
长链非编码RNA 的功能与心血管疾病的研究进展
2014-12-31 11:27:02金亮综述陈筱潮审校
心血管病学进展 2014年6期
金亮 综述 陈筱潮 审校
(1.中山大学孙逸仙纪念医院心内科,广东 广州 510120)
人类基因组测序的完成打开了后基因组研究的大门。ENCODE (encyclopedia of DNA elements)研究指出人类几乎所有的常染色质基因组中的核苷酸都被转录,并确认80% 的基因组都具备某种特定功能[1]。但只有一小部分(<3%)转录本来源于蛋白质的编码基因[2],剩下的非编码转录本可能通过不同的机制影响基因的表达。非编码RNA 根据其长度可分为小非编码RNA(>200 nt)和长非编码RNA(>200 nt),其中小编码RNA 包括miRNAs、piRNA 及siRNA。当前研究证实非编码RNA 参与许多生物学功能的调控,现已明确microRNA 在心力衰竭、冠状动脉粥样硬化、心肌缺血和再灌注损伤等病理过程中对基因表达的改变起关键作用。然而,lncRNA 在心血管系统疾病中的研究才刚刚起步。现就对长链非编码RNA(long noncoding RNAs,lncRNAs)的功能及其在心血管系统疾病中的相关作用的研究做一综述。
1 lncRNA 的结构
lncRNA 是一类转录本长度超过200 nt 的功能性RNA 分子,存在于胞核或胞浆中。lncRNA 根据其基因组的定位和背景可以分为正义(sense)、反义(antisense)、双向(bidirectional)、基因内(intronic)、基因间(intergenic)[3]。与蛋白编码RNA 相比,lncRNA 的保守性要差得多,但在其分子内部,却含有较为保守的序列或二级结构,且其表达具有时空特异性,这些现象都提示lncRNA 具有重要的生理生化功能[3]。有些lncRNA 和mRNA 一样具有5'帽子结构和PolyA 尾结构,通过剪切加工而成熟。在大多数情况下,lncRNA通过不同的作用机制参与表遗传学的调控、转录和转录后调控。同时,lncRNA 还可作为小RNAs 的前体和亚细胞结构的组织框架。特定的lncRNA 发生结构突变,引起表达水平及定位的改变,已被证明参与许多疾病的发生发展[4-6]。
2 lncRNA 的功能
大部分真核基因组经转录,产生了上万种无或少有蛋白编码能力的lncRNA,最初被认为是RNA 聚合酶Ⅱ的转录副产物,不具备生物学功能[7],进一步的研究表明它们通过与DNA、RNA 或蛋白质相互作用调控基因的表达。lncRNA 按照功能机制的不同可分为:信号分子(signal molecule)、诱饵分子(decoy molecule)、引导分子(guide molecule)和骨架分子(Scaffold molecule)等4 类分子(图1)[8]。下面依据lncRNA 参与的生物学功能分别阐述。
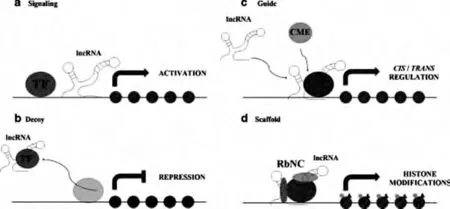
图1 lncRNA 分子机制示意图
2.1 lncRNA 参与表观基因的沉默
INK4 位点反义非编码RNA(ANRIL)是周期蛋白依赖性激酶抑制因子2B(cyclin-dependent kinase inhibitor 2B,CDKN2B,即p15INK4b)基因的反义转录物,参与抑癌基因周期蛋白依赖性激酶抑制因子2A(cyclin-dependent kinase inhibitor 2A,CDKN2A,即p16INK4a)基因的沉默[9]。p15 和p16 都是细胞周期调控因子,与肿瘤的发生密切相关。ANRIL 通过与多梳抑制复合体1(PRC1)中的组成蛋白Pc/chromobox 7(CBX7)相互作用,再通过募集PRC2 顺式介导p16 的转录抑制。研究发现在前列腺癌组织中升高的CBX7 和ANRIL 与降低的p16 密切相关[10]。另一个同源异型框基因反义基因间 RNA (HOX anti-sense intergenic RNA,HOTAIR)转录自同源异型框基因C(homeobox C,HOXC)位点,通过募集PRC2 反式抑制HOXD 位点的转录[11]。HOTAIR 的过表达将引起其连接的PRC2 复合体在全基因组范围内的重新定位,使若干个抑癌基因沉默促进了肿瘤的进展和转移。此外,Xist 在没有聚合酶Ⅱ(polymerase Ⅱ,polⅡ)的情况下建立了一个特别的核区,将要失活的X 染色体被带入这个区域后聚集并在表面形成“外套”,随后获得抑制性的组蛋白修饰(如甲基化),导致失活X 染色体启动子区域的CpG 岛甲基化,X 染色体失活[12]。
