
西双版纳不同胶林复合生态系统的生物量及其分配特征
2014-12-27 09:24萧自位王秀华王小李高丽洪唐建维
中南林业科技大学学报 2014年2期
萧自位,王秀华,郑 丽,王小李,高丽洪,唐建维
(1.中国科学院 西双版纳热带植物园 热带植物资源开放实验室,云南 勐腊 666303;2. 中国科学院研究生院,北京 100049;3.西双版纳州土壤肥料工作站,云南 景洪 666100;4. 云南省科学技术情报研究院,云南 昆明650051;5. 西双版纳州科技情报研究所,云南 景洪 666100)
西双版纳不同胶林复合生态系统的生物量及其分配特征
萧自位1,2,王秀华3,郑 丽4,王小李4,高丽洪5,唐建维1
(1.中国科学院 西双版纳热带植物园 热带植物资源开放实验室,云南 勐腊 666303;2. 中国科学院研究生院,北京 100049;3.西双版纳州土壤肥料工作站,云南 景洪 666100;4. 云南省科学技术情报研究院,云南 昆明650051;5. 西双版纳州科技情报研究所,云南 景洪 666100)
根据4种不同胶林种植模式的样地调查资料,运用已有生物量模型和收获法获取生物量,分析了4种胶林种植模式的生物量组成及其分配特征。结果表明,橡胶 + 茶和橡胶 + 咖啡和橡胶 + 多种植物模式的总生物量分别高于纯胶林69.636 t·hm-2、114.648 t·hm-2和81.924 t·hm-2。活植物体生物量是各胶林类型总生物量的主要组成部分,占97. 79%~98.76%;死植物体生物量所占比例为1.24%~2.21%。各胶林组合模式中橡胶树的生物量均存在“超产”现象(D>0),表明植物多样性对各胶林组合模式中橡胶树的生产力存在正效应,能促进其生物量的积累。
橡胶林;生物量;农林复合生态系统;西双版纳
生物量是在某一时刻调查时单位面积上积存的有机物质,它不仅能够反映生态系统在一定时间内的生产力,而且也是研究生态系统结构、功能、能量流动和物质循环的重要参数[1-5]。作为陆地生态系统的主体,森林生态系统以全球陆地表面31% 的面积储存了陆地生态系统生物量的 70%~90%[5-6],其中热带地区的森林生物量占全球森林生物量的2/3以上[7]。虽然自上世纪60年代以来世界各国的学者对热带森林的生物量进行了广泛研究,但其结果依然存在着许多不确定性[7-9]。因此,对热带森林生物量的进一步研究,将有利于我们深入了解热带森林的动态,养分贮存状况及因森林采伐而引起的碳释放[10]。
橡胶树Hevea brasiliensis为大戟科植物,原产于南美洲亚马逊河流域的热带雨林中,是一种重要的热带经济树种,在热带地区尤其是南亚和东南亚广泛种植。橡胶林作为热带地区主要的人工森林生态系统,至2010年全球橡胶种植园的面积已超过10 000万hm2,仅亚洲就占了90 % 以上;中国是亚洲主要的橡胶种植区域之一,其面积已超过100万hm2[11]。橡胶种植面积的不断扩大,不仅导致了大量热带天然林和次生林被砍伐[12-13],而且大部分的植胶区均以单一的纯胶林为主,由此导致了十分严重的胶林水土流失、土壤肥力衰退、生态环境恶化和生物多样性降低等问题[14-15]。为此,寻求和探索出生物资源的开发与保护、生态与经济效益协调统一的合理开发途径已刻不容缓。胶林复合生态系统作为一种可持续的农林复合生态系统类型,受到了许多学者的广泛关注[16-17]。它不仅可以利用群落内不同物种间的生态互补功能,提高养分的吸收利用效率、减缓水土流失、维持局部小气候稳定;而且还能提高系统内的生物多样性、充分利用其水、肥、光、热等资源,提高生态系统的生产力[17-20]。此外,研究农林复合生态系统中生物多样性与生物量的关系也是了解生物多样性与生态系统功能关系的重要途径。西双版纳是我国主要的橡胶种植地之一,其面积占全国胶林总面积的24 %[21]。虽然近年来许多学者对西双版纳地区不同林龄和海拔的橡胶林生物量进行了深入研究[21-23],但这些研究都局限于单一的纯胶林,对该区胶林复合生态系生物量的研究报道较少,仅见于唐建维等[17]所进行的以橡胶为主的人工雨林生物量研究。因此,本研究以纯胶林为对照,试对西双版纳3种不同胶林复合生态系统的生物量及其分配特征进行分析,并探讨随植物多样性的增加其生物量的动态变化规律,以期为进一步估算胶林复合生态系统的碳储量提供基础数据,同时也为该区农林复合生态系统的发展提供科学依据。
1 研究地区与研究方法
1.1 自然概况
研究地点位于中国科学院西双版纳热带植物园葫芦岛的东区。该区属西南热带季风气候,海拔高度为560 m,一年可分为干热、湿热及雾凉3季,3~5月为干热季,气温较高,降雨量少;6~10月为雨季,气候湿热,全年85 %的雨水集中在此期间;1~2月为雾凉季,降水量减少,但早晚浓雾弥漫,空气湿度较大。年平均气温21.6 ℃,最热月(5月)平均气温25.3 ℃,最冷月(1月)平均气温15.5 ℃,年均降雨量1 557.0 mm,年平均相对湿度86 %,年日照时数1 828.0 h,有雾日数173.7 d,终年无霜,土壤为砖红壤,土层深厚,pH值在4.4~5.5之间。
1.2 研究方法
1.2.1 样地的选择与建立
