
珠江口盆地东部中新统珠江组碳酸盐岩沉积微相研究
2014-12-19 07:16周小康
地质力学学报 2014年1期
冯 琳,邓 敏,卫 哲,曾 驿,周小康
(1.成都理工大学沉积地质研究院,成都610059;2.油气藏地质及开发工程国家重点实验室,成都理工大学,成都610059;3.中海石油 (中国)有限公司深圳分公司研究院,广州510240)
0 引言
碳酸盐岩沉积微相的研究已经从最初依据岩石薄片鉴定单纯进行岩类学和古生物学描述[1]发展到今天综合用于岩石学特征、古生物特征描述、碳酸盐岩分类命名、沉积环境解释、成岩演化历史分析等。碳酸盐岩沉积微相研究的迅猛发展无疑得益于偏光显微镜、阴极发光、流体包裹体、扫描电镜和稳定同位素等分析测试技术的进步,但是,一个重要的物质基础是能够获得足够的样品供研究者开展这些研究。珠江口盆地东部中早中新世珠江期以碳酸盐岩沉积为主,生物礁、滩沉积发育,是重要的油气富集层位。中海石油 (中国)有限公司深圳分公司在对东沙隆起珠江组碳酸盐岩进行油气勘探的过程中,钻探了21口钻井,采集了670 m的岩心,磨制了1483块薄片,这为本文开展碳酸盐岩沉积微相研究提供了物质基础。
前人对珠江口盆地东部东沙隆起中新统珠江组碳酸盐岩的研究主要集中在生物礁及储层特征与成因[1~4]和沉积相与层序地层[5~8]两个方面,鲜有关于沉积微相研究的报道,制约了对珠江组沉积环境的客观科学认识,同时也是对这些来自海上的宝贵的井下岩心资料的浪费,因为这些钻井岩心中所包含的矿物学、岩石学和古生物学信息给人们提供了认知珠江组沉积环境、储层特征等方面的丰富线索。因此,本文在对珠江口盆地东部15口钻井岩心观察和1483块薄片鉴定的基础上,充分考虑Dunham(1962)、Embry和Klovan(1971)、曾允孚 (1970)等各家碳酸盐岩分类命名方案的优缺点,提出了适合珠江组碳酸盐岩沉积发育特点的碳酸盐岩分类命名方案,并据此分类方案,描述珠江组碳酸盐岩的微相类型和组合特征,进而详细分析珠江组的沉积环境,建立珠江组的沉积模式,为南海珠江组碳酸盐岩的油气勘探和沉积演化研究提供地质依据。
1 研究区地质背景
珠江口盆地东沙隆起位于南海北部大陆架南缘,构造位置为中央隆起带东段,呈北东向展布 (见图1)。早中新世珠江期碳酸盐台地主要位于盆地东部东沙隆起上,呈北东走向。台地内不同区域的碳酸盐岩厚度存在明显差异,台地西南缘流花地区有一环礁带,称为流花礁带,沉积时间较长,厚度最大,向北逐渐减薄。20世纪80年代至今,已经在流花地区、陆丰地区发现多个生物礁油气田。
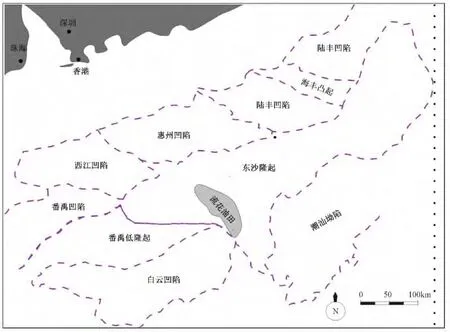
图1 珠江口盆地地理位置及构造区划Fig.1 Geographical location and tectonic map of Pearl River Mouth Basin
笔者在研究东沙隆起带珠江组碳酸盐岩时,着重对下部灰色生屑粒泥岩夹白云质灰岩、中部生屑泥粒岩夹生屑粒泥岩以及上部灰白色生物骨架岩、粘结岩和生物碎屑粒泥-泥粒岩的微相特征进行了较为系统的研究和分析,试图通过微相和其组合特征,对珠江组碳酸盐岩的沉积发育史有更深入的认识和了解。
2 地层特征
