
基于微卫星的中国红火蚁种群遗传结构的研究
2014-11-22 05:41黄奕雯何晓芳陆永跃程代凤
生物安全学报 2014年2期
黄奕雯,何晓芳,陆永跃,曾 玲,程代凤
华南农业大学红火蚁研究中心,广东 广州510642
红火蚁Solenopsis invicta Buren 是一种入侵性强的有害生物(Vinson,1997)。2003年10月以来,我国台湾、广东、香港、澳门、广西、湖南、福建、江西、海南、云南、四川等地区先后报道了红火蚁的发生及危害(陆永跃等,2008、2014;曾玲等,2005a、2005b;Wang et al.,2013)。在新入侵地,该蚁是否因为入侵来源不同或是为了适应新环境变化而发生种群遗传分化等问题值得关注。He et al.(2006)基于COI 基因分析了境内13 个种群的单倍型,认为香港是入侵中国的红火蚁种群中遗传多样性最丰富的地区。邵敬国等(2008a、2008b)利用GP-9 特异性引物和微卫星位点研究了我国红火蚁的社会型和地理种群间遗传差异性。利用微卫星(simple sequence repeat,SSR)的多态性来分析物种进化、生物群体内的遗传变异以及种间关系,甚至在生物个体鉴定等方面已有许多报道:在韩国,研究人员利用SSR 技术成功评估了芒草Miscanthus的遗传 多 样 性(Yook et al.,2014);Wang et al.(2005)利用SSR 技术成功揭示了昆虫致病真菌白僵菌Beauveria bassiana (Bals)Vuill.菌株的遗传多样性和种群结构;在果蝇Drosophila melanogaster Meigen 中也有大量利用SSR 技术进行相关研究的报道(Prokop & Meinertzhagen,2006)。同时,在社会性昆虫协同进化过程中的行为研究和亲缘选择假说中,微卫星标记也已被大量应用(Aanen &Eggleton,2005;Cheng et al.,2013)。
了解红火蚁入侵种群的遗传结构及其多态性,不仅有助于预测这一物种的入侵潜力并推断其生态和进化过程,而且有助于制定相应的控制和管理策略,从而采取有效的措施阻止其进一步扩散,并减少危害。本研究利用SSR 分子标记技术研究中国红火蚁种群的遗传结构,希望能够进一步揭示该蚁种群间存在的遗传变异及其多样性,为控制该蚁提供依据。
1 材料与方法
1.1 试验材料
供试红火蚁样品来自广东、广西、福建、香港等中国4 个红火蚁发生省份(或地区)的14 个种群,以及美国阿拉巴马州的1 个种群。除田间采集的红火蚁样品外,其他样品均为浸泡于75%或95%酒精中、置于-40 ℃低温冰箱保存的标本(表1)。

表1 供试虫体样品信息Table 1 Information of samples used in the study