2011年1月在西双版纳勐腊县勐仑镇葫芦岛的东区选择具有相同年龄(50 a)、相似坡度、海拔、坡向、种植方式和管理方式的4种不同胶林类型(橡胶、橡胶 + 茶Camellia sinensis、橡胶 + 咖啡Coffea liberica和橡胶 + 多种植物multi- plants作为研究对象,并在每种模式中建立3块样地作为重复,其面积为 500 m2(25 m × 20 m)。对每块样地内所有胸径D≥ 2.0 cm的植物个体进行每木调查,测量每株的胸径(1.3 m)、 树高、 冠幅并编号,其中茶树和咖啡树测其基径(Dg)。同时,用红油漆在所测位置做标记。各胶林类型样地的基本特征见表1。

表1 西双版纳不同胶林类型样地的基本特征Table 1 Plot characteristics of monoculture rubber plantation and different mixed rubber-tree plantations in Xishuangbanna
1.2.2 样地的群落学特征
根据样地调查数据可知,在4同橡胶林种植模式中,以橡胶 + 多种植物组合模式的群落最为复杂,其乔木层包含了40多种植物,可划分为3个亚层(Ⅰ、Ⅱ、Ⅲ)。乔木层Ⅰ以1969年种植的橡胶树为主,乔木Ⅱ层由木奶果Baссaurea ramif l ora(1979 年 种 植 ) 、 望 天树Parashorea chinensis、催吐萝芙木Rauvolf i a vomitoria、垂叶榕Ficus benjamana、大叶藤黄Carcinia xanthochvmus、小叶藤黄Carcinia cowa等物种所组成,乔木 Ⅲ 层则是许多乔木的幼树如圆锥木姜子Litsea liyuyingi、南洋杉Araucaria cunninghamii、披针叶楠Phoebe lanceolata、粗毛榕Ficus hirta等;灌木层主要包括小粒咖啡Coffea robusta、大粒咖啡Coffea liberica、地樱花Urena lobata、甜菜Sauropus androgynus、鱼尾葵Caryota ochlandra、狭叶龙血树Dracaena angustifolia等20余种;草本层种类也较丰富达30余种,并以千年健Homalomena occulta、山菠萝Pandanus furacatus、水蕨菜Pteridium excelsum、闭鞘姜Costus speciosus、 水 蓼Polygpnum pinnatum、 海芋Alocasis macrorrhiza、糯米香草Semnostachya menglaensis等多种草本植物为主。橡胶 + 茶和橡胶 + 咖啡两种组合模式的群落结构主要分为两层,上层乔木为橡胶,下层分别为茶树Camellia sinensis和大粒咖啡Coffea liberica;而林下的草本层因人为管理,数量十分稀少,因此在本研究中不做调查。
1.2.3 生物量测定
利用已建立的西双版纳橡胶树、茶树生物量回归模型计算橡胶树和茶树的生物量[23-24];运用Suárez-Pascua(2002)[25]所建立的咖啡树生物量模型计算咖啡树的生物量;对于橡胶 + 多种植物组合模式中其它乔木的生物量,根据其生物生态学特征的相似性,采用已建立的西双版纳季节雨林的生物量模型进行计算[26]。各种树种的生物量回归模型见表2。
另外,在橡胶 + 多种植物组合模式的每块样地内随机设置5个面积为2 m × 2 m的样方和 1 m× 1 m 的小样方,采用样方收获法[27]分别对灌木和草本植物的生物量进行调查。按茎、枝、叶、根分别称重,并称取少量各组分的样品,带回实验室进行烘干处理,其中茎、枝和根在105 ℃、叶在75 ℃下烘干至恒重后称取干重。然后根据干鲜重之比计算各组分的生物量。
1.2.4 凋落物及枯落大枝收集
在每个样地内随机设置10个0.5 m2的凋落物收集框。从2011年1月底开始每月收集凋落物,带回实验室对凋落物各组分(落叶、小枝、花果、杂屑等)进行分捡、取样。同时在每块样地内随机设置5个 5 m × 5 m的样方,用于收集枯落大枝(小头直径 ≥ 2 cm),与凋落物同时收集。凋落物在75 ℃下,枯落大枝在105 ℃ 烘干至恒重后称取干重。

表2 西双版纳不同胶林类型各种植物生物量优化回归模型†Table 2 Optimum biomass regression models of different plants in different rubber plantations in Xishuangbanna
各样地的凋落物、枯落大枝及橡胶 + 多种植物组合模式中灌木和草本的生物量均根据单位面积上各自的生物量来推算整个样地内的生物量。
1.3 植物多样性对橡胶树生产力影响的计算方法
运用3种胶林组合模式橡胶树生物量与纯胶林生物量的偏差比,计算各胶林组合模式中其它植物对橡胶树生物量的影响,其计算方法为:

式中:Di为群落i的D值,Oi为i种胶林组合模式橡胶树的生物量,E为纯胶林的生物量。对一特定的胶林组合模式而言,Di>0说明i种胶林群落中的其它植物对橡胶树的生产力具有正的效应,Di=0说明i种胶林群落中的其它植物对橡胶树的生产力没有影响,Di<0则表明橡胶树在i种胶林群落中的生长受到种间竞争的限制甚于种内竞争的限制[28]。
