
择伐干扰对长白山阔叶林中水曲柳及其伴生树种分布格局及种间相关性的影响
2014-12-22 03:08阚彬彬王庆成吴文娟东北林业大学林学院黑龙江哈尔滨150040
安徽农业科学 2014年35期
阚彬彬,王庆成*,吴文娟(东北林业大学林学院,黑龙江哈尔滨150040)
研究森林群落优势种群结构和空间分布格局对探讨森林生态系统的原始性、可持续性、群落稳定性、演替规律以及物种的生态学及更新特征等非常重要[1],探讨种群分布格局的目的不仅是对种群的水平结构进行定量描述,更重要的是揭示格局的成因,阐述种群及群落的动态变化等,为种群保护措施的制定提供一定的科学依据[2]。种群的空间分布格局不但因种而异,而且同一种在不同发育阶段、不同的生境条件也有明显的差别[3]。种群在受到不同强度人为干扰时,由于生境条件变化程度及各种群恢复进程不同,不同生物学特性的种群空间分布格局也产生差异[4],人类采伐干扰是作用于生态系统的一种外力,它使生态系统的结构发生改变,使生态系统动态过程偏离其自然的演变方向和速度。在干扰的影响下,森林的生态适应对策因物种类型的差异而不同,进而影响植物对有限资源的竞争,决定了种群不同尺度的空间格局[5-7]。
目前关于植物群落分布格局的研究很多,但关于人为干扰对种群分布格局的影响研究相对较少。周建云[8]研究了不同间伐强度下辽东栎种群结构特征和空间分布格局,原民龙[9]运用更新采伐方式对华北落叶松人工林空间格局展开研究,杨梅[4]针对人为干扰对常绿阔叶林主要种群分布格局的影响展开过探讨,周蔚[10]研究了择伐强度对长白山天然云冷杉针阔混交林空间结构的影响,郑丽凤[11]研究了择伐强度对天然针阔混交林更新格局的影响。当前关于采伐干扰对水曲柳(Fraxinus mandshurica Rupr.)的分布格局及其种间关系的研究很少。水曲柳为东北“三大硬阔”之一,是东北主要的珍贵树种,由于其较好的用材价值,使其曾遭到毁灭性的采伐。20世纪50~80年代,长白山地区阔叶林中的有用材价值的水曲柳也曾遭到大面积破坏,只有小面积由于运输、采伐不便才得以保持原始状态,经过国家30多年的封山育林,阔叶林中的水曲柳逐渐进入恢复状态。该文从水曲柳的生态学角度出发,研究采伐干扰对于水曲柳自身的生长及其伴生树种的影响,为水曲柳的保护提供理论依据。
1 材料与方法
1.1 研究区域概况 该研究样地选择在长白山露水河林业局,该局位于长白山西北麓,地理坐标为127°29'~128°02'E,42°20'~42°40'N,全局平均海拔600 ~800 m,海拔1 000 m 以上区域很少,总坡向为西北向。气候为温带大陆性气候,冬夏风向更替明显。由于气温低,蒸发量小,空气湿润,这些因素的综合有利于形成茂密的落叶阔叶与针叶混交林。长白山露水河林业局在20世纪50~80年代曾经开展过大量采伐活动,并且采伐范围大,采伐木材种类多,对原始林中珍贵树种破坏严重,但是经过近30年时间的封山育林,林业整体得到恢复。
1.2 野外调查和数据收集 2012年通过长时间的林地踏查,同时结合当地林业局历史资料,选取立地条件相似、经过不同强度择伐干扰的水曲柳阔叶林为研究对象。选取东升林场21林班2小班为水曲柳未经择伐干扰样地(样地1),此样地作为对照,水曲柳轻度择伐干扰样地选择在东升林场31林班2小班上(样地2),水曲柳重度择伐干扰样地选择在东升林场57林班2小班上(样地3),样地2和样地3中的水曲柳在20世纪70年代经过不同强度的择伐,水曲柳择伐强度分别为15%和30%。林中其他树种破坏较小。
每块样地大小为100 m×100 m,用罗盘将整个样地划分为25个20 m×20 m的小样方,每隔20 m设一基点,以20 m×20 m样方为单位,将样地内的所有胸径(DBH)>1 cm的乔木用铝合金号牌编号,并记录种名、胸径、树高、冠幅(东西冠幅长、南北冠幅长)及其所处坐标,之后转换为样地内的坐标。