1.2 试验方法
1.2.1 供试红火蚁的DNA 提取 分别从每个蚁巢中取1 ~10 头工蚁个体,对整头工蚁进行DNA 抽提。其中,田间采集、实验室饲养的红火蚁活体,先放入低温冰箱速冻至晕再进行研磨,酒精浸泡的红火蚁标本需用200 μL 的TE Buffer 浸泡过夜后再提取DNA,以去除其他化学物质对PCR 等后续试验的影响。用天根血液/细胞/组织基因组DNA 提取试剂盒抽提DNA,详细步骤按其使用说明书进行操作。
1.2.2 SSR PCR 扩增 参考Krieger & Keller(1997)的SSR 引物序列,合成8 对引物(表2),并参照其反应体系和程序进行红火蚁工蚁的SSRPCR 扩增。
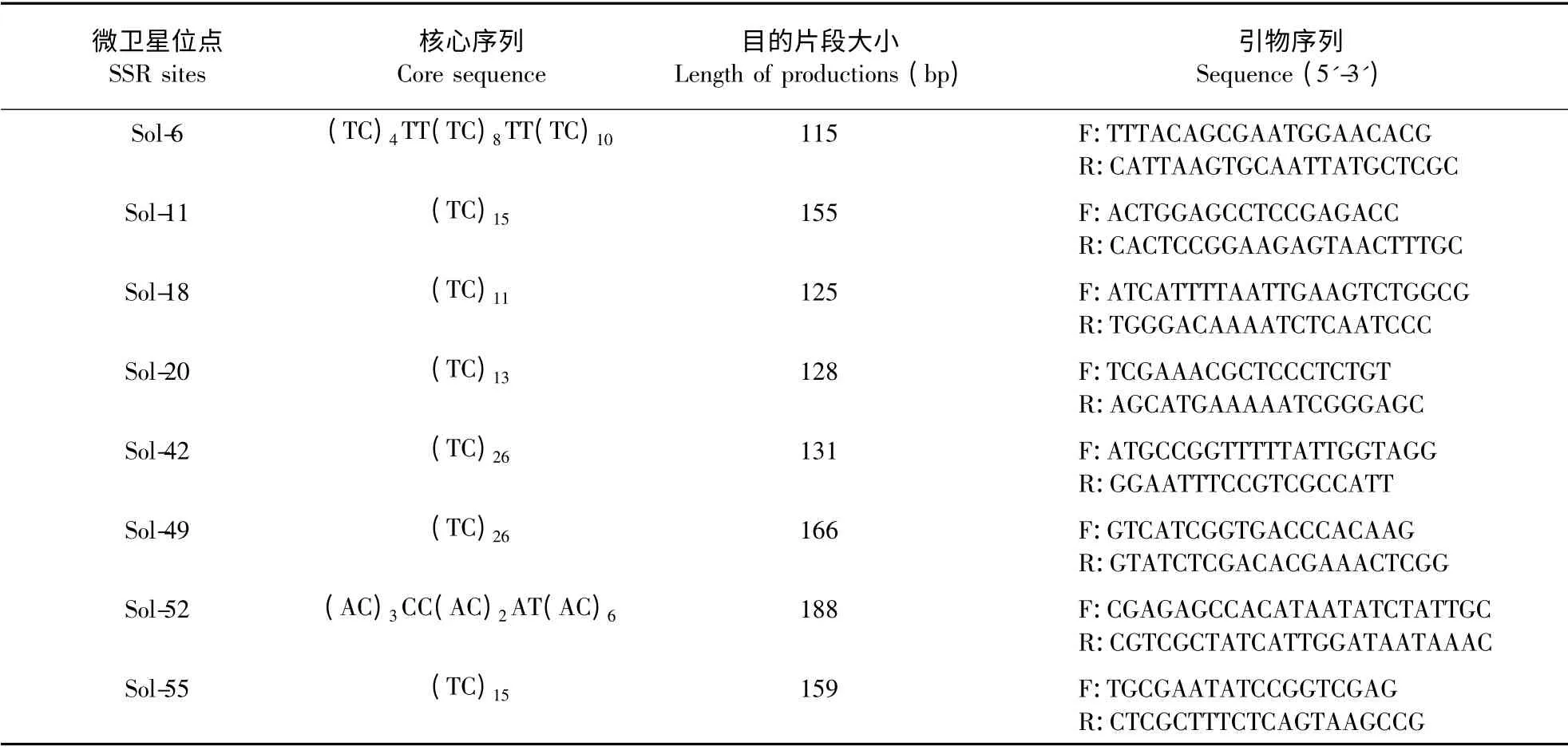
表2 用于工蚁SSR-PCR 扩增的引物序列Table 2 Sequences of primers used in the SSR-PCR
1.2.3 SSR-PCR 产物的PAGE 检测 在25 μL 体系的SSR-PCR 产物中加入5 μL 6 ×Loading Buffer混匀,吸取1.5 μL 混合液进行加样电泳,控制电压为80 V 恒压,电泳100 min。采用快速银染法进行染色,将显影后的PAGE 胶置于凝胶成像系统中成像观察并拍照,将所得图像直接在成像系统中进行处理,对SSR-PCR 扩增结果读带,根据50 bp DNA Ladder 标准分子质量标记标定目的片段分子质量的大小,记录清晰、稳定出现且产物长度在100 ~200 bp 的扩增带。
1.3 数据处理与分析
按照电泳图谱中同一位置上的DNA 条带进行统计,以二倍体形式记录扩增带,从大到小依次记作A、B、C……(李伟丰等,2007)。应用Popgene 1.32软件对全部种群和各单个种群分别进行遗传参数分析,在假定各种群处于Hardy-Weinberg 平衡状态下,计算多态带数(Np)、多态位点百分率(P)、观测等位基因数(NA)、有效等位基因数(NE)、基因多样性指数(Nei's)、Shannon 信息指数(I)、表观杂合度(HO)和预期杂合度(HE)、种群内近交系数(FIS)、总近交系数(FIT)、种群间分化系数(FST)、种群间基因流(Nem)、Nei's 标准遗传距离(D)和遗传相似度(I)。根据D,利用NTSYSpc2.1 软件对各种群进行非加权组平均法(UPGMA)聚类分析。
2 结果与分析
2.1 SSR-PCR 扩增结果
在8 对微卫星扩增位点中,每个位点在这15个红火蚁种群中都出现清晰的扩增条带,没有出现零等位基因的情况。但在Sol-52 位点上只出现1个等位基因,且没有表现出多态性。
2.2 红火蚁地理种群的遗传多样性