1.4 数据处理
所有数据使用SAS 8.1进行统计分析,运用Microsoft Excel进行数据处理。采用单因素方差分析(ANOVA)和最小显著差异法(LSD)进行显著性检验,显著性水平设定为α=0.05。
2 结果与分析
2.1 不同种植模式橡胶树的生物量及其器官分配
根据各样地调查数据,运用表2中橡胶树的生物量回归模型计算每块样地中橡胶树的生物量(表3),结果显示:在橡胶 + 咖啡模式中橡胶树的生物量高于纯胶林23.45% (p<0.05);而在橡胶 + 茶和橡胶 + 多种植物两种模式中橡胶树的生物量分别高于纯胶林13.47% 和6.09%(p>0.05)。不同组合模式橡胶树各器官的生物量以橡胶 + 咖啡模式最高,橡胶 + 茶模式次之,纯胶林最低。其中橡胶 + 咖啡模式各器官的生物量显著高于纯胶林各器官的生物量(p<0.05)(表3)。在不同模式中,橡胶树相同器官的生物量分配比例变化较小,不到1%,且以茎所占比例最大(67.23%~67.87%),其次是枝(17.81 %~17.92 %),叶所占比例最小(2.18%~2.24%)。表明植物多样性对橡胶树各器官的生物量影响较大,而对各器官生物量的分配比例的影响较小。

表3 西双版纳不同橡胶类型活植物体生物量†Table 3 Biomass allocation of living plants and its various organs in monoculture rubber plantation and different rubbertrees intercrop plantations in Xishuangbanna t·hm-2
2.2 不同组合模式中其它植物的生物量及其器官分配
在橡胶 + 咖啡、橡胶 + 茶和橡胶 + 多种植物模式中(表3),多种植物的生物量是咖啡树和茶树生物量的10.43倍和7.17倍。而各器官的生物量所占比例则不同,多种植物只有茎所占的比例高于茶树茎所占的比例(高19.59%),枝、叶和根所占的比例均小于茶树(分别小7.68%、1.68%和10.24%)。虽然植物体各器官的生物量及其分配随物种组成不同差异较大,但其各器官生物量分配比例的大小顺序均为茎>根>枝>叶(表3)。
在橡胶 + 多种植物模式中,多种植物由其它乔木、灌木和草本3部分组成(表3),其中其它乔木的生物量(45.329 t·hm-2)分别高于灌木43.799 t·hm-2和草本 44.113 t·hm-2。从各器官生物量分配比例方面来看,虽然其它乔木茎和枝所占比例分别是灌木的1.46倍和1.47倍,但根和叶所占比例则比灌木低10.30%和14.64%。其它乔木和灌木各器官生物量的分配比例均以茎最高 (64.43%和44.09%),其次是根(17.36% 和32.00%),叶所占比例最小(3.89% 和14.19%)。而草本植物各器官生物量的分配比例以茎所占的比例最大(38.84%),叶次之(34.07%),根最低(27.09%)。说明橡胶 + 多种植物各组分的生物量及其分配比例因植物种类的不同而存在着较大差异。
2.3 不同种植模式活植物体生物量及其器官分配
不同种植模式中活植物体的总生物量随植物种类的变化而各不相同(表3),以纯胶林的生物量最低(453.227 t·hm-2),而橡胶 + 茶、橡胶+ 咖啡和橡胶 + 多种植物3种模式的活植物体生物量分别是纯胶林的1.15、1.24和1.17倍。橡胶树是各胶林组合模式活植物体生物量的主要组成部分,所占比例达 90.01%~99.18%;其它植物所占比例较低,仅为0.82%~9.09%。活植物体生物量的器官分配同样以纯胶林最低(表3)。在3种胶林复合模式中,茎的生物量以橡胶 + 咖啡模式最高(379.740 t·hm-2),其次是橡胶 + 多种植物(353.279 t·hm-2),橡胶 + 茶最低(350.779 t·hm-2);枝的生物量也以橡胶 + 咖啡模式最高(100.251 t·hm-2),但其次是橡胶 + 茶(93.409 t·hm-2),最低为橡胶+ 多种植物(92.300 t·hm-2);叶的生物量以橡胶+ 多种植物模式最大(13.679 t·hm-2),其次是橡胶 + 茶模式(11.780 t·hm-2),橡胶 + 咖啡模式最低(12.192 t·hm-2);根生物量的变化与其它器官不同,其大小顺序为橡胶 + 多种植物>橡胶 + 咖啡>橡胶 + 茶(表3)。各胶林模式活植物体同一器官生物量所占比例及橡胶树各器官占活植物体总生物量的比例变化较小,不到2%。茶树和橡胶+ 多种植物模式中的其它植物各器官生物量占活植物体总生物量的比例差异相对较大(约5%)。
2.4 不同种植模式死植物体生物量及其组分分配
不同种植模式死植物体总生物量及其各组分的生物量均表现出随植物种类的增加而增加的趋势(表4)。与纯胶林相比,橡胶 + 茶、橡胶 +咖啡、橡胶 + 多种植物3种胶林组合模式死植物体的生物量分别是纯胶林的1.33倍、1.66倍和2.09倍(p<0.05)。在死植物体各组分的生物量组成中,以凋落物的生物量所占比例较大,其中最高的是纯胶林(95.42%),其次是橡胶 + 茶组合模式(95.31%),橡胶 + 多种植物组合模式最低(81.13%),随植物多样性的增加而减少;而枯落大枝的生物量所占比例较小(5.26%~18.87%),并以纯胶林为最低(5.26%),橡胶 +多种植物模式最大(12.94%)(表4),呈现出随植物多样性的增加而增加的趋势,