东沙隆起新近系下中新统珠江组发育良好,自下而上可分为底部砂岩段,中部灰岩段,上部砂泥岩段 (见表1),与珠海组呈整合接触,并与下伏珠海—恩平组、文昌组,上覆韩江组在纵向上形成一套良好的生、储、盖组合。本文研究位于珠江组中部的碳酸盐岩,最厚562 m,以生物碎屑粒泥-泥粒岩、生物骨架岩和粘结岩为主,局部发生白云岩化。流花地区珠江组主要发育灰岩,惠州及陆丰地区仅在珠江组下部发育灰岩。

表1 珠江组及其相邻地层概况 (据文献[9]修改)Table 1 Stratigraphic profiles about Zhujiang Formation and the adjacent strata
3 微相类型及特征
3.1 颗粒组分
颗粒和基质类型及其特征是划分碳酸盐岩微相类型、分析沉积环境的重要依据[10]。珠江组灰岩的颗粒类型主要为生物碎屑,以及少量的内碎屑。
①生物碎屑:主要为珊瑚藻、有孔虫,其次还有红藻石[11]、珊瑚、海绵、腕足、腹足、双壳、棘皮类和少量的介形虫等。
②内碎屑:这种颗粒类型不如生物碎屑常见,为盆地内弱固结的碳酸盐灰泥经波浪或流水打碎,搬运再沉积下来的碎屑颗粒,多为砂屑,含少量砾屑。
3.2 微相类型划分
本文在综合前人碳酸盐岩分类命名方案优缺点的基础之上,建立了适宜于珠江组灰岩的命名方案。生物成因灰岩及生物灰岩主要参考Duham(1962)、Embry等 (1971)的方案,同时利用曾允孚 (1970)方案对颗粒类型进行细化的优势,在划分珠江组的颗粒-灰泥石灰岩时,将其按颗粒类型以及相对含量的多少进行细分。微相的划分则以Flügel(2004)的标准微相类型为依据,根据颗粒和基质类型、沉积组构等微相资料将珠江组的碳酸盐岩划分为22个微相 (见图2)。
3.2.1 灰泥岩/MF1
该微相为基质支撑,基质全为泥晶方解石,偶见少量生物,如抱球虫、介形虫,含量一般少于10%,或见海绿石附生组合 (见图2a),该类岩石粒度很细,孔隙基本不发育。主要形成于局限台地或潮坪的低能环境。
3.2.2 粒泥岩/MF2
按生物颗粒类型以及相对含量的多少,又可以划分为4个亚类。
3.2.2.1 生屑粒泥岩/MF2-1
这种微相类型在台地中并不多见,生物碎屑极破碎,大多数碎屑原始组构已难以辨别,多为珊瑚、海绵、双壳、棘皮类等的碎片 (见图2b)。为高能带生物颗粒经强烈的波浪或沿岸流作用打碎,后经波浪或潮汐搬运,沉积于浪基面以下的低能环境,局部见少量白云石化作用。
3.2.2.2 珊瑚藻屑粒泥岩/MF2-2
碎屑颗粒主要为珊瑚藻的碎片,种属包括古石枝藻Archaeolithothamnium属、石叶藻Lithophyllum属、珊瑚藻Jania属[12],横切面呈圆形,纵切面呈长条状,分枝或不分枝 (见图2c)。镜下可见丝体规则排列,少数可见被亮晶充填的孢子囊和生殖窝。除藻屑外还有一些含量很少的有孔虫、腕足、腹足类等生物的碎片,总量小于10%。破碎的珊瑚藻为高能带的珊瑚藻骨架经强烈的波浪作用打碎,后经波浪或潮汐搬运,沉积于浪基面以下开阔台地的低能环境。
3.2.2.3 红藻石粒泥岩/MF2-3
颗粒主要为红藻石,基质为微亮晶方解石,红藻石粒泥岩中的红藻石一般粒径较小,为珊瑚藻呈纹层状包卷生长形成,发育特征的精细网格组织及孢子囊 (见图2d)。这类岩相多出现于台地前斜坡,或者是台地边缘构成礁顶或礁基的红藻石滩中。
3.2.2.4 含海绿石抱球虫粒泥岩/MF2-4
