
per2基因参与边缘系统昼夜节律的研究进展
2014-11-27 07:19:16朱毅郭佳宝李凝岳雨珊张文毅谢斌
中国康复理论与实践 2014年2期
朱毅,郭佳宝,李凝,岳雨珊,张文毅,谢斌
生物钟在细胞的新陈代谢、增殖等过程中起着至关重要的作用,它们参与一系列的昼夜调节转录翻译的反馈、前馈回路[1],其特异性核心组成是生物钟基因及其蛋白产物。生物钟基因通过直接或间接的方式控制DNA的表达,例如损伤的识别和修复[2],其蛋白产物也可以通过调节下游基因的转录来调节其表达[3]。
生物钟基因包括clock、bmal1、per-1,2,3、cry-1,2、reverbα和dec-1,2,主要分布在中枢神经系统脑和脊髓以及周围各个器官,如肠道、胃、心脏、肝脏、肾脏、胰腺、睾丸、卵巢、骨骼肌、口腔黏膜、皮肤、骨骼和单细胞白血球等[4-5],不仅参与生物周期节律的调节,还参与器官功能的调节,当其突变或者缺失时,会引起节律系统的紊乱。bmal1/clock家族主要分布于视交叉上核(suprachiasmatic nucleus,SCN)、杏仁核(amygdala)、下丘脑腹侧核(ventromedial hypothalamus,VMH)、嗅球(olfactory bulb,OB)、外侧系带(lateral habenula)、海马(hippocampus)和下丘脑(hypothalamus)内的一些核团[6-8],在对内脏活动和情绪的调控中,边缘系统起到重要作用。per2基因是bmal1/clock家族一个成员,参与一系列的正负反馈环路,在边缘系统对情绪和内脏活动的昼夜节律调节中扮演重要的角色[9],除了受到SCN调节,还接受上级边缘系统(如终纹床核、杏仁核)和下游皮质醇的负反馈调节,而其在边缘系统的作用以及边缘系统内各功能区之前的联系尚不清楚。本文主要针对per2基因形态学、生物学以及其参与边缘系统对情绪、内脏活动昼夜节律调节机理做一综述。
1 per2基因形态学概述