2.5 不同种植模式的总生物量
不同橡胶种植模式的总生物量随植物种类数量的增加呈现出先增加后降低的趋势(表5),以橡胶 + 咖啡模式的总生物量最大(573.581 t·hm-2),是纯橡胶林的1.25倍(p<0.05),橡胶 + 茶和橡胶 + 多种植物次之,分别是纯胶林的1.15倍和1.18倍 (p>0.05)。在总生物量各组分中,活植物体生物量是各胶林类型总生物量的主要成分,其所占比例高达97.79 %~98.76 %,呈现出随植物多样性的增加而降低的趋势;死植物体的生物量所占的比例较小,仅为1.24 %~2.21 %。但其生物量及所占比例都随植物多样性的增加而增加。
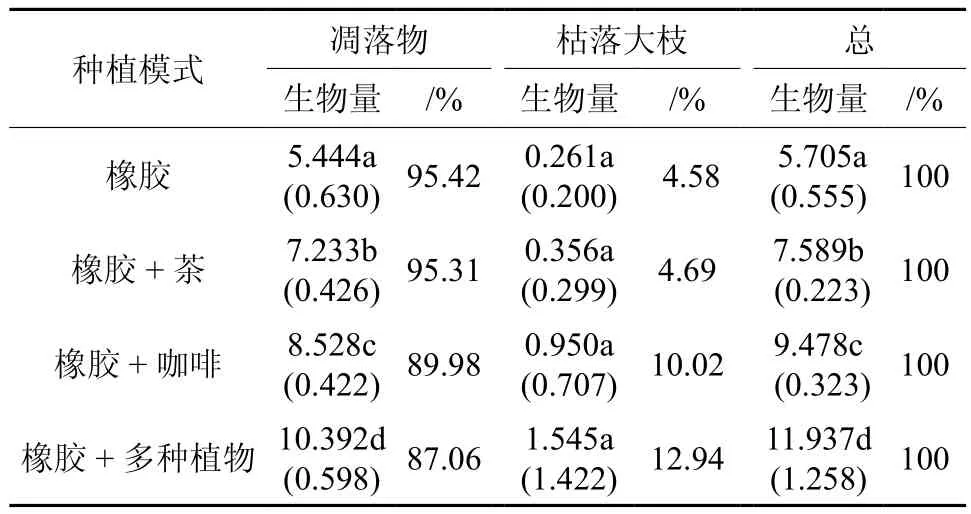
表4 西双版纳不同胶林类型死植物体生物量†Table 4 Biomass of dead plants of monoculture rubber plantation and different mixed rubber-tree plantations in Xishuangbanna t·hm-2

表5 西双版纳不同橡胶林类型的总生物量†Table 5 Total biomass of monoculture rubber plantation and different mixed rubber-trees plantations in Xishuangbanna t·hm-2
2.6 植物多样性对橡胶树生产力的影响
从3种胶林组合模式的橡胶树D值可以看出(图1),橡胶树在3种胶林组合模式中的D值均大于0。说明在互补效应的作用下,橡胶树在各胶林组合模式中比在纯胶林中具有更强的资源利用能力和生长优势,而存在着“超产”现象。但就各胶林组合模式橡胶树的D值而言,其变化较大。在不同的物种水平上,橡胶 + 茶和橡胶 + 咖啡两种模式的橡胶树D值分别是橡胶 + 多种植物的2.21倍和3.85倍。 即使在同一植物多样性水平上,橡胶 + 咖啡模式中橡胶树的D值也是橡胶 +茶模式的1.71倍。表明各胶林组合模式中橡胶树的生产力不仅受胶林群落中物种数量的影响,而且亦受物种组成的影响。

图1 植物多样性对橡胶树生物量的影响Fig.1 Effects of plant diversity on biomass of rubber trees
3 讨 论
3.1 植物多样性对不同胶林类型的生物量的影响
生物多样性与生态系统功能之间的关系是目前生态学领域所研究的核心问题之一[29-30]。虽然许多研究发现生物多样性能够促进群落的生物量生产[31-32],但对其相关作用机制的解释却存在着两种不同的假说,即“生态位互补”和 “抽样效应”[33-34]。其中生态位互补假说认为,不同植物种类组成的群落能够通过物种间的互补效应提高群落的生产力,并且在不同多样性水平间及同一多样性水平内,植物群落生产力变化均较大[17,33,35];抽样效应假说则认为,物种多样性高的群落,其包含更多高产物种的可能性增加,而这些高产物种往往是生态系统功能的贡献者[33-34]。尽管两种作用机制各不相同,但从群落的长期演替过程来看,在群落建立初期,抽样效应是多样性作用的主要机制,而随着时间推移生态位互补的作用会逐渐加强[34]。本研究的结果表明,具有两个物种以上的胶林群落,其生物量均高于纯胶林,显示出植物多样性对胶林群落生产力的正效应关系。但在农林复合生态系统中,物种多样性对群落生产力的作用在很低(5~10种)的时候就能达到饱和[32,36]。因此,胶林群落生产力并不随物种多样性的增加而无限上升。当胶林群落中物种数量比较少的时候,因不同物种的功能地位不同,它们之间的互补性使得胶林生态系统的生产力随植物多样性的增加而增加,因此橡胶 + 茶和橡胶 +咖啡组合模式的生物量高于纯胶林的生物量。但随着植物种类及数量的增加,胶林群落内植物的种内及种间竞争加剧,从而削弱了物种多样性对胶林群落生产力的促进作用,这使物种数量和个体密度较高的橡胶 + 多种植物组合模式(乔木层的物种数量达40种以上,其个体密度为2 086株·hm-2,是纯胶林橡胶 + 茶和橡胶 + 咖啡组合模式的5倍左右)的生物量低于橡胶 + 咖啡组合模式。所以在纯胶林、橡胶 + 咖啡和橡胶 + 多种植物3种组合模式中,胶林群落的生物量随植物多样性的增加,呈现出先增加后降低的单峰曲线模式。而在纯胶林、橡胶 + 茶和橡胶 + 多种植物3种组合模式中,胶林群落的生物量与植物多样性表现出单调上升的关系。这表明,虽然在橡胶 + 多种植物模式中存在着一定程度的竞争作用,但植物多样性对胶林群落生产力的促进作用依然十分明显。