1.3 研究方法 点格局及种间关系都采用以Ripley’K函数为基础的方法进行研究(Ripley,1981),Ripley’K函数由Ripley率先提出[12],该函数将每个个体视为二维空间的一个点,以全部个体组成的二维点图为基础进行格局分析,可以进行多尺度空间格局以及空间关联分析,能更好地反映种群空间特征,故广泛应用于植物种群的空间格局分析中。Besag在Ripley’K函数的基础上进行了优化,提出了用Ripley’K函数的变形—L(t)函数[13]。该研究采用L(t)函数进行数据分析,过程采用Programit软件(2010版)完成,空间尺度r范围为0~50 m,步长1 m,利用Monte-Carlo方法通过100次计算得到由2条上下包迹线围成的98%置信区间。在分布格局研究中若函数曲线在上包迹线上为聚集分布,在下包迹线下为均匀分布,在上下包迹线之间为随机分布;在种间关系研究当中,使用由Ripley’s K12(t)函数变形而来的L12(t)函数确定双变量点格局间的空间关系,如果函数曲线在上包迹线上为二者正关联,在下包迹线下为二者负关联,在2条包迹线间为二者不关联[14]。
2 结果与分析
2.1 样地树木组成分析 选取样地中的水曲柳及其主要伴生树种——椴树(Tilia amurensis Rupr.)、红松(Pinus koraiensis Sieb.et Zucc.)、色木槭(Acer mono Maxim.)、榆树(Ulmus pumila L.)为研究对象。样地1中水曲柳共89棵,平均胸径45.84 cm,其胸高断面积占群落总胸高断面积43%,整块样地中水曲柳位于主林层,演替层中的水曲柳少有,更新层中水曲柳在距离主林层水曲柳一定范围内偶有生长,林分整体保持原始状态。样地2因曾经择伐原因,主林层中水曲柳有所减少,但水曲柳仍旧占据着主林层,演替层中和更新层中也有分布,在更新层存在部分幼苗。样地3中水曲柳曾经遭到重度择伐,水曲柳在主林层中分布较少,红松占据部分主林层,水曲柳绝大部分分布于演替层和更新层。样地中相关树种数据见表1。

表1 3块样地中水曲柳及主要伴生树种胸高断面积、胸径及株数
2.2 水曲柳及伴生树种空间格局分析 研究发现在样地1和样地2中水曲柳均为随机分布(图1 a-1,a-2),但样地1中的水曲柳在大于30 m尺度上有聚集分布的趋势,因为样地1中的水曲柳保持原始状态,经过种间和种内竞争,处于一个相对稳定的群落中,大部分水曲柳处于主林层,而当物种处于主林层时分布格局通常都是随机分布,虽然样地2中的水曲柳经过轻度择伐,但对于水曲柳群落总体来讲破坏不大,因此依然保持随机分布格局。样地3中主林层中的水曲柳经过重度择伐,形成了大小不等的林窗,形成的林窗有利于处于更新层和演替层水曲柳的生长。韩有志等[15]人研究发现林分光照环境在水曲柳更新中的作用十分关键,林分光照的空间异质性对更新格局起决定性作用。样地3中的水曲柳处于更新层和演替层,因此其分布格局趋于聚集分布,这也是更新层树种的分布格局特点。
椴树在样地1中0~7 m聚集分布,大于7 m随机分布(图1 b-1),在样地2中0~5 m聚集分布,5~43 m随机分布,大于43 m均匀分布(图1 b-2),在样地3中0~27 m随机分布,大于27 m聚集分布(图1 b-3)。椴树在样地1和样地2中多处于演替层,但在椴树附近有小的更新苗存在,因此小尺度上会出现聚集分布格局。样地3中在主林层破坏后有局部地方存在密度相对较大的椴树种群,通过实地勘察,与试验结果相符合。
红松在样地1和样地2中均为随机分布(图1 c-1,c-2),样地3中的红松0~39 m呈现聚集分布,大于39 m随机分布(图1 c-3)。红松在样地1和样地2中大部分处于演替层,生长相对稳定,但有占据主林层的生长趋势。更新层中的红松很少,几乎没有。这样的生长状况容易形成随机分布格局。样地3中由于主林层中的水曲柳择伐严重,形成很多的林隙空地,这为样地中的红松聚集生长提供了空间。择伐干扰与自然干扰下形成的林隙不同,影响着种群的结构及动态过程,从而形成不同的格局分布[16]。这些林窗使地温增加,枯落物分解速度加快,某些喜光的树种加速生长导致种群聚集性增强。
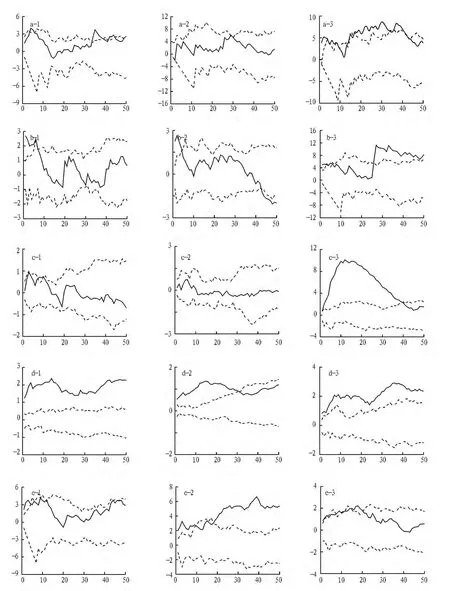