per2主要是由二聚体PAS-A和PAS-B通过1,4-二巯基苏糖醇(1,4-dithiothreitol)连接形成PAS-AB二聚体,PAS-AB二聚体和后面的α螺旋体结构(α-helical extension,αF)构成per2的中央片段[10]。per2基因首先在果蝇中被发现,随后根据同源性分析,在哺乳动物中也发现表达该基因,小鼠为mper2,人类为hper2,两者均含有封闭的PAS结构域[11]。人类的per2基因与果蝇和小鼠的per2基因分别具有44%和77%的同源系类[12]。斑马鱼的clock1a、clock1b是哺乳动物clock1的同源基因[13]。果蝇的clock基因具有惊人的序列保守性,与哺乳动物的节律基因clock有高度的相似性,他们都含有基础螺旋-环-螺旋结构和PAS区域,以及一个转录活化区域[14]。
per2与CLOCK/BMAL1复合物生物功能密切相关。bmal1和clock均为碱性螺旋-环-螺旋(basic helix-loop-helix,bHLH)转录因子的C组家族成员[15]。动物bHLH转录因子中包含A、B、C、D、E、F 6组共45个家族[10,16],最早发现于小鼠体内,是最早被确定的哺乳动物钟基因;bHLH转录因子几乎存在于所有的真核生物体内,在生长发育中起到极其重要的调控作用。clock基因是哺乳动物生物钟节律的重要调控者,在需要时钟调控的组织中起着中心作用。见图1。
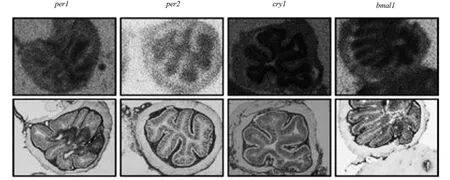
图1 钟基因在老鼠结肠上皮的表达[17]
2 per2基因的生物功能
2.1 per2的合成和储存
由两个转录因子编码的蛋白,含有bHLH-PAS结构域,然后以异二聚体的形式结合到per2上游的E盒,从而激活per2基因的转录,其表达产物为PER2。然后per2基因与另外一种钟蛋白组成异二聚体,发挥抑制作用。在哺乳动物中,转录因子CLOCK和BMAL1通过bHLH-PAS结构域形成异二聚体,二聚化的CLOCK-BMAL1结合到per1-3和cry1-2上游启动子的E盒,从而激活基因转录,进而翻译产生PER和CRY蛋白。PER蛋白与CRY蛋白在细胞质内形成异二聚体,该二聚体入核后通过与CLOCK/BMAL1二聚体结合,抑制CLOCK/BMAL1二聚体与E盒的结合,从而呈现生物钟的负反馈环路调节[18]。
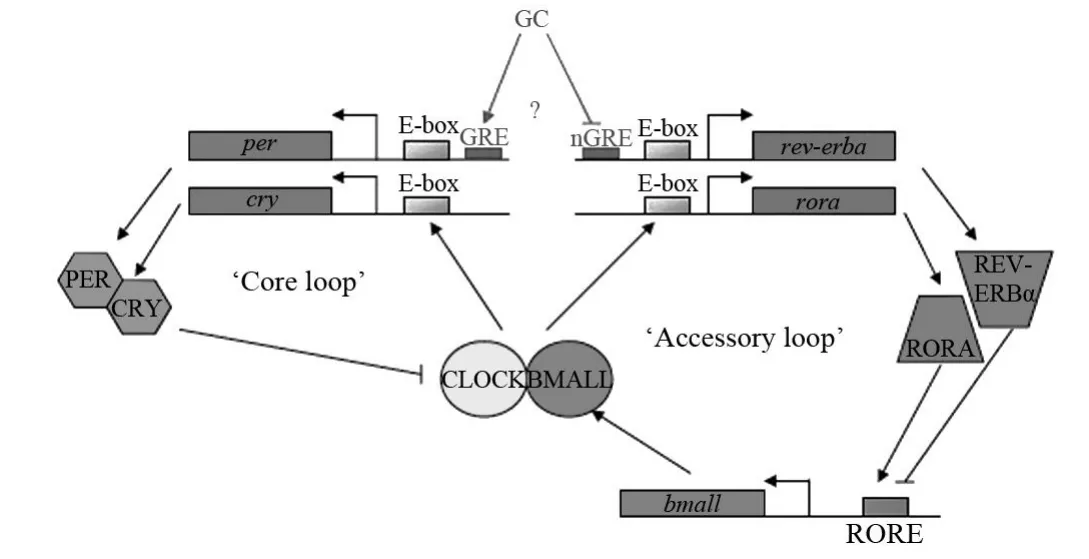
图2 钟基因生物节律反馈路径[19-22]
2.2 per2的释放和失活