在复合生态系统中物种多样性是影响凋落物的重要因素,与单一的植物群落相比,具有多种植物组成的植物群落,其凋落物的生产量更高[37-38]。而且研究表明,热带地区的人工林由于具有较快的生长速率和较高的生产力,使其产生更多的凋落物[39]。从本研究的结果可知,各胶林类型均具有较高的凋落物生物量,并且呈现出随植物多样性增加而显著增加的正相关关系。另外,物种组成对复合生态系统凋落物生物量也有重要影响[38,40],所以在橡胶 + 茶和橡胶 + 咖啡两种胶林组合模式中,虽然都由两个物种组成,但橡胶 + 咖啡组合模式的凋落物量大于橡胶 + 茶组合模式。此外,枯落大枝的生物量则表现出与凋落物量相类似的变化规律。说明植物多样性对于增加胶林复合生态系统的死植物体生物量具有显著的促进作用。
植物群落中的生态系统过程不仅受物种数量的影响,同时也受每个物种的功能特性及其相互关系的影响[41-42]。由不同物种所形成的群落小环境是影响植物生长发育的一个重要因素,因此物种组成的差异影响着植物群落的生产力[35,43]。尽管一些植物种类在群落中的数量较少,但对生态系统的功能具有十分重要的作用[29]。并且在不同物种组成的群落间,其生产力和互补效应也存在着较大差异,即使在同一物种多样性水平,这种差异也较为明显[44]。在本研究中,各胶林组合模式中橡胶树的相对生物量(D值)显示了不同的结果。在各胶林组合模式中橡胶树的生物量均高于纯胶林橡胶树的生物量,表现出明显的物种间生态位互补特征。但受植物个体间的竞争作用影响,不同胶林群落橡胶树生物量随植物多样性的增加亦呈现出先增加后降低的单峰曲线模式。并且处于同一多样性水平的橡胶 + 茶和橡胶 + 咖啡两种组合模式的D值差异也较大。这说明不仅物种多样性对生态系统的功能具有重要影响,而且物种组成的特性对生态系统生产力、养分循环等过程同样具有较大的影响[45]。由此可见,尽管茶树、咖啡树和多种植物的生物量在各自的胶林组合模式中所占的比例较小,但其对胶林群落的生产力同样具有较大的促进作用。因此,建立多种物种组成的胶林复合生态系统是进行生物多样性保护、提高生物生产力、促进热带退化山地恢复与重建的有效途径。而在胶林复合生态系统的建立过程中,合理的植物物种选择和个体密度不仅有利于植物的生长发育,并且对胶林群落的演替与发展同样具有十分重要的促进作用。
3.2 不同胶林类型的生物量与其它森林类型的比较
对不同森林生物量的比较,将有助于我们更深一步了解各森林类型间在结构和功能上的变化[46]。与世界其它热带地区森林相比,本研究4种不同胶林类型地上部分的生物量在396~492 t·hm-2之间,分别高于南美巴西低地湿润雨林(281 t·hm-2)[47]、非洲刚果热带湿润雨林(374 t·hm-2)[48]和亚洲印尼热带湿性雨林(390 t·hm-2)[49]及柬埔寨热带常绿林(256 t·hm-2)[50]的地上部分生物量;与墨西哥热带漫滩森林 (416 t·hm-2)[51]、利比亚热带湿润雨林(466 t·hm-2)[48]及马来西亚热带雨林 (403 t·hm-2)[52]地上部分生物量相近。4种不同胶林类型的总生物量在458.933~573.581 t·hm-2之间,高于巴西热带原始森林的426 t·hm-2[53],但低于马来西亚龙脑香林的650 t·hm-2[54]。与我国热带地区的森林相比,各胶林类型的总生物量分别高于海南岛山地雨林的总生物量(453 t·hm-2)[55]和西双版纳的热带季节雨林 (424 t·hm-2)[26],是本地区次生林的总生物量(113 t·hm-2)[56]的4~5倍左右;但其变化范围处在该地区热带雨林生物量的变化范围(362~693 t·hm-2)之内[57]。由此可见,具有较大个体密度(360株·hm-2~400株·hm-2)的西双版纳50龄的胶林生态系统在人为抚育管理下,具有较大的生物量积累。
与本地区及海南岛的橡胶林相比,本研究50年生纯橡胶林的生物量(458.933 t·hm-2)是本地区25年生橡胶林生物量(171.112 t·hm-2)[23]的 2.7倍及海南25年生更新橡胶林的生物量(103.6 t·hm-2)[58]的4.4倍。与我国热带和亚热带地区其他人工林的生物量相比,本研究50年生橡胶纯林的生物量是 29年生不同密度的杉木生物量(190~200 t·hm-2)[59]的2.3~2.4倍、闽南35年生(Ⅷ龄级)福建柏人工林生物量(238.34 t·hm-2)[60]的1.93倍、海南岛30年生海南木莲人工林生物量(144.066 t·hm-2)[61]的3.2倍;是广西南宁27年生观光木人工林生物量(101.26 t·hm-2)[62]的4.5倍和17 年生桉树人工林生物量 (112.89 t·hm-2)[63]的 4.1倍。而3个胶林复合生态系统的生物量(528.569~573.581 t·hm-2)是32年生福建柏檫树混交林生物量(222. 32 t·hm-2)[64]的 2.4~ 2.6倍。这表明西双版纳地区的橡胶林具有较高的生物量积累,而胶林复合生态系统则具有更大的生物量积累优势。另一方面也说明胶林复合生态系统具有更大的碳贮存潜力。
[1] 李 博.生态学[M].北京:高等教育出版社,2000 .
[2] 孙儒泳,李庆芬,牛翠娟,等.基础生态学(第二版) [M].北京:高等教育出版社,2002.
[3] Overman J P M,Witte H J L,Saldarriaga J G. Evaluation of regression models for above-ground biomass determination in Amazon rainforest[J]. Journal of Tropical Ecology, 1994, 10(2):207-218.