2.2 lncRNA 参与细胞凋亡和细胞周期调控
研究表明,lncRNA 也参与细胞生长的调控,这主要是通过对细胞周期和凋亡调控来实现。生长阻滞特异转录物5(growth arrest-specific transcript 5,Gas5)通过模拟糖皮质激素应答元件结合糖皮质激素受体的DNA 结合结构域,阻止糖皮质激素受体与应答元件的相互作用,从而抑制下游基因的表达,促进细胞凋亡[13]。在细胞DNA 受到损伤时,p53 能结合到lincRNA-p21 的启动子区,激活lincRNA-p21 转录,该RNA 与核不均一核糖核蛋白K(heterogeneous nuclear ribonucleoprotein K,hnRNP-K)结合后能特异性地定位到目的基因的启动子区,导致这些基因的转录受阻,从而引起细胞凋亡[14]。周期素D1(CCND1)基因上游区域转录出一段lncRNA,参与CCND1 基因转录的负向调节。通过连接一个RNA 连接蛋白TLS,形成lncRNA-TLS 复合体定位于CCND1 启动子区,变构抑制组蛋白乙酰转移酶活性,阻止CCND1 基因的转录[15],抑制细胞周期。
2.3 lncRNA 参与剪切调控
剪切调控作为一种转录后修饰广泛参与多种生物学过程。肺腺癌转移相关转录本1 (metastasis-associated in lung adenocarcinoma transcript,MALAT-1)与早期非小细胞肺癌相关[16]。两个新近的研究发现,MALAT-1 通过与剪切机器中富含丝氨酸和精氨酸家族的核磷蛋白相互作用调控可变剪切[17-18]。据认为,MALAT-1 是作为一个结构停泊位点聚集特异性的剪切因子(如高效可变剪切所必需的磷酸化SR 蛋白)参与相关剪切,并调节前mRNA 剪切因子分布至核斑点处参与SR 蛋白的磷酸化,改变剪切模式,使MALAT-1缺失的细胞误定位和未磷酸化的SR 蛋白水平增加,导致外显子失表达数目增加[18]。
2.4 lncRNA 参与翻译水平的调控
β-分泌酶1 反义转录物(BACE1-antisense transcript,BACE1-AS)是编码在染色体11q23.3 上的保守RNA,转录自BACE1 反义链。BACE1 蛋白通过切割β淀粉样肽前体,产生更多的β 淀粉样肽[19]。而升高的β 淀粉样肽、BACE1 蛋白和BACE1-AS 水平与老年痴呆症相关,这预示着BACE1-AS 表达的改变可能参与该疾病的进展[19]。BACE1-AS 与正义BACE1 mRNA相结合将增强BACE1 mRNA 的稳定性,从而增加BACE1 蛋白在脑内的表达,最终导致β 淀粉样肽的有毒性聚集[19]。
2.5 lncRNA 参与小RNA 的形成
全基因组研究表明,lncRNA 长链可能是长度>200 kb的小RNA 的前体[20]。通过不同的酶剪切修饰,如Drosha 酶和Dicer 酶序列剪切产生的miRNA 其前体也可能是一段长链lncRNA[21],也可能通过加工lncRNA 产生piRNA[22]。
3 lncRNA 与心血管疾病
lncRNA 作为一种新的生物调节模式在心血管疾病中的研究才刚刚起步。其在疾病中的调控及其作用机制尚不明确,但已有一些研究指出lncRNA 促进胚胎发育中正常血管的形成,其表达失衡会引起心脏发育不全、动脉粥样硬化性血管病及心肌梗死等。深入研究相关机制有助于进一步阐明疾病发生发展过程,也为新的治疗提供潜在靶点。
3.1 lncRNA 与心脏、血管发育