细胞外信号激活下游的一些蛋白激酶,然后激活胞内环磷酸腺苷(cAMP)和细胞分裂素活化蛋白激酶(MAPK),使PER2蛋白磷酸化,继而转录激活因子,从而使其与per2基因的启动因子结合,诱导per2的激活[13]。而其失活的主要原因是PER2蛋白磷酸化[23]:PER2蛋白生成后,在胞浆中累积,在酪蛋白激酶的作用下不断磷酸化,磷酸化状态下的PER2蛋白不稳定,可被广泛生物素化而降解[11]。
3 per2参与边缘系统调节对情绪和内脏运动的影响
在下丘脑以上的中枢神经系统中,己明确边缘系统通过调节下丘脑-垂体-肾上腺(hypothalamus-pituitary-adrenal,HPA)轴的活动而参与对情绪和内脏应激反应的调节。边缘系统-下丘脑-垂体-肾上腺(limbic system-HPA,LHPA)轴被称为是连接中枢神经系统和机体周围组织的“信息高速公路”[24]。per2参与边缘系统的昼夜节律表达。per2基因主要表达在终纹床核(bed nucleusof thestria terminalis,BNST)、中央杏仁核(central nucleus of amygdala,CEA)、海马齿状回(dentate gyrus,DG)、基底外侧杏仁核(basolateral amygadaloid,BLA)等脑内核团[25-26],且均为应激时神经内分泌参与运动、情感、行为和心理调节的重要区域,提示per2在边缘系统的作用可能涉及应激和药物滥用相关的行为和神经内分泌反应,摄食、学习和记忆的调节,母性行为和生殖等领域,涉及多个靶器官[19,27]。per2在维持边缘系统功能的完整性上发挥重要作用[11],所以per2也可能通过参与边缘系统的昼夜节律的调节来参与情绪和肠道活动[28]。见图3。
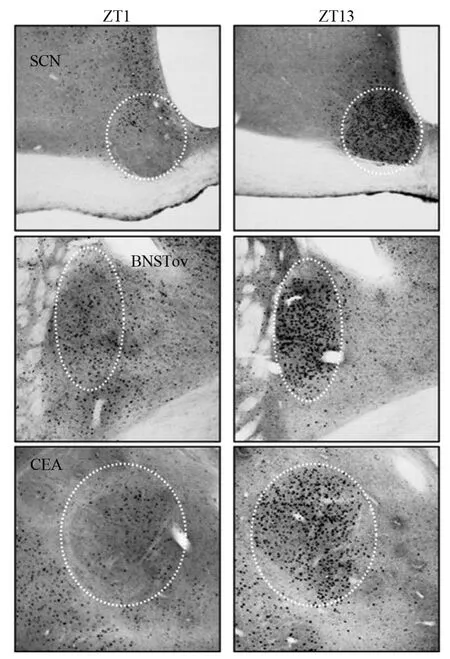
图3 在ZT1或ZT13,per2在大鼠的SCN、BNSTov和CEA的免疫反应性
3.1 SCN对per2在边缘系统作用的同步调控