[4] 刘贤词,文仕知,冯汉华,等.四川桤木人工林不同年龄段生物量的研究[J].中南林业科技大学学报,2007, 27(2):83-86.
[5] Houghton R, Hall F, Goetz S J. Importance of biomass in the global carbon cycle[J]. Journal of Geophysical Research, 2009,114(G00E03):1-13.
[6] Flejzor L. State of the World’s Forests[R]. Rome: Food and Agriculture Organization of the United Nations, 2011,110-119.
[7] Pan Y, Birdsey R A, Fang J, et al. Large and Persistent Carbon Sink in the World’s Forests[DB/MT].www.sciencemag.org/cgi/content/full/science.1201609/DC1.(2011-7-14) [2012-5-10].
[8] Houghton RA. Aboveground forest biomass and the global carbon balance[J]. Global Change Biology, 2005, 11(6): 945-958.
[9] Houghton RA, Goetz SJ. New satellites help quantify carbon sources and sinks[J]. Ecosystems, 2008, 89(43): 417-418.
[10] Brown I F, Martinelli L A, Thomas W W, et al. Uncertainty in the biomass of Amazonian forests: an example from Rondonia,Brazil[J]. Forest Ecology and Management, 1995, 75(1-3): 175-189.
[11] Ridder R M. Global forest resources assessment 2010:Extent of forest resources[R]. Rome: Food and Agriculture Organization of the United Nations, 2007: 33-34.
[12] Li H M, Aide T M, Ma Y X, et al. Demand for rubber is causing the loss of high diversity rain forest in SW China[J]. Biodiversity and Conservation, 2007, 16(6): 1731-1745.
[13] Werner C, Zheng X H, Tang J W, et al. N2O, CH4and CO2emissions from seasonal tropical rainforests and a rubber plantation in Southwest China[J]. Plant and Soil, 2006, 289(1):335-353.
[14] 鲍雅静,李政海,马云花,等.橡胶种植对纳板河流域热带雨林生态系统的影响[J]. 生态环境, 2008,17(2):734-739.
[15] 何云玲,杨根灿.滇南热带季节雨林和橡胶林对降雨侵蚀力的减缓效应[J]. 生态学杂志, 2006, 25(7) : 731-737.
[16] 周再知, 郑海水, 杨曾奖,等. 橡胶-砂仁复合系统生物产量、营养元素空间格局的研究[J]. 生态学报, 1997,17(03): 225-233.
[17] 唐建维,张建候,宋启示,等. 西双版纳热带人工雨林生物量及净第一性生产力的研究[J]. 应用生态学报, 2003, 14(001):1-6.
[18] Montagnini F, Nair PKR. Carbon sequestration:an underexploited environmental benefit of agroforestry systems[J]. Agroforestry Systems, 2004, 61: 281-295
[19] 孟庆岩,王兆骞,余盛兴. 我国热带地区胶—茶—鸡农林复合模式社会经济效益分析[J]. 中国人口资源与环境, 2001,11(52): 44-46.
[20] 沈守艮. 西双版纳两个海拔胶―茶群落结构和生产力研究[D]. 西双版纳:中国科学院西双版纳热带植物园,2007.
[21] 宋清海,张一平.西双版纳地区人工橡胶林生物量、固碳现状及潜力[J]. 生态学杂志, 2010, 29(10): 1887-1891.