心脏发育研究表明,胚胎在不同的发育阶段lncRNA表达存在很大差异,很可能是参与正常心脏的发育的新机制[23]。Braveheart(Bvht)是一个在大鼠体内与心脏相关的lncRNA,通过对胚胎干细胞不同的分化处理,发现Bvht 对于胚胎从新生中胚层到成形心肌是必需的[24],通过与PRC2 亚单元SUZ12 相互作用调控心肌表型[24]。同时,Bvht 对于诱导MesP1(一种保守的重要转录因子)及其下游的靶标包括核心心脏转录因子如Gata4,Gata6,Hand1,Hand2,Tbx2 和Nkx2.5等是必需的[25]。Fendrr 是一个侧中胚层特异性的lncRNA,控制中胚层的分化及通过连接组蛋白重构复合体PRC2 和TrxG/MLL 使其分化成为心脏和体壁[26]。缺失Fendrr 的胚胎表现为中胚层特异性转录因子的调节异常以及PRC2 在富集位点的减少。lncRNA Kcnq1ot1 通过调控一种钾离子通道编码基因(Kcnq1)的表达参与心脏的正常发育[27]。
3.2 lncRNA 与冠状动脉粥样硬化
全基因组关联研究发现,在lncRNA 心肌梗死相关转录本(MIAT)中有六个SNP 与心肌梗死相关。一个SNP(A11741G)使MIAT 在体外转录增加1.3倍[28]。编码MIAT 的基因有5 个外显子,体外翻译分析显示,MIAT 不编码任何蛋白质,很可能是一个功能性的RNA,而体外功能分析显示,第五外显子上的SNP 变异增加了MIAT 的转录,这项研究指出SNP 引起的MIAT 表达水平的改变在心肌梗死中可能起作用[28]。
Leung 等研究发现RAS 系统也可以通过lncRNA参与心肌、血管病变的发生发展[29]。用血管紧张素Ⅱ培养血管平滑肌细胞,评估其转录组反应,发现许多蛋白编码mRNA 和非编码lncRNA 受血管紧张素Ⅱ调控,且受调控的Lnc-Ang362 能够作为miR-221 和miR-222 宿主转录本,这两个miRNA 都与细胞增殖有关。用siRNA 干扰Lnc-Ang362 的表达降低了血管平滑肌细胞增殖[29]。这为血管紧张素Ⅱ在冠心病研究中提供新的思路,相关非编码转录本也可能成为临床药物作用的新位点。
ANRIL 是惟一一个发现的位于染色体9p21 区域的与动脉粥样硬化性血管病敏感性有关的转录物[30]。ANRIL 的功能研究显示它可作为PCG 复合物(polycomb group,PCG)的骨架与PRC1、PRC2 相连接,通过表观顺式作用抑制p16INK4a/p15INK4b的表达[10]。阻止血管平滑肌细胞由G1 期进入S 期,从而抑制血管重塑,防止病理性血管内膜增生,抑制动脉粥样硬化的形成。
3.3 lncRNA 与心力衰竭
lncRNA 可通过调节心肌的病理重构和心肌肥厚参与心力衰竭的进展。心肌重构一个最明显的改变是α-心肌肌球蛋白重链(α-myosin heavy chain,α-MHC)和β-心肌肌球蛋白重链(β-myosin heavy chain,β-MHC)的比例倒置,这种亚型的改变常发生在糖尿病、甲状腺功能低下压力过负荷以及能量缺乏的病理情况下[31]。进一步的研究显示,这种改变很可能是由来源于α-MHC 和β-MHC 基因间4.5 kb 的转录本NATs 所介导[32]。新近研究显示,lncRNA 心脏肥大相关因子(CHAF)通过内源性吸附miR-489,使其表达水平下调。而使miR-489 的靶标髓样分化初级反应基因88(myeloid differentiation primary response gene 88,Myd88)上调,介导心肌肥厚[33]。此外,与心肌梗死相关lncRNA 的调节异常也是心力衰竭的重要原因之一。全基因组SNP 分析证实MIAT 基因的改变与心肌梗死的易感性有关[28]。
4 展望及研究方向
随着研究的深入,我们对lncRNA 的功能及与人类疾病的关系也逐渐清晰,lncRNA 不仅能和DNA、RNA 或蛋白质相互作用调节基因的表达,还可能作为结构分子影响其他RNA(如miRNA)的功能。由于其表达具有时间空间特异性,研究确定lncRNA 在正常人中的表达水平,以及具有临床诊断意义的变化程度,对于其作为相关疾病的生物分子标志物为临床诊断提供新的思路。研究中通过高通量芯片技术找到对应疾病的lncRNA 并阐明其表达水平的变化,进一步揭示其主要功能和作用机制,有助于发现新的疾病调控机制,为临床的诊断和治疗提供新的靶点。