生物钟可分为母钟和子钟。哺乳动物的母钟位于下丘脑前部的SCN,它被认为是主要的昼夜节律的起搏器,控制着机体行为和生理[31]。在哺乳动物中,光信号能直接启动生物的节律。外界环境的光信号进入眼睛,直接作用于光敏感视黄醇神经细胞,通过视网膜下丘脑纤维,最后到达SCN,诱导per2基因的转录,进而翻译产生PER2蛋白[13],在边缘系统内PER2蛋白的节律表达受到SCN的控制,SCN中的PER2蛋白的节律能同步调控在边缘系统中的PER2蛋白的振荡器,进而调控PER2蛋白在边缘系统作用。在BNST、CEA、BLA和DG 4个区域内,我们发现了PER2蛋白节律的表达呈现2个相反的模式,在BNST和CEA上,PER2蛋白表达的最高点大约在白天转变为夜晚的这个过渡时间段内,且他们的节律与在SCN内的PER2节律同步,相反,在BLA和DG内,PER2节律的峰值在早上,并与SCN、BNST、CEA的节律相反[11]。
3.2 PER2蛋白参与BNST和CEA的节律调节
Alheid和Heimer提出,终纹床核(bed nucleus of stria terminalis,BST)是杏仁核的延伸部,因为它们与杏仁核有类似的下行投射,还有证据表明它们由苍白嵴发育而成。这两个结构在神经发生过程中是同一来源,所以结构、功能相似,但是他们参与的通路有些差异[32]。CEA和BNST参与激活HPA轴的应激反应,下丘脑室旁核(hypothalamic paraventricular nucleus,PVN)是HPA轴活动的直接控制部位,CEA可能直接作用于PVN,也可能通过BNST等中继核团信息的传递来作用于PVN[24]。HPA轴的激活能促进类固醇激素的分泌,而在CEA和BNST上PER2的节律对这些激素的改变表现敏感。如抑制下丘脑室旁核的肾上腺皮质激素释放因子(corticotropin releasing factor,CRF)表达,阻碍CRF进入BNST,从而减少PER2蛋白的水平,CRF参与PER2蛋白表达取决于细胞内信号而不是直接作用于per2基因的转录。见图4。
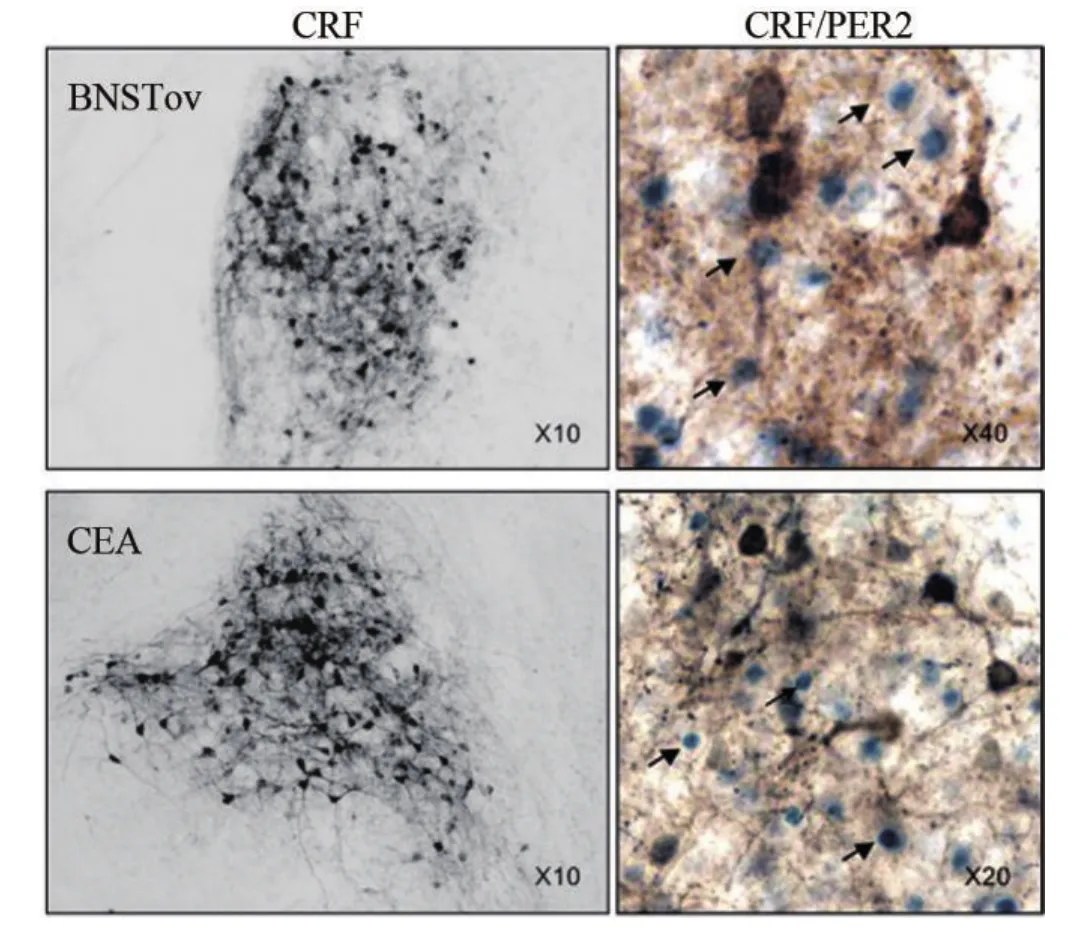
图4 CRF和CRF/PER2在BNSTov和CEA的表达
另外,以皮质醇(corticosterone,CS)影响为主的糖皮质激素(glucocorticoid,GC)发出信号,由在BNST和CEA上的激素受体介导,通过调节per2基因在BNST和CEA内的表达而产生相应生物学效应。