对7 对表现出多态性的扩增结果分析发现,7对微卫星引物在15 个红火蚁种群中共检测到28个等位基因,每个位点获得的等位基因数有2 ~6个,其中Sol-20 位点上拥有最多的等位基因为6个,Sol-6 位点和Sol-18 位点上的等位基因最少,仅为2 个(表3)。这表明用于SSR-PCR 扩增的这7对引物具有一定的特异性。各微卫星位点的表观杂合度HO在0.1957 ~0.3582 之间,平均值为0.2802;预期杂合度HE在0.1705 ~0.3199 之间,平均值为0.2623。各位点的等位基因表观杂合度与预期杂合度基本一致,均相差不大,均符合Hardy-Weinberg 平衡。
15 个红火蚁地理种群的遗传变异参数见表4。各种群的观测等位基因数NA在2.1057 ~3.4215 之间,平均为2.7028;有效等位基因数NE在1.5095 ~2.8253 之间,平均为2.1066。Shannon 信息指数I在0.2522(广西南宁)~0.3524(广东深圳)之间,平均为0.3174;表观杂合度HO在0.2433(广西南宁)~0.3284(广东深圳)之间,平均为0.2848;预期杂合度HE在0.2324(广西南宁)~0.3096(广东深圳)之间,平均为0.2708;基因多样性指数Nei's 在0.1977(广西南宁)~0.2979(广东深圳)之间,平均为0.2629;各个种群内的多态位点百分率P 为37.17% ~49.73%,平均每个种群为43.63%,最高的是广东深圳种群,最低的是广西南宁种群。从表中可以看出,种群的Shannon 信息指数I、表观杂合度HO、预期杂合度HE、基因多样性指数Nei's 以及多态位点百分率P 呈正相关。
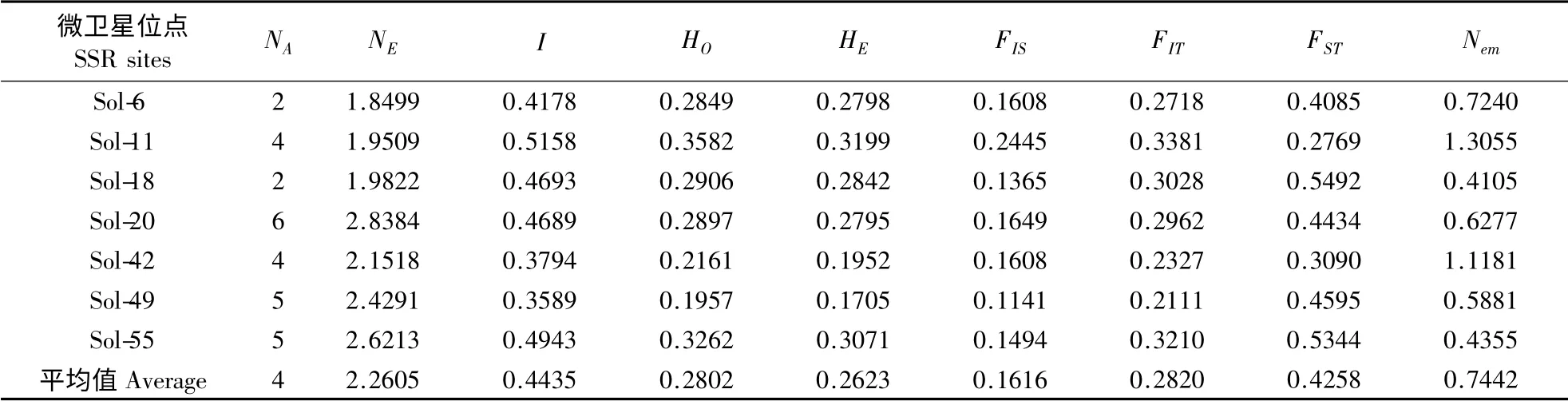
表3 7 个微卫星位点的遗传变异参数、F-statistics 和基因流Table 3 Indices of genetic diversity and genes flow deduced by 7 SSRs primers

表4 15 个红火蚁地理种群在7 个微卫星位点上的遗传变异参数Table 4 Indices of genetic diversity of 15 S.invicta populations based the 7 SSRs primers
2.3 红火蚁地理种群的遗传分化