主要颗粒类型为海绿石及浮游有孔虫,并含少量粒度较细的珊瑚藻屑、介形虫等的化石(见图2e),填隙物为泥微晶方解石。该类岩相很少出现,海绿石及广海浮游生物的组合,指示该岩相可能来自于局限台地的风暴沉积。
3.2.3 泥粒岩/MF3
泥粒岩是研究区整个碳酸盐台地中分布最广泛的一种微相类型,颗粒类型丰富,大小和形态各异。按照颗粒类型的差异以及相对含量的多少,可以划分为9个亚类 (见图2f—2n)。
3.2.3.1 有孔虫藻屑泥粒岩/MF3-1
生物颗粒主要为藻屑,有孔虫次之,藻屑颗粒较破碎,粒度不一,有孔虫则保存完好,粒度相当,多为1 mm左右,或还含有少量的腹足、棘皮及双壳类碎片;基质主要为泥微晶方解石,局部发生弱白云化。这种微相类型在碳酸盐岩台地中较为常见。

图2 珠江组碳酸盐岩微相类型Fig.2 Carbonate microfacies types in Zhujiang Formation
3.2.3.2 藻屑有孔虫泥粒岩/MF3-2
碎屑颗粒的含量以有孔虫相对较多,藻屑次之,常具定向性,还含有很少量的苔藓虫碎片以及棘屑、珊瑚砾屑、腹足碎片,偶见海绵。生物碎屑多保存完好,藻屑则较破碎,粒径较小,部分相带发育的这类岩相的颗粒边界模糊不清,泥晶化严重。基质为亮晶方解石胶结物。根据上述特征,推测这类岩相可能形成于水动力条件很强的台地边缘或台地边缘后浅滩环境。
3.2.3.3 生屑珊瑚藻屑泥粒岩/MF3-3
生物颗粒除了藻屑颗粒外,还含有类型丰富的极度破碎的生物化石颗粒,颗粒原始结构已难以辨别,可能为珊瑚、海绵、有孔虫、腹足等的碎屑颗粒,颗粒的高度破碎反映其为异地沉积,这类微相形成于台地内能量相对较低的环境。
3.2.3.4 藻屑泥粒岩/MF3-4
生物颗粒主要为藻屑颗粒,其他类型的生物颗粒含量很少 (小于10%)。藻类主要为红藻门珊瑚藻科。藻碎屑一般呈长条状,亦有未完全打碎,体积较大的藻团块。此种亚类型并不在泥粒岩微相中占主要地位。
3.2.3.5 有孔虫泥粒岩/MF3-5
生物颗粒为保存完好的有孔虫,同时还有含量很少的藻类及苔藓虫的碎片。有孔虫粒度不一,以底栖有孔虫为主,有孔虫泥粒岩多出现于开阔台地内水相对较深的低能环境。
3.2.3.6 红藻石泥粒岩/MF3-6
同MF2-3一样,红藻石泥粒岩中红藻石一般为小红藻石,不参与造礁,多出现于构成礁顶或礁基的红藻石滩相中。
3.2.3.7 苔藓虫泥粒岩/MF3-7
生物颗粒主要为苔藓虫,还含有少量其他生物碎屑,这种微相中苔藓虫未呈大规模出现,并未参与造礁。
3.2.3.8 珊瑚屑粒泥岩/MF3-8
常与珊瑚礁伴生,颗粒主要为极其破碎的珊瑚碎屑,以及少量的海绵、有孔虫及棘皮动物的碎片,碎屑颗粒分选较差,基质主要为泥晶方解石。这类微相并不常见,主要出现于礁的骨架孔中,包含较多灰泥基质。
3.2.3.9 砂屑泥粒岩/MF3-9
这种岩相为富含内碎屑的石灰岩,其碎屑粒度不一,粒径多为砂屑级,原始组构已难以识别,多见于水动力较强的台前斜坡碎屑流和浅水台地沉积中,其中有的碎屑在搬运前已完全胶结,但仍可将其作为盆地内的准同生再造物质,即认为它们为内碎屑。
在泥粒岩分类当中,以MF3-1、MF3-2、MF3-3等3类微相占整个碳酸盐岩台地岩相的主要部分,也是珠江组灰岩段最为常见的岩石类型。
3.2.4 颗粒灰岩/MF4