甲状腺激素(thyroid hormone)通过调节per2基因的转录来影响PER2蛋白的表达[11];应激负荷时HPA轴分泌释放的激素可以调节消化道的功能,如促甲状腺激素释放激素(thyroliberin,TRH)可使小肠及大肠运动亢进,而促肾上腺皮质激素释放激素(corticotrophin-releasing hormone,CRH)则减低胃的运动但使大肠运动亢进[33]。所以,当外界一个刺激作用于机体时,激活CEA和BNST上的神经元,通过兴奋HPA轴分泌释放激素来参与情绪和内脏应激的调控。HPA轴分泌的激素又能产生相应的反馈作用,调节在CEA和BNST上PER2蛋白的表达水平,导致per2基因在CEA和BNST上的节律受到影响,从而影响CEA和BNST的功能。所以per2基因参与CEA和BNST的节律调节,是情绪和消化道中枢调控的途径之一。见图5。

图5 生物钟系统和HPA轴在多个层次活动的相互作用[34]
3.3 PER2蛋白参与BLA和DG的节律调节
作为HPA轴的高位调节中枢,大量研究表明海马能抑制HPA轴的激活[1]。海马齿状回是海马的传入门户[35],当一个外界刺激作用时,神经元兴奋累计到一定数量时引起整个DG的兴奋,DG和BLA的兴奋对HPA轴的激活起到抑制作用,而PER2蛋白在BLA和DG上的节律表达对BLA和DG的功能产生影响,从而通过影响HPA轴来参与情绪和内脏的调控。
4 PER2和HPA轴对肠道节律性运动的调节及临床意义
肠道的运动也遵循一定的昼夜节律,它的运动通常发生在白天,尤其在醒后,很少发生的夜间,通过改变生活作息来打破这种正常的肠道节律会导致一系列的肠道症状,这说明肠道运动与生物钟有关[35],而失眠同样也是钟基因异常的表现[36-37]。运用免疫组织化学的研究方法,在对神经递质合成和肠功能调节具有重要作用的肠上皮细胞(intestinal epithelial cells,CE)、肠肌丛(myenteric plexus,MP)和肌间神经丛标本(longitudinal muscle myenteric plexus,LMMP)中发现了PER2蛋白,说明其在肠道的细胞和神经元内普遍存在[3]。有研究表明,破坏per1和per2基因后发现小鼠的肠道运动减弱,而单独破坏per1和per2基因的老鼠在肠道运动的表达也产生了时间差异[38],所以per2及其编码的蛋白参与了肠道的运动,在肠道生理活动上扮演重要的角色[3,19]。