15 个红火蚁地理种群在7 个微卫星位点上的F-statistics 分析结果见表3。从表中可以看出,15个红火蚁地理种群在各微卫星位点上的FIS为0.1141(Sol-49)~0.2445(Sol-11),平均值0.1616;FIT为0.2111(Sol-49)~0.3381(Sol-11),平均值0.2820。种群间的分化系数FST在Sol-18 位点最大为0.5492,在Sol-11 位点最小为0.2769,FST平均值为0.4258。从FST可知,供试红火蚁种群有42.58%的遗传变异发生在种群间,57.42%的遗传变异发生在种群内。种群间的基因流Nem均较小,范围从0.4105(Sol-18)~1.3055(Sol-11),平均值仅为0.7442。
供试红火蚁地理种群间Nei's 标准遗传距离D和遗传相似度I 见表5,从表中可以看出,所研究的15 个红火蚁地理种群两两之间的遗传相似度范围在0.7625 ~1.0000 之间,均大于0.6,表明各种群相互之间存在着非常频繁的基因交流。Nei's 标准遗传距离D 显示出,河源种群和中山种群之间的遗传距离最大为0.2711,集美种群和厦门种群之间的遗传距离最小为0,几乎没有遗传差异。其中,河源红火蚁地理种群与其他地理种群间的遗传距离范围在0.1248 ~0.2711 之间,均相对高于其他各种群间的遗传距离;而美国阿拉巴马州红火蚁种群与国内红火蚁各种群之间的遗传距离在0.0244 ~0.1651 之间,表明其与国内红火蚁种群间的遗传差异不大。

表5 15 个红火蚁地理种群间Nei's 标准遗传距离(D)和遗传相似度(I)Table 5 Nei's genetic distance and genetic identity coefficient of the 15 population for S.invicta
2.4 聚类分析
根据Nei's 标准遗传距离,利用NTSYSpc2.1 中的UPGMA 法对15 个红火蚁地理种群进行聚类分析,结果见图1。从图中可以看出,系统发育树首先分为2 个大分支,其中河源种群与其他种群之间的遗传距离最远,存在明显的遗传分化,成为独立的一支。然后其他种群又由2 个分支组成,中山种群与其他剩余种群之间的遗传距离相对较远,成为独立的一支;而其他剩余种群又明显地分为2 个大分支,香港种群和珠海种群聚为一支,深圳、梅州、博罗、增城、广州、厦门、集美、泉州、东莞、南宁与美国阿拉巴马州种群又构成一个十分复杂的分支。
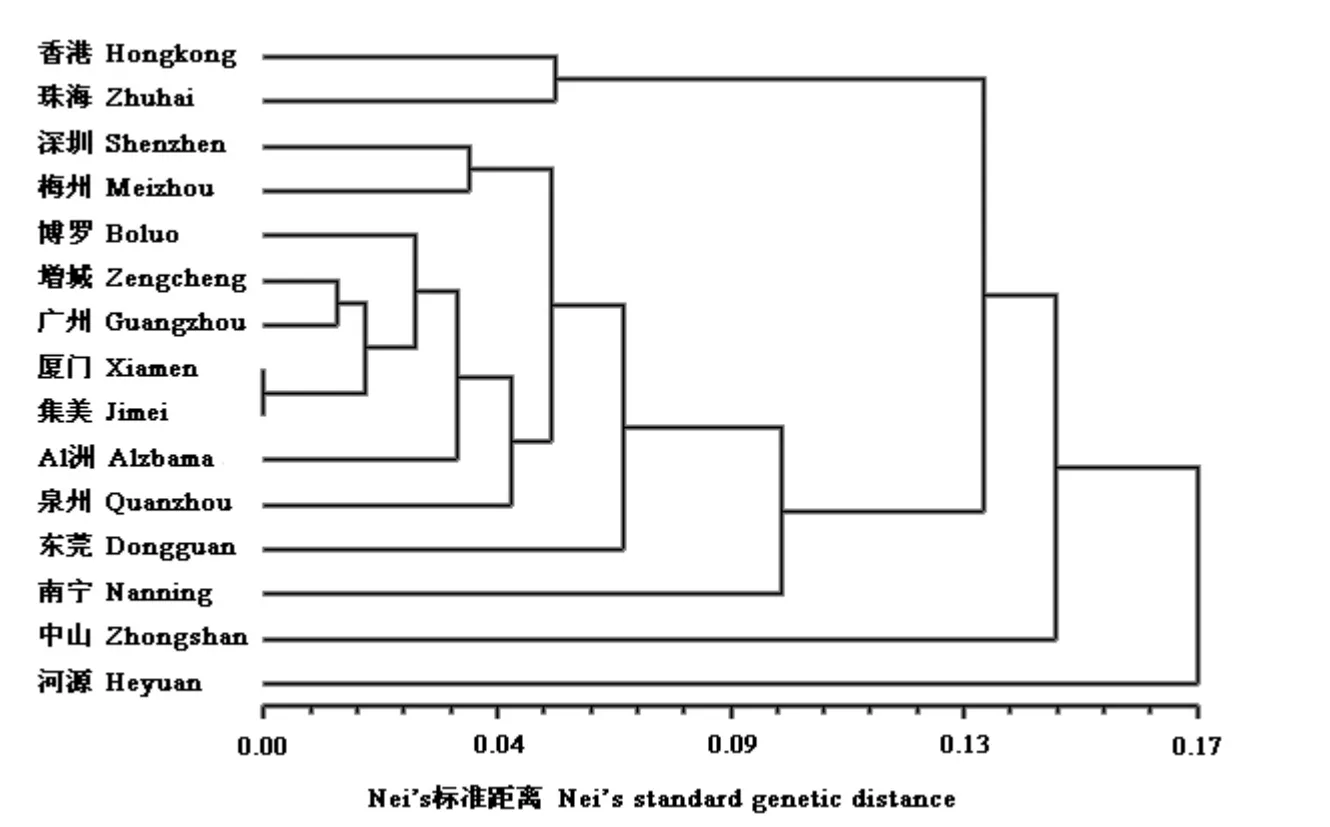
图1 红火蚁地理种群间Nei's 标准遗传距离的UPGMA 法聚类图Fig.1 UPGMA cluster analysis using the Nei's standard genetic distance indices for S.invicta populations
3 讨论
3.1 Hardy-Weinberg 平衡
