中处理多维空间数据的困难性和复杂性以及还能分析较大尺度上的空间结构,为研究昆虫种群分布格局和变迁规律提供了最为有效的途径[13-15]。目前,在我国运用地统计学探究昆虫空间格局的课题已见多例,而利用此方法分析云斑天牛幼虫的空间格局的研究鲜见报道。本文应用地统计学方法对不同受害程度的白蜡林中云斑天牛幼虫种群空间格局进行调查,以期为种群动态监测和防治等提供参考和依据。
1 研究区概况
试验地设在山东省林业科学研究院盐碱地造林试验站,位于渤海湾西南岸、寿光市北部,地理位置为118°40′E、37°11′N,平均海拔35m,属暖温带季风区大陆性气候,年平均气温13℃,年均降水量600 mm,年均蒸发量2 200mm,地下水位2~3m,全年无霜期195d[16]。植被类型包括:以白蜡树、杨树、旱柳为主,还有少量榆树、刺槐、柽柳。
2 材料与方法
2.1 调查方法
2013年6-8月,根据试验地的大小及白蜡树的分布情况,采取逐株取样调查的方法,从白蜡树的基部往上至树梢,每株分别按1m以下、1~2m、2~3m和3m以上4个高度区域划分,统计每个高度区域排粪孔数量,并以此作为密度指标,同时在坐标纸上记录样地中每株白蜡树的相对空间位置。
2.2 分析方法
2.2.1 垂直分布 利用SPSS Statistics 22.0对不同样地内云斑天牛幼虫4个高度区域垂直分布进行单因素方差分析(ANOVA)与多重比较(LSD),判别幼虫垂直分布的差异显著性(α=0.05)。
2.2.2 半变异函数 昆虫种群数量是空间上分布的变量,即区域性变量[7,17-18]。因此,变异函数理论可以运用到云斑天牛幼虫种群数量一定范围内空间分布的相关性分析,从而阐释其爆发机制,并制定相应的虫害防治策略。
半变异函数也称为半方差函数,它是地统计学中研究空间变异性的关键函数,是指区域化变量Z(xi)和Z(xi+h)的增量平方的数学期望,即区域化变量增量的方差,其公式为:
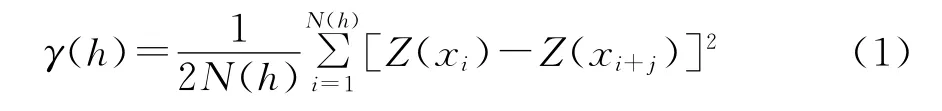
式中:γ(h)为相对于距离h的估计半方差函数值,N(h)是被h分割的数据对(xi,xi+h)的对数,Z(xi)和Z(xi+h)分别是xi和,xi+h处样本的测量值,h是分割两样点的距离[19-22]。
半变异函数是空间变异性的度量,是区域化变量结构性和随机性的定量描述[14],而块金系数、基台值和变程是其重要参数。块金系数是由于存在测量误差或者空间变异,使得2采样点非常接近时半变异函数值不为0而产生的截距。基台值是指随采样距离(h)的变化达到稳定时的函数值,变程指函数值从初始值达到稳定值的采样间隔,揭示空间相关性的作用范围[9]。
2.2.3 变异函数理论模型的拟合 地统计学中变异函数理论模拟昆虫种群空间格局特征常用的模型有表征所研究种群为聚集分布的球型、指数型、高斯型,以及表征种群呈随机分布或均匀分布的直线型。前者指半变异函数值未达到稳定状态之前,空间依赖性的大小随采样距离增大而逐渐降低[8-9];后者表明在抽样尺度下没有空间相关性[23]。具体选择采用哪种理论模型主要依据决定系数R2、残差RSS、变程和块金系数的大小依次判断,从而得到最优拟合模型[20-22]。
2.2.4 空间分布图的生成 克里金(Kriging)法是地统计学的主要内容,可以定量地描述地理空间分布格局。从统计意义上说,是从变量相关性和变异性出发,利用半变异函数与原始数据之间存在的结构性特征,在有限区域内对区域化变量的取值进行无偏、最优估计的一种方法;从插值角度讲是对空间分布的数据求线性最优、无偏内插估计一种方法[14]。克里格法的适用条件是区域化变量存在空间相关性,云斑天牛幼虫种群的空间分布特征是区域化的变量,并且假设其具有一定的空间相关性。因此,在得到最优拟合模型后,利用普通Kriging插值法对云斑天牛幼虫数量的空间格局进行插值分析,在Surfer 8.0软件的支持下生成空间分布图。
3 结果与分析
3.1 调查样地受危害基本情况