主要为有孔虫珊瑚藻屑颗粒岩 (见图2o),该岩石类型在台地较少出现,只间或出现在流花地区珠江组灰岩取心段的下部,颗粒支撑,含泥很少,颗粒主要为珊瑚藻屑、有孔虫,粒间孔很发育,形成于水动力很强的台地边缘环境。
3.2.5 骨架岩/MF5
按主要造礁生物类型可以将骨架岩划分为2种类型 (见图2p—2q)。
3.2.5.1 珊瑚骨架岩/MF5-1
珊瑚是珠江组最为常见的造礁生物,以枝状六射珊瑚居多,类型包括孔珊瑚、柱珊瑚、鹿角珊瑚、山星珊瑚、近星珊瑚、星孔珊瑚等,为原地生长的具抗浪骨架的碳酸盐建隆。除珊瑚外,参与造礁的生物还有珊瑚藻、海绵、大底栖有孔虫、苔藓虫等,居礁生物主要为有孔虫、腕足、腹足、双壳、棘皮类等。波浪和潮汐共同作用的高能环境适宜于珊瑚的生长,因此珊瑚骨架岩沉积多形成于台地边缘或开阔台地内水动力很强的环境当中,构成台地边缘礁或开阔台地内点礁。
3.2.5.2 珊瑚藻骨架岩/MF5-2
造礁生物主要为珊瑚藻,在岩心和薄片中很容易见到,多为保存完好的藻体,也有藻屑。岩心上常呈皮壳状和结核状,薄片中可见到分枝状、皮壳状。皮壳状珊瑚藻粘结生长,而结核状珊瑚藻呈缠绕状,为原地生长具抗浪骨架的生物,构成珊瑚藻骨架礁,多发育于台地边缘水动力较强的礁核亚相。
3.2.6 粘结岩/MF6
3.2.6.1 红藻石粘结岩/MF6-1
这类微相主要岩性为红藻石粘结岩,红藻石主要是由非固着的珊瑚藻构成的独立钙质结核。珠江组具粘结结构的红藻石普遍是由珊瑚藻和结壳有孔虫或苔藓虫互层包卷生长形成的,具粘结结构的红藻石多呈球状、椭球状或包壳状 (见图2r),与滩相的红藻石相比,体积更大,参与造礁。其形态的多样性反应了沉积环境的差异性,其中球形的、椭球形的和纹层状的生长模式代表高能的沉积环境,要求水体能量高促使其频繁滚动。
在珠江组灰岩的取心段中,红藻石粘结岩非常发育并具有一定的规模。其中相当数量的红藻石颗粒都较大,其直径最大者可达15 cm,一般2~6 cm。较大的红藻石颗粒具有较强的抗浪能力,对灰泥起着遮挡和捕集作用,其粘结和包裹着大量灰泥基质,常构成礁顶或礁基的包壳粘结岩,称之为粘结礁。这种藻类的包壳颗粒形成于中等高能,极浅水环境。
3.2.6.2 苔藓虫粘结岩/MF6-2
为片状—板状苔藓虫粘结,包裹着各种居礁生物和大量灰泥基质 (见图2s),不具备抗浪骨架。陆丰地区,苔藓虫粘结岩呈厚层状广泛发育,或为苔藓虫层状礁。该微相并不是台地的主要生物礁类型。
3.2.7 障积岩/MF7
沉积物中含有丰富的茎状或树枝状生物化石遗体,多为异地搬运沉积下来的生物化石,如珊瑚、海绵、海百合茎等 (见图2t),通过障集作用,捕集灰泥,在其后堆积成丘,基质在含量上相对较多,抗浪能力差,一般出现在低能环境。
3.2.8 白云岩/MF8
白云岩在珠江组分布较少,主要出现于珠江组灰岩取心段的下端,常与灰岩共存。白云岩中残留部分原岩的结构和组分,可以推测这类白云岩为次生白云岩。按照白云石相对含量的多少和晶粒的大小,可以划分为2个亚类。
3.2.8.1 灰质白云岩/MF8-1
多为细中晶颗粒,半自形—自形晶,偶见陆源石英混入,仍可见原岩结构和组分,其原岩多为生物碎屑泥粒岩 (见图2u)。原岩残余组构中广盐度生物普遍出现,推测此微相可能形成于间或暴露的碳酸盐缓坡或潮坪环境。
3.2.8.2 细粉晶白云岩/MF8-2
白云石含量90%以上,半自形晶,呈紧密镶嵌式接触 (见图2v)。偶见陆源石英混入,以及少量生物碎屑。