Hardy-Weinberg 平衡定律也叫遗传平衡定律,是指在理想状态下,各等位基因的频率和等位基因的基因型频率在遗传中稳定不变,即保持着基因平衡。所谓的理想状态需满足5 个条件:种群足够大,种群中个体间可以随机交配,不发生突变,没有新基因加入,没有自然选择。显然,遗传平衡在自然状态下是无法达到的,但在一个足够大的种群中,如果个体间自由交配,且没有明显的自然选择,往往可以近似地看作符合遗传平衡(Stark,2006)。本研究中的15 个红火蚁地理种群在各微卫星位点上的等位基因表观杂合度与预期杂合度基本相符,均符合Hardy-Weinberg 平衡,即表明各等位基因的基因型频率近似稳定。
3.2 红火蚁地理种群的遗传多样性分析
一般所说的遗传多样性是指种内的遗传多样性,即种内个体之间或一个群体内不同个体之间的遗传变异总和,它反映了一个物种适应环境的能力及其被改造和利用的潜力。
在自然进化过程中,如果种群发生一定程度的隔离,那么,地理种群间的遗传差异就会逐步增加,而地理种群内的遗传差异则会逐渐减小,从而使得地理种群内以及地理种群间杂合体的比例发生变化。因此,种群杂合度的高低反映了群体的遗传一致性程度,种群杂合度越低,表明该种群的遗传一致性越高,从而导致种群遗传变异程度越低,种群遗传多样性越低(李伟丰等,2007)。本研究中的15个红火蚁地理种群的表观杂合度HO在0.2433(广西南宁)~0.3284(广东深圳)之间,平均为0.2848;预期杂合度HE在0.2324(广西南宁)~0.3096(广东深圳)之间,平均为0.2708。可以看出,广东深圳红火蚁种群的表观杂合度和预期杂合度相对于其他种群均较高,表明这个地区的红火蚁种群的遗传变异程度较高,遗传多样性也相对更丰富一些。
Nei & Li(1979)认为,基因多样性指数是群体内基因变异的理想测度,它可以适用于任何生物。15 个红火蚁种群的基因多样性指数Nei's 在0.1977(广西南宁)~0.2979(广东深圳)之间,平均为0.2629。广东深圳种群的基因多样性指数在所有种群中最高,这与该种群拥有最高的表观杂合度和预期杂合度相一致,表明广东深圳的红火蚁种群内的遗传分化程度和遗传多样性比较高。Botstein et al.(1980)首先提出了衡量基因变异程度的多态位点百分率P,当P >50%时为高度多态位点,25% <P≤50%时为中度多态位点,当P≤25%时为低度多态位点。15 个红火蚁地理种群在7 个微卫星位点上的多态位点百分率P 为37.17% ~49.73%,平均为43.63%,表明这7 个微卫星位点均具有中等程度的多态性,可以作为有效的遗传标记用于红火蚁地理种群遗传多样性和系统发生关系的分析,同时表明红火蚁种群具有比较丰富的遗传多样性。
3.3 红火蚁地理种群的遗传分化与变异