[1]Birney E,Stamatoyannopoulos JA,Dutta A,et al.Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project[J].Nature,2007,447(7146):799-816.
[2]Maher B.ENCODE:The human encyclopaedia[J].Nature,2012,489(7414):46-48.
[3]Ponting CP,Oliver PL,Reik W.Evolution and functions of long noncoding RNAs[J].Cell,2009,136(4):629-641.
[4]Struhl K.Transcriptional noise and the fidelity of initiation by RNA polymeraseⅡ[J].Nat Struct Mol Biol,2007,14(2):103-105.
[5]Wang KC,Chang HY.Molecular mechanisms of long noncoding RNAs[J].Mol Cell,2011,43(6):904-914.
[6]Wapinski O,Chang HY.Long noncoding RNAs and human disease[J].Trends Cell Biol,2011,21(6):354-361.
[7]Yap KL,Li S,Munoz-Cabello AM,et al.Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a[J].Mol Cell,2010,38(5):662-674.
[8]Rinn JL,Kertesz M,Wang JK,et al.Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs[J].Cell,2007,129(7):1311-1323.
[9]Collins LJ,Chen XS.Ancestral RNA:the RNA biology of the eukaryotic ancestor[J].RNA Biol,2009,6(5):495-502.
[10]Kino T,Hurt DE,Ichijo T,et al.Noncoding RNA gas5 is a growth arrest-and starvation-associated repressor of the glucocorticoid receptor[J].Sci Signal,2010,3(107):a8.
[11]Huarte M,Guttman M,Feldser D,et al.A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response[J].Cell,2010,142(3):409-419.
[12]Wang X,Arai S,Song X,et al.Induced ncRNAs allosterically modify RNAbinding proteins in cis to inhibit transcription[J].Nature,2008,454(7200):126-130.
[13]Ji P,Diederichs S,Wang W,et al.MALAT-1,a novel noncoding RNA,and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer[J].Oncogene,2003,22(39):8031-8041.
[14]Bernard D,Prasanth KV,Tripathi V,et al.A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression[J].EMBO J,2010,29(18):3082-3093.