调查样地的林分树高、胸径及受害情况见表1。3个调查样地受害白蜡的平均胸径在7.10~9.84 cm之间,平均树高在7.32~10.08m之间。根据样地受害程度不同分为轻度受害、中度受害和重度受害林,有虫株数在8.8%~32.7%之间。

表1 调查样地基本情况统计Table 1 Basic information of the survey stands
3.2 云斑天牛幼虫的垂直分布
基于SPSS Statistics 22.0软件,对不同受害林分(轻度、中度、重度)和综合(3种样地分不同高度幼虫数量的均值)样地幼虫垂直分布的差异显著性检验结果显示(图1),云斑天牛幼虫在白蜡树上的分布具有明显(α=0.05)的垂直分布特征,主要在基部往上1m以内的高度集中分布,该高度的幼虫分布在重度、中度、轻度受害林和综合样地中分别占67.8%、77.4%、82.1% 和75.0%;高度在1m以上时,幼虫的分布与1m以下分布数量均呈现极显著(p<0.01)差异,而1m以上各高度区域幼虫垂直分布差异不显著(p>0.1),表明幼虫在白蜡树1m以上分布均很少。

图1 不同受害林分云斑天牛幼虫垂直分布(α=0.05)Fig.1 Statistical characteristics of the vertical distributions of the larva of B.horsfieldi in different forests(α=0.05)
3.3 云斑天牛幼虫的半变异函数及空间分布分析
基于GS+软件,对不同危害程度的样地云斑天牛幼虫空间分布进行地统计学分析,其种群变异函数理论模型、拟合参数及分布型见表2。
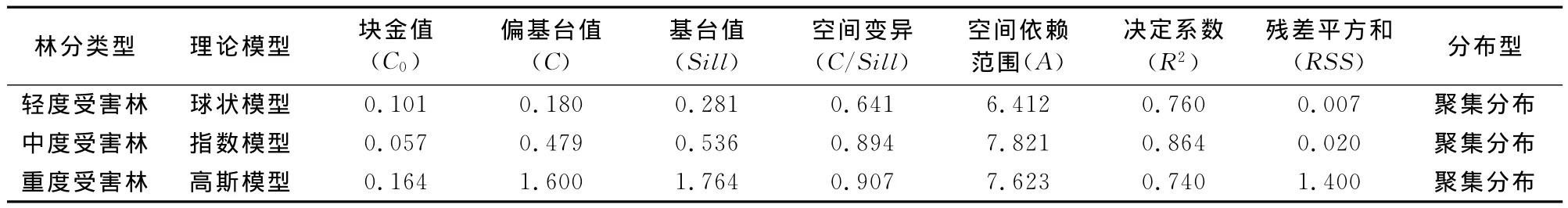
表2 云斑天牛幼虫种群数量的空间分布分析Table 2 Analysis of spatial distribution pattern of the population for the larvae of B.horsfieldi
3.3.1 轻度受害林 由表2和图2可以看出,轻度受害林中云斑天牛幼虫种群的最优拟合模型为球状模型,决定系数R2为0.760,说明幼虫的空间分布具有空间依赖性,其强度随距离的增大而减小。变程(A)为6.412m,这说明在轻度受害林分内任何两样点之间距离<6.412m时云斑天牛幼虫数量都有一定的空间相关或依赖关系,而偏基台值与基台值(C/Sill)的比值主要反映这种空间相关性的强弱程度,该值越大,说明空间相关性越强。在轻度危害林中云斑天牛幼虫的空间变异性64.1%来自于空间自相关现象,证明幼虫的空间分布格局为较强的聚集分布。半变异函数理论解释幼虫种群分布时块金值(C0)产生的可能原因有:试验样地为苗圃地,经常进行林下喷农药除草,以及枝干修剪,人为活动从而使幼虫数量有所减少;试验地内同时进行云斑天牛引诱试验,也可能影响其分布。
由云斑天牛幼虫种群数量的矢量等值线平面图可以看出,该样地中幼虫聚集分布于几个明显的聚集点,从聚集点中心向四周幼虫的数量逐渐减小,反映幼虫的聚集强度和空间尺度,也说明了其扩散方向。由图2半变异函数曲线变化趋势可以看到,当采样间距(h)达到变程(A=6.412m)之后直到20 m左右,随采样距离函数值保持稳定并略有下降,说明该林分内任意两点间距离大于6.412m后幼虫种群数量不会再有空间依赖关系或者空间上有连续性,即不会发生扩散;但是,当采样距离(h)>20 m时,函数值出现了一个小的增量,表明幼虫可能存在潜在扩散的趋势,所以以后的研究也应适当增大抽样的空间尺度。
3.3.2 中度受害林 由表2和图3可以看出,中度受害林中云斑天牛幼虫种群的最优拟合模型为高斯模型,决定系数R2为0.864,说明幼虫的空间分布具有很高的空间依赖性,其强度随距离的增大而减小,且模型的拟合度较轻度危害林还要高。变程(A)为7.821m,这说明在中度受害林分内任何两样点之间距离<7.821m时云斑天牛幼虫数量都有一定的空间相关或依赖关系。中度危害林中空间自相关现象可以解释89.4%的云斑天牛幼虫的空间变异性,证明幼虫的空间分布格局为很强的聚集分布,样地中幼虫聚集性要比轻度危害林(64.1%)强。该样地块金值(C0)产生的可能原因有:试验样地为苗圃地,经常进行林下喷农药除草,以及枝干修剪,人为活动从而使幼虫数量有所减少;样地中白蜡树较高,3m以上的排粪孔数量统计不完全而造成一定误差。