图1 5个树种在不同样地中空间分布格局
色木槭在样地1和样地3中聚集分布(图1 d-1,d-3),样地2中0~30 m尺度上聚集分布,大于30 m尺度上随机分布(图1 d-2)。色木槭在3块样地中分布较广、数量大,大多处于更新层和演替层,因此在样地1和经过重度择伐的样地3中形成了聚集分布格局,在样地2中大尺度上有部分色木槭生长到演替层,因此在大尺度上形成了一定随机分布格局。
榆树在样地1和样地3中随机分布(图1 e-1,e-3),样地2中的榆树在10~18 m尺度上随机分布,其他尺度上聚集分布(图1 e-2)。榆树在3块样地中大部分处于演替层,经过种间和种内竞争数量相对较少,因此随机分布。
2.3 水曲柳与伴生树种空间关联性分析 在空间关联性研究当中发现水曲柳与椴树在样地1和样地2中总体表现为不关联(图2 f-1,f-2),在样地3中0~33 m不关联,大于33 m正关联(图2 f-3)。因为样地1和样地2中水曲柳主要处于主林层,而椴树处于演替层,二者相对处于不同层次,群落相对稳定,种间竞争较小,因此二者表现为种间不关联,样地3中的椴树为聚集生长的水曲柳更新苗提供了很好的环境庇护,如避免强光照射等,因此二者在大尺度上表现为正相关。水曲柳与红松在样地1和样地2中不关联(图2 g-1,g-2),在样地3中0~5 m不关联,5~47 m负关联(图2 g-3)。样地1和样地2中的群落是经过种间竞争相对稳定的植物群落,虽然水曲柳处于主林层,而且红松也有向主林层生长的趋势,但由于种群生长相对稳定,因此二者种间表现为不关联。样地3中主林层水曲柳的破坏虽然为林下更新层聚集生长的红松提供了生长空间,但是红松幼苗阶段是喜阴生长,因此水曲柳的砍伐不利于红松幼苗的生长。

图2 水曲柳与伴生树种在不同样地中空间关联性
水曲柳与色木槭在样地1中0~23 m不关联,大于23 m正关联(图2 h-1);在样地2中二者不关联(图2 h-2),样地3中二者正关联(图2 h-3)。在样地1中群落在自然生长状态下相对稳定,水曲柳处于主林层,色木槭大部分处于更新层,二者相互影响小。在样地2中,因为部分水曲柳的择伐,造成主林层中的水曲柳与演替层和更新层中的色木槭生长距离相对较远,二者空间影响较小,因此表现为不关联。样地3中水曲柳的破坏为阔叶树色木槭生长提供了空间,因为样地3中二者表现为正相关。
水曲柳与榆树在样地1中0~30 m不关联,大于30 m二者正关联(图2 i-1),样地2中13~15 m,20~34 m二者正关联,其他尺度不关联(图2 i-2),样地3中0~35 m二者不关联,大于35 m负关联(图2 i-3)。
3 结论与讨论
种群空间格局是指种群个体在水平空间的配置状况或分布状态,也是种群生物学特性、种内种间关系及环境条件综合作用的结果,还是种群基本数量特征之一[17-18]。不同的空间格局类型和空间关联性可以反映出种群利用环境资源的状况,揭示种群生物学内涵,是其在群落中地位与生存能力的外在表现[19-20]。在原始状态下,未经过择伐干扰以水曲柳为主林层的阔叶林,水曲柳呈现随机分布状态,符合物种在较低的林层中呈明显的聚集分布,而在较高的林层则呈随机或者规则性的分布格局,聚集度也随林层的增高而降低的一般规律[21]。但是轻度择伐对于水曲柳与椴树、红松的种间关系改变并不明显,对于水曲柳与榆树和色木槭种间关系有所改变,在未经择伐干扰的水曲柳样地中,水曲柳与榆树和色木槭种间关系都表现在小尺度上不相关,中尺度和大尺度上的正相关。重度择伐对于水曲柳与椴树、红松、色木槭、榆树的种间关系有重要影响。
该研究得出结论:轻度择伐主林层中的水曲柳有助于改善种群结构,但对于处于更新层中的伴生阔叶树种如榆树、色木槭有一定的影响,对于水曲柳与椴树、红松种间关系改变不明显。重度择伐阔叶林中的水曲柳有助于阔叶树种椴树的生长,但不利于处于更新层中针叶树种红松的生长。在原始林中对于水曲柳与色木槭、榆树在大尺度上二者正关联这一现象还有待进一步研究。
[1]刘足根,朱教君,袁小兰,等.辽东山区次生林主要树种种群结构和格局[J].北京林业大学学报,2007,29(1):12 -17.