而PVN不仅通过节律性释放CRF调节肠道节律运动,还通过激活HPA轴和脑内边缘系统CRF神经元完成应激反应,调节自主神经系统兴奋性,来影响肠道运动,使HPA轴促肾上腺皮质激素(adrenocorticotrophic hormone,ACTH)水平增加,节律性减弱,结肠组织皮质醇(cortisol)水平升高[39]。由此可见per2基因与HPA轴的相互作用在肠道功能性疾病的研究有重要意义。
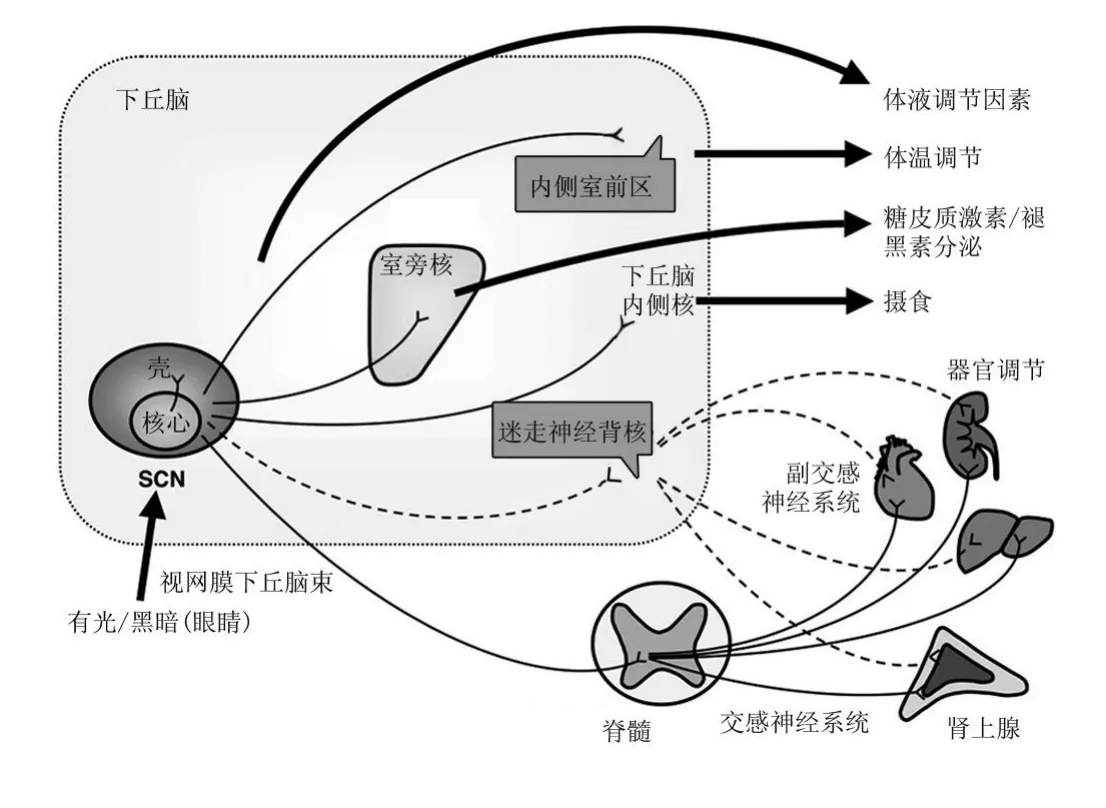
图6 SCN通过神经及体液机制调节中枢及脏器活动[34]
5 小结
应激是所有生物对紧张性事件的适应性反应,对生物的存活具有十分重要的意义。某些刺激能影响生物钟的输出和昼夜节律的表达,这对机体的生理功能和行为都产生影响[40]。杏仁核、海马等重要的脑内核团参与创伤、心理、生理、社会等因素导致的应激反应的过程。
生物钟基因在中枢和外周组织均有表达,对全身各系统的功能产生影响。边缘系统是中枢神经系统的重要组成部分,它对调节情绪和应激反应方面具有重要的作用。PER2蛋白是一个重要的时钟调节器,在维持这些边缘区域功能的完整性方面扮演了重要的角色。本文概述了PER2蛋白参与边缘系统昼夜节律调节对情绪和内脏活动的研究进展,旨在进一步明确昼夜节律在蛋白水平的表达调控机制,同时也为应激研究和因应激产生昼夜节律紊乱的治疗提供研究思路。
[1]李经才,于多,王芳,等.生物钟基因研究新进展[J].遗传,2004,26(1):89-96.
[2]Mazzoccoli G.Clock gene expression levels and relationship with clinical and pathological features in colorectal cancer patients[J].Chronobiol Int,2011,28(10):841-851.
[3]Hoogerwerf WA.Role of clock genes in gastrointestinal motility[J].Am JPhysiol Gastrointest Liver Physiol,2010,299(3):549-555.