图2 轻度受害林中云斑天牛幼虫种群空间分布的变异函数图与矢量等值线平面Fig.2 Variogram and overlaying the contour and vector plots of the population number for the larva of B.horsfieldi in lightly infested stand of forest
由云斑天牛幼虫种群数量的矢量等值线平面图也可以看出:该样地中幼虫聚集性以及危害程度要比轻度危害林强:每株白蜡的幼虫平均头数以及分布范围明显大于轻度危害林。几个明显的聚集点从聚集点中心向四周幼虫的数量逐渐减小,同样反映该样地幼虫的聚集强度和空间尺度,也说明了其扩散方向。由图3半变异函数曲线变化趋势可以看到,当采样间距(h)达到变程(A=7.821m)之后函数值基本稳定于基台值(sill=0.536)附近,说明该林分内任意两点间距离>7.821m后幼虫种群数量不会再有空间依赖关系或者空间上有连续性,即扩散的可能性不明显。

图3 中度受害林中云斑天牛幼虫种群空间分布的变异函数图与矢量等值线平面Fig.3 Variogram and overlaying the contour and vector plots of the population number for the larva of B.horsfieldi in medium infested stand of forest
3.3.3 重度受害林 由表2和图4可以看出,重度受害林中云斑天牛幼虫种群的最优拟合模型为指数模型,决定系数R2为0.740,说明幼虫的空间分布具有显著空间依赖性,其强度也随距离的增大而减小,且模型的拟合度介于轻度与中度之间。变程(A)为7.623m,这说明在重度受害林分内任何2样点之间距离<7.623m时云斑天牛幼虫数量都有一定的空间相关或依赖关系。重度危害林中空间自相关现象对云斑天牛幼虫的空间变异性的解释度高达90.7%,证明幼虫的空间分布格局为很强的聚集分布。3块样地种,该样地中幼虫聚集性最强。该样地块金值(C0)产生的可能原因有:试验样地白蜡树较高,3m以上的排粪孔数量统计不完全而造成一定误差;试验地内为防治云斑天牛进行过注干输液,可能影响其幼虫的分布。
由云斑天牛幼虫种群数量的矢量等值线平面图也可以看出,该样地中幼虫聚集性最强,每株白蜡的幼虫平均头数以及分布范围明显大于前两者,但是聚集点数量相当。几个明显的聚集点从聚集点中心向四周幼虫的数量逐渐减小,同样反映该样地幼虫的聚集强度和空间尺度,也说明了其扩散方向。由图4半变异函数曲线变化趋势可以看到,当采样间距(h)达到变程(A=7.623m)之后直到17m左右函数值出现明显的降低趋势,而当采样距离再次增大后函数值又出现了上升的态势,并且已经超过了高斯模型的基台值(sill=1.764),但是并没有超过采样距离为9m左右时的函数值(2.34),所以假设采样距离(h)继续增大半变异函数值和变程的变化就变得不明显,所以以后的研究也应适当增大抽样的空间尺度。矢量等值线平面图中可以看出,边缘的分布密度较大,原因可能是因为旁边有一片杨树林,对云斑天牛成虫的取食和产卵有一定的影响,从而也影响其幼虫的空间分布。

图4 重度受害林中云斑天牛幼虫种群空间分布的变异函数图与矢量等值线平面Fig.4 Variogram and overlaying the contour and vector plots of the population number for the larva of B.horsfieldi in heavily infested stand of forest
4 结论与讨论