[22] 贾开心,郑 征,张一平.西双版纳橡胶林生物量随海拔梯度的变化[J]. 生态学杂志, 2006, 25(9): 1028-1032.
[23] 唐建维,庞家平,陈明勇,等. 西双版纳橡胶林的生物量及其模型[J]. 生态学杂志, 2009, 28(10): 1942-1948.
[24] 王丽娟. 不同遮荫树种和多样性变化对茶园土壤、茶叶品质及净生产量的影响[D]. 西双版纳:中国科学院西双版纳热带植物园, 2011.
[25] Suárez-Pascua D. Cuantificación y valoración económica del servicio ambiental almacenamiento de carbono en sistemas agroforestales de café en la Comarca Yassica Sur, Matagalpa,Nicaragua[D]. CATIE thesis. Costa Rica: Turrialba, 2002.
[26] 吕晓涛,唐建维,何有才,等.西双版纳热带季节雨林的生物量及其分配特征[J]. 植物生态学报, 2007, 31(001): 11-22.
[27] 巩合德, 程希平, 马月伟. 云南杨梅灌丛生物量的分配特征[J]. 经济林研究, 2012, 30(4): 106-108.
[28] Loreau M. Separating sampling and other effects in biodiversity experiments[J].Oikos, 1998, 82(3): 600-602.
[29] Loreau M, Naeem S, Inchausti P, et al. Biodiversity and ecosystem functioning: current knowledge and future challenges[J]. Science, 2001, 294(5543):804-808.
[30] 邓立斌, 刘德晶. 基于层次分析法的尖峰岭自然保护区生态评价[J]. 中南林业科技大学学报, 2010, 30(7):26-31.
[31] Pfisterer A B, Schmid B. Diversity-dependent production can decrease the stability of ecosystem functioning[J]. Nature, 2002,416(6876):84-86.
[32] 江小雷,张卫国,严 林.植物群落物种多样性对生态系统生产力的影响[J]. 草业学报, 2004,13(6):8-13.
[33] Tilman D, Reich P B, Knops J, et al. Diversity and productivity in a long-term grassland experiment[J]. Science, 2001, 294(5543):843-845.
[34] 张全国, 张大勇. 生物多样性与生态系统功能:最新的进展与动向[J].生物多样性, 2003, 11(005): 351-363.
[35] 江小雷, 张卫国. 一年生植物群落内取样效应和互补效应的分离[J]. 生态学杂志, 2006, 26(6): 1896-1902.
[36] (美)蔡 平,(美)马特森,(美)穆尼著. 陆地生态系统生态学原理[M]. 李 博, 赵 斌, 彭荣豪, 等译.北京:高等教育出版社, 2005.
[37] Wang Q, Wang S, Fan B, et al. Litter production, leaf litter decomposition and nutrient return in Cunninghamia lanceolata plantations in south China: effect of planting conifers with broadleaved species[J]. Plant and Soil, 2007, 297 : 201–211.
[38] Scherer-Lorenzen M, Bonilla J L, Potvin C. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment[J]. Oikos, 2007, OE:1-17.
[39] Lugo A E. Comparison of tropical tree plantations with secondary forests of similar age[J].Ecological monographs,1992,62(1):1-41.[40] Tang J W, Cao M, Zhang J H, et al. Litterfall production,decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: a 10-year study[J].Plant and Soil, 2010, 335:1-18.
[41] Cabido M. Vive la Différence: plant functional diversity matters to ecosystem processes[J]. Trends in Ecology & Evolution, 2001,16(11):646-655.
[42] 彭方仁, 王良桂. 不同密度林分的生长发育与生物生产力. 经济林研究[J], 1998,16(3): 12-16.
[43] 张一平,刘玉洪,马友鑫.热带森林不同生长时期的小气候特征[J]. 南京林业大学学报(自然科学版), 2002,26(001):83-87.
[44] 江小雷,张卫国,段争虎.资源互补效应对多样性-生产力关系的影响[J].植物生态学报,2005, 29(4):523-529.
[45] Tilman D, Lehman C L, Thomson K T. Plant diversity and ecosystem productivity: theoretical considerations[J].Proceedings of the National Academy of Sciences, 1997, 94(5):1857-1861.
[46] Cairns M A, Olmsted I, Granados J, et al. Composition and aboveground tree biomass of a dry semi-evergreen forest on Mexico’s Yucatan Peninsula[J]. Forest Ecology and Management,2003, 186:125-132.
[47] Chave J, Condit R, Lao S, et al. Spatial and temporal variation of biomass in a tropical forest: results from a large census plot in Panama[J]. Journal of Ecology, 2003, 91(2): 240-252.
[48] Brown S, Gaston G. Use of forest inventories and geographic information systems to estimate biomass density of tropical forests: application to tropical Africa[J]. Environmental Monitoring and Assessment, 1995, 38(2): 157-168.
[49] Noordwijk M V, Rahayu S, Hairiah K, et al. Carbon stock assessment for a forest-to-coffee conversion landscape in Sumber-Jaya (Lampung, Indonesia): from allometric equations to land use change analysis[J]. Science In China (Series C), 2002,45:75-86.
[50] Top N, Mizoue N, Kai S. Estimating forest biomass increment based on permanent sample plots in relation to woodfuel consumption: a case study in Kampong Thom Province,Cambodia[J]. Journal of Forest Research, 2004, 9(2): 117-123.