[4]Ptitsyn AA,Zvonic S,Gimble JM.Digital signal processing reveals circadian baseline oscillation in majority of mammalian genes[J].PLoSComput Biol,2007,3(6):e120.
[5]KarbovskyĭLL,Minchenko DO,Garmash IaA,et al.[Molecular mechanisms of circadian clock functioning].[Article in Ukrainian][J].Ukr Biokhim Zh,2011,83(3):5-24.
[6]Abe M,Herzog ED,Yamazaki S,et al.Circadian rhythms in isolated brain regions[J].JNeurosci,2002,22(1):350-356.
[7]Abraham U,Prior JL,Granados-Fuentes D,et al.Independent circadian oscillations of Period1 in specific brain areas in vivo and in vitro[J].JNeurosci,2005,25(38):8620-8626.
[8]Harbour VL,Robinson B,Amir S.Variations in daily expression of the circadian clock protein,PER2,in therat limbic forebrain during stable entrainment to a long light cycle[J].JMol Neurosci,2011,45(2):154-161.
[9]Amir S,Stewart J.Motivational modulation of rhythms of the expression of the clock protein per2 in the limbic forebrain[J].Biol Psychiatry,2009,65(10):829-834.
[10]King HA,Hoelz A,Crane BR,et al.Structure of an enclosed dimer formed by the drosophila period protein[J].JMol Biol,2011,413(3):561-572.
[11]徐晨,钱睿哲,金惠铭.生物钟基因与心血管疾病[J].中国病理生理杂志,2006,22(7):1450-1453.
[12]王朝霞.生物钟基因Per与肿瘤[J].国际生殖健康计划生育杂志,2009,28(6):377-379.
[13]王明勇,黄国栋,王晗.斑马鱼生物钟研究进展[J].遗传,2012,34(9):1133-1143.
[14]郑向忠,张亚平,朱定良,等.生物钟基因period的分子生物学[J].动物学研究,1998,19(6):473-481.
[15]王勇,姚勤,陈克平.动物bHLH转录因子家族成员及其功能[J].遗传,2010,32(4):307-330.
[16]王勇,陈克平,姚勤.bHLH转录因子家族研究进展[J].遗传,2008,30(7):821-830.
[17]Sladek M,Rybova M,Jindrakova Z,et al.Insight into the circadian clock within rat colonic epithelial cells[J].Gastroenterology,2007,133(4):1240-1249.
[18]王艳红,文勇立,齐莎日娜,等.哺乳动物生物钟模型及研究进展[J].四川生理科学杂志,2006,28(1):33-35.
[19]Hoogerwerf WA,Hellmich HL,Cornélissen G,et al.Clock gene expression in the murine gastrointestinal tract:endogenous rhythmicity and effects of a feeding regimen[J].Gastroenterology,2007,133(4):1250-1260.
[20]Konturek PC,Brzozowski T,Konturek SJ.Gut clock:implication of circadian rhythms in the gastrointestinal tract[J].J Physiol Pharmacol,2011,62(2):139-150.
[21]Dickmeis T,Foulkes NS.Glucocorticoids and circadian clock control of cell proliferation:at the interface between three dynamic systems[J].Mol Cell Endocrinol,2011,331(1):11-22.
[22]SzántóováK,Zeman M,VeseláA,et al.Effect of phase delay lighting rotation schedule on daily expression of per2,bmal1,rev-erbα,pparα,and pdk4 genes in the heart and liver of Wistar rats[J].Mol Cell Biochem,2011,348(1-2):53-60.
[23]Vanselow K,Kramer A.Role of phosphorylation in the mammalian circadian clock[J].Cold Spring Harb Symp Quant Biol,2007,72:167-176.
[24]浙江省医学会.2006年浙江省医学会心身医学会议论文汇编[C].杭州:2006年浙江省医学会心身医学会议,2006.