了解昆虫的空间分布及动态变化对有效的进行虫害管理与防治具有重要意义[25]。传统的空间格局分析方法难以区别不同空间格局,主要是因为对昆虫种群数量空间分布信息与依赖范围的获取不够充分,加之可能由于不同空间拟合方法的运用使得同一组数据出现不同的理论分 布型[17,23,26];而地统计学分析方法则弥补了这些不足,对昆虫在林间的实际分布有更直观的反映,并可以通过不同时空尺度上昆虫种群空间结构特征的对比揭示其动态规律,研究昆虫聚集、扩散等行为所带来的影响[7]。
云斑天牛幼虫在白蜡树上的垂直分布特征表明(图1),云斑天牛幼虫集中分布在1m以下的高度区域,平均分布率可以达到75.0%,这与杨桦[2]等的研究结果一致;但就1m以上高度而言,本研究显示1m以上各高度区域幼虫分布均呈现不显著的特征,分布量在35.0%以下,且随受害程度的增强有明显的增加趋势,这说明幼虫会优先分布在1 m以下的区域,可能当1m以下虫量达到一定数值时,才会选择更高的高度,这或许与云斑天牛种内竞争[2]、当地林分的采光条件[22]以及气候条件如降雨密切相关。同时,当地云斑天牛在白蜡林中的防治应该多集中在1m以下进行,且提倡“预防为主,防治结合”的方针,因为在受害程度较低时1m以上的幼虫分布最少(18.0%),且会随着受害程度加重而增加,早防治可能会大大提高防止效率,节约防治成本。
在受害程度不同(轻度、中度和重度危害)的白蜡林中,云斑天牛幼虫空间分布格局均呈现较明显的(R2>0.7)聚集分布的状态,但是其最佳拟合模型却不尽相同:依次为球状、指数和高斯模型(表2),这就表示各种受害程度林分中幼虫的聚集状态时不同的,且聚集性的强度也不同:危害越严重,聚集强度越大。就其扩散的可能性而言,目前的观测数据说明轻度和中度危害林中天牛幼虫的潜在扩散能力不强(图2、3),而重度危害林中半边以函数曲线(图4)在采样距离>17m后函数值有一种持续上升的态势,虽然目前的数据无法断定其扩散强度,但是可以确定的是其扩散的可能性很大。所以该地的云斑天牛防治最主要的是找到几个天牛分布的聚集点,特别是对于重度危害林,再沿着扩散方向逐步防治,而不是大面积的撒网式防治,这样就可以节约防治成本并降低对林分的干扰。
在我国云斑天牛为2a1代,由于整个世代中绝大部分(>90%)的时间都隐蔽于树干中生活,这给直接调查云斑天牛的种群数量造成较大困难[2,5]。李友常[2,19]等在调查杨树上光肩星天牛种群空间格局时采用排粪孔、羽化孔和刻槽的数量作为幼虫、成虫、卵的调查指标,而云斑天牛整个幼虫期只有1个排粪孔,羽化成成虫后钻蛀1个羽化孔钻出,因此本试验以样地中白蜡树上排粪孔的数量为调查指标是准确可靠的。对于云斑天牛的空间格局,先前的研究均使用的是传统的统计学方法,本试验首次利用地统计学对云斑天牛幼虫空间格局进行研究,得到云斑天牛幼虫在3种受害程度不同的白蜡林中,其空间格局均呈现聚集分布,且一致性较为明显,这与用传统的统计学方法得出的空间分布型结论一致[8-11]。云斑天牛幼虫自身不具备扩散能力,成虫的飞行扩散能力及产卵习性则成为决定幼虫空间分布格局的重要因素之一[2],而补充营养寄主,以野蔷薇、梨、枫杨等树种为主,对成虫的扩散行为以及产卵刻槽具有重要影响。其次,寄主树木的树高、胸径等外部因素也会影响幼虫的种群空间特征。
在轻度危害林和重度危害林云斑天牛幼虫分布的半变异函数曲线图(图2,图4)中均可以看到,假设采样距离(h)超过一定值继续增大时半变异函数值和变程的变化可能有不同的响应,所以在以后的调查研究中需注意采样的空间尺度,增大可能对这种空间分布的分析更为全面。另外,应用地统计学对云斑天牛种群在时间上的依赖性与连续性的研究今后还需增加调查次数,获得更多资料进一步完善。
当前对云斑天牛虫害的防治,通常采用物理、化学以及生物防治相结合进行。在被害林木附近或林中存在大量成虫补充食物源的地方喷洒农药可大量毒杀成虫;在产卵期人工砸卵,可将危害率大大降低;当幼虫蛀入木质部后,可采用在蛀孔处注射久效磷或插堵毒签的方法,治虫效果达95%以上。利用天敌防治,在产卵期释放卵寄生蜂或在幼虫期释放管氏肿腿蜂、花绒寄甲可达到防治效果[3-6]。另外,在营林时,切勿与野蔷薇、梨、枫杨等云斑天牛补充营养树种混交种植。
[1] 陈世骧,谢蕴贞,郑国藩.中国经济昆虫志-天牛科[M].北京:科学出版社,1959:84-85.