[15]Tripathi V,Ellis JD,Shen Z,et al.The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation[J].Mol Cell,2010,39(6):925-938.
[16]Faghihi MA,Modarresi F,Khalil AM,et al.Expression of a noncoding RNA is elevated in Alzheimer's disease and drives rapid feed-forward regulation of beta-secretase[J].Nat Med,2008,14(7):723-730.
[17]Affymetrix ENCODE Transcriptome Project;Cold Spring Harbor Laboratory ENCODE Transcriptome Project.Post-transcriptional processing generates a diversity of 5'-modified long and short RNAs[J].Nature,2009,457(7232):1028-1032.
[18]Lee Y,Kim M,Han J,et al.MicroRNA genes are transcribed by RNA polymerase Ⅱ[J].EMBO J,2004,23(20):4051-4060.
[19]Aravin AA,Hannon GJ,Brennecke J.The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race[J].Science,2007,318(5851):761-764.
[20]Zhu JG,Shen YH,Liu HL,et al.Long noncoding RNAs expression profile of the developing mouse heart[J].J Cell Biochem,2014,115(5):910-918.
[21]Klattenhoff CA,Scheuermann JC,Surface LE,et al.Braveheart,a long noncoding RNA required for cardiovascular lineage commitment[J].Cell,2013,152(3):570-583.
[22]Kriegmair MC,Frenz S,Dusl M,et al.Cardiac differentiation in Xenopus is initiated by mespa[J].Cardiovasc Res,2013,97(3):454-463.
[23]Grote P,Wittler L,Hendrix D,et al.The tissue-specific lncRNA Fendrr is an essential regulator of heart and body wall development in the mouse[J].Dev Cell,2013,24(2):206-214.
[24]Korostowski L,Sedlak N,Engel N.The Kcnq1ot1 long non-coding RNA affects chromatin conformation and expression of Kcnq1,but does not regulate its imprinting in the developing heart[J].PLoS Genet,2012,8(9):e1002956.
[25]Ishii N,Ozaki K,Sato H,et al.Identification of a novel non-coding RNA,MIAT,that confers risk of myocardial infarction[J].J Hum Genet,2006,51(12):1087-1099.
[26]Leung A,Trac C,Jin W,et al.Novel long noncoding RNAs are regulated by angiotensin Ⅱin vascular smooth muscle cells[J].Circ Res,2013,113(3):266-278.
[27]Burd CE,Jeck WR,Liu Y,et al.Expression of linear and novel circular forms of an INK4/ARF-associated non-coding RNA correlates with atherosclerosis risk[J].PLoS Genet,2010,6(12):e1001233.
[28]Giger J,Qin AX,Bodell PW,et al.Activity of the beta-myosin heavy chain antisense promoter responds to diabetes and hypothyroidism[J].Am J Physiol Heart Circ Physiol,2007,292(6):H3065-H3071.
[29]Haddad F,Qin AX,Bodell PW,et al.Intergenic transcription and developmental regulation of cardiac myosin heavy chain genes[J].Am J Physiol Heart Circ Physiol,2008,294(1):H29-H40.
[30]Liu F,Zhou LY,Long B,et al.A long noncoding RNA,CHRF regulates cardiac hypertrophy by targeting miR-489[J].Circ Res,2014,114(9):1377-1388.
猜你喜欢
今日农业(2021年11期)2021-08-13 08:53:24
四川轻化工大学学报(自然科学版)(2021年1期)2021-06-09 06:12:12
汉字汉语研究(2020年2期)2020-08-13 07:52:48
电子制作(2019年22期)2020-01-14 03:16:24
疯狂英语·新读写(2018年3期)2018-11-29 22:37:11
山东冶金(2018年5期)2018-11-22 05:12:46
西安建筑科技大学学报(自然科学版)(2016年1期)2016-11-08 12:15:18
铁道科学与工程学报(2015年4期)2015-12-24 12:11:01
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:58
遗传(2014年3期)2014-02-28 20:58:49