[51] Jaramillo V J, Kauffman J B, Rentería-Rodríguez L, et al.Biomass, carbon, and nitrogen pools in Mexican tropical dry forest landscapes[J]. Ecosystems, 2003, 6(7):609-629.
[52] Hoshizaki K, Niiyama K, Kimura K, et al. Temporal and spatial variation of forest biomass in relation to stand dynamics in a mature, lowland tropical rainforest,Malaysia[J]. Ecological Research, 2004, 19(3): 357-363.
[53] Brown S, Lugo A E. Aboveground biomass estimates for tropical moist forests of the Brazilian Amazon[J]. Interciencia Caracas,1992, 17(1): 8-18.
[54] Proctor J, Anderson J M, Chai P, et al. Ecological studies in four contrasting lowland rain forests in Gunung Mulu National Park,Sarawak: I. Forest environment, structure and fl oristics[J].The Journal of Ecology ,1983,71(1): 237-260.
[55] 陈德祥,李意德,Liu H,等.尖峰岭热带山地雨林生物量及碳库动态[J].中国科学:生命科学,2010, 40(007):596-609.
[56] 唐建维,张建侯,宋启示,等.西双版纳热带次生林净初级生产量的初步研究[J].植物生态学报, 2003,27(006):756-763.
[57] Zheng Z, Feng Z, Cao M, Li Z, et al. Forest structure and biomass of a tropical seasonal rain forest in Xishuangbanna,Southwest China[J]. Biotropica, 2006, 38(3): 318-327.
[58] 周再知,郑海水,尹光天,等. 橡胶树生物量估测的数学模型[J]. 林业科学研究, 1995, 8(6): 624-629.
[59] 林开敏,俞新荽,何智英,等.不同密度杉木林分生物量结构与土壤肥力差异研究[J].福建林业科技, 1996,32(5):385-391.[60] 魏影景.闽南福建柏人工林生物量结构的研究[J]. 福建林业科技, 2001,3(3): 21-23.
[61] 周光益,曾庆波,林明献,等. 海南木莲人工林生物量及养分分配[J]. 林业科学研究, 1997,10(5): 453-457.
[62] 黄松殿,吴庆标,廖克波,等. 观光木人工林生态系统碳储量及其分布格局[J].生态学杂志,2011,30(11): 2400-2404.
[63] 温远光,陈 放,刘世荣,等.广西按树人工林物种多样性与生物量关系[J]. 林业科学, 2008, 44(4): 14-19.
[64] 邹双全. 福建柏檫树混交林生物量及分布格局研究[J]. 福建林学院学报, 1998, 18(1): 40-43.
Biomass and its allocation pattern of monoculture and mixed rubber-tree plantations in Xishuangbanna
XIAO Zi-wei1,2, WANG Xiu-hua3, ZHENG Li4, WANG Xiao-li4, GAO Li-hong5, TANG Jian-wei1
(1.Open Lab. of Tropical Plant Resource Science, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666303, Yunnan, China; 2. Graduate University of Chinese Academy of Science, Beijing 100049, China; 3. Xishuangbanna Station of Soil Fertilizer, Jinghong 666100, Yunnan, China; 4. Yunnan Academy of Sci-tech Information, Kunming 650051, Yunnan,China; 5. Xishuangbanna Institute of Sci-tech Information, Jinghong 666100, Yunnan, China)
In order to quantify the biomass of monoculture rubber plantation and mixed rubber-tree plantations, three plots with a size of 20 m × 25 m were established in each of four different rubber plantations, i.e., a monoculture rubber plantation and three mixed rubber-tree plantations, respectively, in Xishuangbanna. The constructed biomass regression models of different tree species were used to estimated their biomasses. The biomasses of monoculture rubber plantations were 69.636 t·hm-2, 114.648 t·hm-2and 81.924 t·hm-2lower than that of mixed Hevea brasiliensis + Camellia sinensis, Hevea brasiliensis + Coffea liberica and Hevea brasiliensis + multi plants plantations, respectively. The biomass of living plants occupied 97.79%~98.76% of total biomass in four different rubber plantations,and dead plants only took up 1.24%~2.21% . The results indicate that the plant species richness had positive effects on rubber biomass and enhanced bio-productivity of integrated rubber-tree plantations.
rubber plantation; biomass; agro-forestry system; Xishuangbanna of Yunnan province
S794.1
A
1673-923X(2014)02-0108-09
2013-06-10
中国科学院农业项目办公室项目(KSCX2-YW-N-44-09);西双版纳州科学技术局项目(200915)
萧自位(1986-),男,云南龙陵人,硕士,主要从事农林复合生态系统碳储量研究;E-mail:xzw464999488@126.com
唐建维(1964-),男,广西全州人,博士,研究员,主要从事热带森林生态系统与农林复合生态系统的研究工作;
E-mail:tangjw@xtbg.org.cn
[本文编校:文凤鸣]
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
热带生物学报(2022年1期)2022-03-09
农业科技与信息(2021年9期)2021-12-07
农业科技与信息(2021年8期)2021-12-06
云南画报(2021年8期)2021-12-02
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
中国野生植物资源(2019年2期)2019-06-11
世界热带农业信息(2016年6期)2016-07-02
云南画报(2015年10期)2015-01-09