[2] 杨桦,杨茂发,杨伟,等.杨树云斑天牛幼虫空间分布格局及生息坑道[J].四川农业大学学报,2010,28(2):148-152.YANG H,YANG M F,YANG W,et al.A study on the spatial distribution pattern and the living-inhabiting tunnel of the larvae of Batocera horsfieldi (Hope)[J].Journal of Sichuan Agricultural University,2010,28(2):148-152.(in Chinese)
[3] 梅爱华.云斑天牛幼虫空间分布型及抽样技术[J].昆虫知识,1997,34(2):94-95.Mei A H.Studies on spatialdistribution and sampling technique of Batocera horsfieldi[J].Entomological Knowledge,1997,34(2):94-95.(in Chinese)
[4] 吴开明,张建强,代方银,等.云斑天牛危害桑树及生物学特性研究[J].蚕业科学,1995,21(3):53-54.
[5] 李建庆,杨忠岐,张雅林,等.杨树上云斑天牛种群的空间格局及抽样技术[J].昆虫学报,2009,52(8):860-866.LI Y Q,YANG Z Q,ZHANG Y L,et al.Spatial distribution pattern and sampling technique for Batocera horsfieldi(Coleoptera:Cerambycidae)in poplar[J].Acta Entomologica Sinica,2009,52(8):860-866.(in Chinese)
[6] 严敖金,嵇保中,钱范俊,等.云斑天牛Batocera horsfieldi(Hope)的研究[J].南京林业大学学报,1997,21(1):1-6.
[7] 马宁远,王惠卿,张伟,等.基于地统计学的新疆棉田烟粉虱(Bemisiatabaci(Gennadius))危害动态与时空分布[J].生态学报,2008,28(6):2654-2662.MA N Y,WANG H Q,ZHANG W,et al.The damage dynamics and spatio-temporal distribution of Bemisiatabaci(Gennadius)in cotton field based on GS analysis in Xinjiang[J].Acta Ecologica Sinica,2008,28(6):2654-2662.(in Chinese)
[8] 侯景儒,黄竞先.地质统计学及其在矿产储量计算中的应用[M].北京:地质出版社,1982:23.
[9] 侯景儒,黄竞先.地质统计学的理论与方法[M].北京:地质出版社,1990:21.
[10] GOOVAERTS P.Geostatistics in soil science:tate of the art and per-spectives[J].Geoderma,1999,89(1/2):1-45.
[11] GOOVAERTS P.Geostatistical approaches for incorporating elevation into the spatial interpolation of rainfall[J].Journal of Hydrology,2000,228(1-2):113-129.
[12] KUNZLI N,JERRETT M,MACK W J,et al.Ambient air pollution and ath-eroselerosis in Los Angeles[J].Environmental Health Perspectives,2005,113(2):201-206.
[13] 周国法,徐汝梅.生物地理统计学[M].北京:科学出版社,1998:3-10.