次生白云岩对沉积环境的识别有一定的指导意义,根据研究区的构造背景、岩石学以及沉积特征,判断其原岩可能为灰泥岩或生物碎屑粒泥岩,该微相或形成于水浅暴露的环境。
4 沉积模式及沉积相分析
4.1 碳酸盐岩缓坡模式 (Ⅰ期)
该模式出现在碳酸盐岩台地沉积之前,因珠江组取心灰岩段下部普遍有白云石化现象和广盐度生物同时出现 (见图3),推测碳酸盐岩台地形成之前其沉积环境应为碳酸盐缓坡。缓坡低能带水动力较弱,主要沉积泥晶碳酸盐沉积物,高能带则沉积颗粒较粗的碳酸盐颗粒,主要为生物碎屑泥质颗粒岩-颗粒质泥岩以及一些灰泥岩 (见图4),生物种类繁盛,数量较多,并因水浅暴露,常见次生的白云质灰岩、灰质白云岩、细粉晶白云岩发育。
4.2 镶边碳酸盐岩台地模式 (Ⅱ期)
根据前述的微相分析,并结合剖面结构的沉积特征,在综合前人碳酸盐岩相模式的基础上,将该碳酸盐岩台地划分为台前斜坡相、台地边缘礁相、台地边缘滩相、开阔台地相、局限台地相、台地潮坪相6种相带类型。
4.2.1 台地前斜坡相
除了台地前垮塌角砾沉积,台前斜坡上还发育着生屑滩、红藻石滩、点礁或塔礁,含广海生物。
4.2.2 台地边缘礁
高能带,生物礁具有抗浪骨架,能粘结生物、捕集灰泥。岩性主要为珊瑚藻骨架岩和珊瑚骨架岩,有孔虫,苔藓虫,海绵,绿藻也参与造礁,红藻石滩常构成礁顶和礁基。本区构成造礁格架或骨架的生物主要是珊瑚和珊瑚藻,其次是苔藓虫。居礁生物主要为有孔虫、腹足、双壳、棘皮类等。生物礁向海一侧为高能带,波浪作用强烈,适宜于具抗浪骨架的珊瑚的生长,形成以块状珊瑚为主的骨架岩沉积;向陆一侧,水体能量减弱,各种生物都较发育,珊瑚藻及腕足、腹足等的含量增加,于岩心上常见生物扰动等沉积构造。生物礁因其孔隙发育,常为储层分布和发育的有利相带。
4.2.3 台地边缘滩
台地边缘浅滩相主要为生屑滩,岩性为泥质颗粒灰岩和颗粒岩,基质多为亮晶胶结,代表其水动力极强的高能环境。该环境由于受到波浪和潮汐的共同作用,能量很高,灰泥含量一般很少,当水体能量交替变化或暴露交替时,可形成生物礁和滩的互层。
4.2.4 开阔台地
开阔台地可划分为台地内浅滩和滩间亚相。浅滩为生屑滩,岩性主要为生物碎屑泥粒灰岩,为台地内水动力较强的环境;而滩间主要为生物碎屑粒泥岩,灰泥基质含量相对于生屑滩升高,测井曲线上表现为GR的值升高,代表台地内相对较弱的水动力环境。
4.2.5 局限台地相
主要为位于礁、滩或障壁后的澙湖,其岩性主要为生物碎屑粒泥灰岩及灰泥岩。因水循环受限,生物种类有限,澙湖底以软体动物、绿藻、有孔虫为主,生物扰动强烈,向岸过渡为潮坪。

图3 惠州Y井珠江组沉积相综合柱状图Fig.3 Comprehensive column of sedimentary facies in Zhujiang Formation about Well Y in Huizhou area
4.2.6 台地潮坪相
出现于珠江组灰岩取心段的下部,该相带可进一步划分为潮间带和潮下带。潮间带主要沉积的岩石为灰泥岩、生物碎屑粒泥灰岩,生物碎屑颗粒细小,一般具定向性,还可见到一些灰质白云岩或细粉晶白云岩,常有藻席发育;由于潮汐水流方向反复变换,羽状交错层理发育。潮间带的动物群种类有限,仅腹足类比较丰富,伴有介形虫、有孔虫及双壳类。潮下带几乎很少出露水面,相对于潮间带,灰泥含量稍减少,水动力条件略高于潮间带环境,岩石类型与潮间带基本相同,多见生物扰动构造。