[25]Amir S,Lamont EW,Robinson B,et al.A circadian rhythm in the expression of PERIOD2 protein reveals a novel SCN-controlled oscillator in the oval nucleus of the bed nucleus of the striaterminalis[J].JNeurosci,2004,24(4):781-790.
[26]Lamont EW,Robinson B,Stewart J,et al.The central and basolateral nuclei of the amygdala exhibit opposite diurnal rhythms of expression of the clock protein Period2[J].Proc Natl Acad Sci USA,2005,102(11):4180-4184.
[27]Herman JP,Ostrander MM,Mueller NK,et al.Limbic System mechanismsof stressregulation:hypothalamo-pituitary-adrenocortical axis[J].Prog Neuropsychopharmacol Biol Psychiatry,2005:29(8):1201-1213.
[28]PolidarováL,Sládek M,Soták M,et al.Hepatic,duodenal,and colonic circadian clocks differ in their persistence under conditions of constant light and in their entrainment by restricted feeding[J].Chronobiol Int,2011,28(3):204-215.
[29]Thiriet M.Circadian clock[J].Biomathematical and Biomechanical Modeling of the Circulatory and Ventilatory Systems,2012,2:329-356.
[30]Amir S,Stewart J.Behavioral and hormonal regulation of expression of the clock protein,PER2,in the central extended amygdale[J].Prog Neuropsychopharmacol Biol Psychiatry,2009,33(8):1321-1328.
[31]倪银华,吴涛,王露,等.肾上腺糖皮质激素与生物钟基因表达调控的相关研究进展[J].遗传,2008,30(2):135-141.
[32]杨权.下丘脑-垂体-肾上腺皮质轴应激反应的中枢控制[J].生理科学进展,2000,31(3):222-226.
[33]松枝启.肠易激综合征时的消化管运动障碍与中枢神经[J].日本医学介绍,2002,23(9):416.
[34]Nader N,Chrousos GP,Kino T.Interactions of the Circadian CLOCK System and the HPA axis[J].Trends Endocrinol Metab,2010,21(5):277-286.
[35]姚志彬.海马——研究神经科学的理想模型[J].解剖学研究,1989,11(1):17-21.
[36]Miyamoto H,Nakamaru-Ogiso E,Hamada K,et al.Serotonergic integration of circadian clock and ultradian sleep-wake cycles[J].JNeurosci,2012,32(42):14794-14803.
[37]Hongo M.Epidemiology of FGID symptoms in Japanese general population with reference to life style[J].JGastroenterol Hepatol,2011,26(3):19-22.
[38]Hoogerwerf WA,Shahinian VB,Cornélissen G,et al.Rhythms changes in colonic motility are regulated by period genes[J].Am J Physiol Gastrointest Liver Physiol,2010,298(2):143-150.
[39]Heitkemper MM,Cain KC,Deechakawan W,et al.Anticipation of public speaking and sleep and the hypothalamic-pituitary-adrenal axis in women with irritable bowel syndrome[J].Neurogastroenterol Motil,2012,24(7):626-631.
[40]Meerlo P,Sgoifo A,Turek FW.The effects of social defeat and other stressors on the expression of circadian rhythmsrek[J].Stress,2002,5(1):15-22.
猜你喜欢
浙江大学学报(医学版)(2023年2期)2023-08-08 14:12:27
疯狂英语·新读写(2022年6期)2022-11-22 10:35:44
磁共振成像(2022年6期)2022-07-30 08:05:14
基层中医药(2021年5期)2021-07-31 07:58:12
中国医学影像技术(2021年2期)2021-03-30 09:23:58
音乐生活(2020年12期)2020-01-05 05:38:51
奥秘(2018年1期)2018-07-02 10:56:34
皖南医学院学报(2016年4期)2016-03-13 17:30:06
今日文摘(2015年10期)2015-05-30 10:48:04
发明与创新(2015年25期)2015-02-27 10:39:14