[14] 向昌盛,袁哲明.地统计方法在昆虫学研究中的应用[J].中国农学通报,2009,25(17):191-194.XIANG S C,YUAN Z M.Application of geostatistics in the research of insect[J].Chinese Agricultural Science Bulletin,2009,25(17):191-194.(in Chinese)
[15] 于新文,刘晓云.昆虫种群空间格局的研究方法评述[J].西北林学院学报,2001,16(3):83-87.YU X W,LIU X Y.Review of research methods on spatial pattern of insect population[J].Journal of Northwest Forestry University,2001,16(3):83-87.(in Chinese)
[16] 房用,刘德玺,马昭让,等.滨海盐碱地大果沙棘适应及耐盐碱试验[J].东北林业大学学报,2005,33(2):38-39.FANG Y,LIU D X,MA Z R,et al.Tests on the adaptation and salt and alkali resistance of HippophaerhamnoidesL.in seashore[J].Journal of Northeast Forestry University,2005,33(2):38-39.(in Chinese)
[17] LECOUSTRE L,FARGETTE D,et al.Analysis and mapping of the spatial spread of African cassava mosaic virus using Geostatistics and kriging technique[J].Phytopathology,1989,79(9):913-920.
[18] LIEBHOLD A M,ROSSI R E,KEMP W P.Geostatistics and geographic information systems in applied insect ecology[J].Annual Review of Entomology,1993,38:303-327.
[19] 李友常,夏乃斌,屠泉洪,等.杨树光肩星天牛种群空间格局的地统计学分析[J].生态学报,1997,17(4):393-401.LI Y C,XIA N C,TU Q H,et al.A geostatistical analysis on spatial patterns of Anoplophora glabripennis in poplars[J].Acta Ecologica Sinica,1997,17(4):393-401.(in Chinese)
[20] 宗世祥,骆有庆,许志春,等.沙棘木蠹蛾卵和幼虫空间分布的地统计学分析[J].生态学报,2005,25(4):831-836.ZONG S X,LUO Y Q,XU Z C,et al.Geostatistical analysis on spatial distrbution of Holcocerus hippophaecolus eggs and larvaes[J].Acta Ecologica Sinica,2005,25(4):831-836.(in Chinese)
[21] 潘杰,王涛,宗世祥,等.红脂大小蠹种群空间格局地统计学分析及抽样技术[J].生态学报,2011,31(1):195-202.PAN J,WANG T,ZONG S X,et al.Geostatistical analysis and samping technique on spatial distrbution patterns of Dendroctonus valens population[J].Acta Ecologica Sinica,2011,31(1):195-202.(in Chinese)
[22] 章一巧,宗世祥,刘永华,等.陕西省大黄枯叶蛾蛹的空间分布[J].生态学报,2012,30(20):6306-6317.ZHANG Y Q,ZONG S X,LIU Y H,et al.Spatial distribution of Trabala vishnou gigantina yang pupae in Shaanxi Province[J].Acta Ecologica Sinica,2012,30(20):6306-6317.(in Chinese)
[23] 高书晶,庞保平,史丽,等.小菜蛾幼虫空间格局的地统计学分析[J].昆虫知识,2004,41(4):324-327.GAO S J,PANG B P,SHI L,et al.Geostatistical analysis on spatial patterns of Plutella xylostellalarvae[J].Entomological Knowledge,2004,41(4):324-327.(in Chinese)
[24] 毕守东,邹运鼎,耿继光,等.棉蚜及龟纹瓢虫空间格局的地学统计学研究[J].应用生态学报,2000,11(3):421-424.BI S D,ZOU Y D,GENG J G,et al.Conspatial patterns of Aphisgossypii and Propylaca japonica[J].Chinese Journal of Applied Ecology,2000,11(3):421-424.(in Chinese)
[25] 邵崇斌,徐振武,韩明耀.杨树蛀干害虫空间格局的研究[J].西北林学院学报,1997,12(Supp.):33-36.SHAO C B,XU Z W,HAN M Y.A research on space distribution pattern of poplar stem borer[J].Journal of Northwest Forestry University,1997,12(Supp.):33-36.(in Chinese)
[26] 张连翔,温豁然,吕尚彬,等.昆虫种群空间格局研究中的几个检验问题[J].西北林学院学报,1996,11(3):59-65.ZHANG L X,WEN H R,LU S B,et al.Some test problems in the study of spatial distribution of insects populations[J].Journal of Northwest Forestry University,1996,11(3):59-65.(in Chinese)
猜你喜欢
林业资源管理(2022年6期)2023-01-18
现代园艺(2021年23期)2021-12-01
小哥白尼(野生动物)(2021年1期)2021-07-16
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
小学生必读(低年级版)(2018年10期)2019-01-04
故事作文·低年级(2018年10期)2018-10-25
作文与考试·小学低年级版(2015年11期)2015-07-17