通过上述沉积相类型以及沉积模式的分析,可见珠江组灰岩的发育经历了由碳酸盐岩缓坡到台地的沉积过程。

图4 珠江口盆地碳酸盐岩沉积模式图及各微相类型在沉积相序中的分布[13~14]Fig.4 Sedimentary model of Pearl River Mouth Basin and distributions of all types of microfacies in the sedimentary facies sequence
5 结论
珠江口盆地东部珠江组碳酸盐岩的主要颗粒类型为生物碎屑 (包括藻类、有孔虫、珊瑚、苔藓虫、海绵、腕足、腹足、双壳、棘皮、介形虫等)和少量内碎屑。岩性则以生物碎屑泥粒-粒泥灰岩和骨架岩为主,并发育具有相当规模的红藻石粘结岩。
根据珠江组灰岩的生物组合,颗粒、基质类型及特征将该地区的碳酸盐岩划分为22种微相类型,分属于台前斜坡相、台地边缘礁相、台地边缘滩相、开阔台地相、局限台地相、台地潮坪相以及缓坡相。沉积模式及沉积相带组合表明,在早中新世该区在海平面上升的过程中碳酸盐岩的发育经历了由碳酸盐缓坡—镶边碳酸盐台地的演化过程。
[1] Flügel E.石灰岩微相[M].曾允孚,译.北京:地质出版社,1989.Flügel E.Limestone microfacies[M].ZENG Yun-fu,Translated.Beijing:Geological Publishing House,1989.
[2] 付振群,傅恒,汪瑞良,等.珠江口盆地东沙隆起珠江组储层特征及主控因素[J].油气地质与采收率,2013,20(4):10~16.FU Zhen-qun,FU Heng,WANG Rui-liang,et al.Reservoir characteristics and main controlling factors of Zhujiang Formation in Dongsha Uplift,Pearl River Mouth Basin[J].Petroleum Geology and Recovery Efficiency,2013,20(4):10~16.
[3] 古莉,胡光义,罗文生,等.珠江口盆地流花油田新近系生物礁灰岩储层特征及成因分析 [J].地学前缘,2012,19(2):49~58.GU li,HU Guang-yi,LUO Wen-sheng,et al.Characteristics and genesis of reservoir spaces in Neocene reef reservoir of Liuhua oilfield,Perrl River Mouth basin[J].Earth Science Frontiers,2012,19(2):49 ~58.
[4] 曾驿,汪瑞良,刘军,等.东沙隆起碳酸盐岩储层特征及预测[J].石油天然气学报,2011,33(6):69~73.ZENG Yi,WANG Rui-liang,LIU Jun,et al.Carbonate reservoir characteristics and prediction of Dongsha Uplift[J].Journal of Oil and Gas Technology,2011,33(6):69~73.