[2]张兴旺,张小平,郝朝运,等.安徽琅琊山青檀种群结构与空间分布格局研究[J].广西植物,2011,31(6):782 -788.
[3]康华靖,刘鹏,陈子林,等.不同生境香果树种群的径级结构与分布格局[J].林业科学,2007,43(12):22 -27.
[4]杨梅,林思祖,曹光球,等.人为干扰对常绿阔叶林主要种群分布格局的影响[J].中国生态农业学报,2007,15(1):9 -11.
[5]陈宝瑞,杨桂霞,张宏斌,等,不同干扰类型下羊草种群的空间格局[J].生态学报,2010,30(21):5868 -5874.
[6]CONDIT R,ASHTON P S,BAKER P,et al.Spatial patterns in the distribution of tropical tree species[J].Science,2000,288(5470):1414 -1418.
[7]HE F,DUNCAN R P.Density-dependent effects on tree survival in an old- growth douglas fir forest[J].Journal of Ecology,2000,88(4):676 -688.
[8]周建云,李荣,张文辉,等.不同间伐强度下辽东栎种群结构特征与空间分布格局[J].林业科学,2012,48(4):149 -155.
[9]原民龙,陈金龙,崔丽红,等.更新采伐对华北落叶松人工林空间格局的影响[J].河北林果研究,2012,27(3):255 -259.
[10]周蔚,杨华,亢新刚,等.择伐强度对长白山区天然云冷杉针阔混交林空间结构的影响[J].西北林学院学报,2012,27(4):7 -12.
[11]郑丽凤,周新年,罗积长,等.择伐强度对天然针阔混交林更新格局的影响[J].福建林学院学报,2008,28(4):310 -313.
[12]RIPLEY B D.Spectral analysis and the analysis of pattern in plant communities[J].Journal of Ecology,1978,66(3):965 -981.
[13]BESAG J.Contribution to the discussion of Dr Ripley’s paper[J].Journal of the Royal Statistical Society,1977,B39:193 -195.
[14]张春雨,赵秀海,王新怡,等.长白山自然保护区红松阔叶林空间格局研究[J].北京林业大学学报,2006(S2):52-58.
[15]韩有志,王政权,谷加存.林分光照空间异质性对水曲柳更新的影响[J].植物生态学报,2004,28(4):468 -475.
[16]魏媛,喻理飞.不同干扰条件下构树萌枝种群结构和分布格局[J].福建林学院学报,2009,29(4):362 -367.
[17]吴承祯,洪伟,吴继林,等.珍稀濒危植物长苞铁杉的分布格局[J].植物资源与环境学报,2000,9(1):31 -34.
[18]张金屯.植被数量生态学方法[M].北京:中国科学技术出版社,1995:36-49.
[19]DALE M R T.Spatial pattern analysis in plant ecology[M].Cambridge:Cambridge University Press,1999:20 -22.
[20]张金屯,孟东平.芦芽山华北落叶松林不同龄级立木的点格局分析[J].生态学报,2004,24(1):35 -40.
[21]张健,郝占庆,姚晓琳,等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J].应用生态学报,2007,18(8):1681 -1687.
猜你喜欢
植物研究(2023年5期)2023-09-09
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
林业科技情报(2021年1期)2021-04-14
森林工程(2018年4期)2018-08-04
现代营销(创富信息版)(2016年6期)2016-11-21
现代营销(创富信息版)(2016年12期)2016-02-08
杂草学报(2015年2期)2016-01-04
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