[5] 胡诚,郑荣才,戴朝成.珠江口盆地珠江组流花生物礁及储层特征[J].岩性油气藏,2010,9(3):59~64.HU Cheng,ZHENG Rong-cai,DAI Zhao-cheng.Liuhua organic reef and reservoir characteristics of Zhujiang Formation in Pearl River Mouth Basin[J].Lithologic Reserivoirs,2010,9(3):59 ~64.
[6] 黄诚,傅恒,汪瑞良.珠江口盆地 (东部)新近系珠江组碳酸盐岩沉积相及沉积模式[J].海洋地质前沿,2011,9(9):18~25.HUANG Cheng,FU Heng,WANG Rui-liang.Sedimentary facies and depositional model of the Neogene Zhujiang Formation in the Pearl River Basin(East)[J].Marine Geology Frontiers,2011,9(9):18~25.
[7] 周小康,汪瑞良,曾驿,等.珠江口盆地东沙隆起珠江组碳酸盐岩层序地层及沉积模式 [J].石油天然气学报,2011,33(9):1~6.ZHOU Xiao-kang,WANG Rui-liang,ZENG Yi,et al.Sequence stratigraphy and depositional model of Zhujiang Formation in Dongsha Uplift,Pearl River Mouth Basin[J].Journal of Oil and Gas Technology,2011,33(9):1 ~6.
[8] 黄诚,傅恒,汪瑞良,等.珠江口盆地东沙隆起珠江组层序界面的识别及划分[J].海相油气地质,2010,15(3):32~39.HUANG Cheng,FU Heng,WANG Rui-liang,et al.Identification and division of Tertiary Zhujiang sequence boundaries in Dongsha Uplift,Pearl River Mouth Basin[J].Marine Origin Petroleum Geology,2010,15(3):32 ~39.
[9] 李胜勇.珠江口盆地东部中新统珠江组碳酸盐岩与生物礁层序地层研究[D].成都:成都理工大学,2011.LI Sheng-yong.The sequence stratigraphy about the carbonate platform and organic reef of the Miocene Zhujiang Formation in Pearl River Mouth Basin[D].Chengdu:Chengdu University of Technology,2011.
[10] 徐锦龙,贾志海,王伟.四川江油—广元地区上石炭统碳酸盐岩微相及其沉积环境分析[J].沉积学报,2012,10(5):834~846.XU Jin-long,JIA Zhi-hai,WANG Wei.Microfacies and environment analysis of the Carboniferous carbonate in Jiangyou-Guangyuan area,Sichuan Province[J].Acta Sedimentologica Sinica,2012,10(5):834 ~846.
[11] 刘丽,郭荣涛,吴和源,等.红藻石和蓝藻石概念及分类归属的综述和修订[J].地球科学与环境学报,2011,33(2):137~141.LIU Li,GUO Rong-tao,WU He-yuan,et al.Review and modification of definition and classification of Rhodoid and Cyanoid[J].Journal of Earth Sciences and Environment,2011,33(2):137 ~141.
[12] 余素玉.化石碳酸盐岩[M].北京:地质出版社,1989:1~137.YU Su-yu.Fossil carbonate[M].Beijing:Geological Publishing House,1989:1~137.
[13] Flügel E.Microfacies of Carbonate Rocks[M].New York:Springer-verlag,Berlin Heidelberg,2004:1 ~924.
[14] Flügel E.碳酸盐岩微相——分析、解释及应用 [M].马永生,译.北京:地质出版社,2006.Flügel E.Carbonate microfacies:Analysis,interpretation and application [M].MA Yong-sheng,Translated.Beijing:Geological Publishing House,2006.
猜你喜欢
海洋地质与第四纪地质(2022年6期)2023-01-10
阅读(快乐英语高年级)(2022年3期)2022-03-30
环境与发展(2020年7期)2020-08-14
科学Fans(2019年2期)2019-04-11
沉积与特提斯地质(2018年2期)2018-11-22
伴侣(2018年2期)2018-02-28
普洱(2016年9期)2017-05-20
普洱(2016年9期)2017-05-20
录井工程(2017年4期)2017-03-16
浙江海洋大学学报(自然科学版)(2017年6期)2017-03-08