产生遗传分化的原因有多方面,包括有性生殖、体细胞突变、选择、基因流、遗传漂变以及环境的影响等(李伟丰等,2007)。本试验所研究的15个红火蚁种群分别来自我国广东、广西、福建、香港4 个省(地区)和美国阿拉巴马州,在不同生境作用下,红火蚁种群会形成一定程度的遗传分化,种群间分化系数FST值越大,说明种群的分化程度越高。15 个红火蚁地理种群间分化系数的平均值达到0.4258,表明各种群之间有较高程度的分化,接近一半的遗传变异发生在种群间。种群内近交系数FIS与总近交系数FIT的值均较小,平均值分别仅为0.1616与0.2820,表明这15 个种群在各种群内部的近交程度都比较低。
基因流是指生物个体从其发生地分散出去而导致不同种群间基因交流的过程,可发生在同种或不同种的生物种群之间。基因流的强弱程度因不同的种群、不同的时间地点而有很大差异,但其基本作用是削弱种群间的遗传差异,提高遗传多样性的程度。由此可见,基因流是影响种群遗传分化的另一个重要因素,高水平的基因流可以防止种群间的遗传分化,使种群趋于一致。而遗传漂变是指由于某种随机因素,某一等位基因的频率在群体中出现世代传递的波动现象。当种群间的基因流Nem>1时即能有效抑制由遗传漂变引起的遗传分化,当Nem<1 时基因流就不足以抵制种群内因遗传漂变而引起的种群分化(Slatkin,1985)。15 个红火蚁地理种群间的Nem仅为0.7442,说明各种群间存在着较高程度的遗传分化,而除了地理隔离与环境因素,遗传漂变很可能是引起较大分化的一个重要原因。
利用遗传距离可以估测种群间的进化关系,关于计算遗传距离的方法很多,但目前应用较广泛的遗传距离指标是Nei & Li(1979)于20 世纪70年代提出来的,即为Nei's 标准遗传距离。根据遗传距离D 显示,河源红火蚁种群与其他种群间的遗传距离均相对高于其他各种群间的遗传距离,表明河源种群与其他地理种群之间的遗传差异比较大,存在明显的遗传分化,可能是较为原始的类型;而美国阿拉巴马州红火蚁种群与国内红火蚁各种群之间的遗传距离均较小,表明其与国内红火蚁种群间的遗传差异不大。从根据Nei's 标准遗传距离构建的UPGMA 聚类图可以看出,厦门和集美的红火蚁种群聚为一支,广州五山和增城的红火蚁种群聚为一支,这说明遗传距离与实际地理距离之间有一定的关系;但是美国阿拉巴马州的红火蚁种群与国内红火蚁种群之间的遗传距离比起河源种群与国内其他种群之间的遗传距离却小得多,这说明遗传距离与实际地理距离之间不一定呈正相关。从供试红火蚁种群复杂的遗传结构可以看出,中国红火蚁各种群间既存在着由于地理因素造成的遗传分化,又存在着因为种苗、花卉、草皮等带土植物的贸易调运所带来的种群扩散。
3.4 中国红火蚁种群的扩散途径
一般来说,在正常的种群扩散过程中,如果没有大的自然屏障,地理上相邻地区种群间的生物多样性应该比较接近。从UPGMA 聚类图中可以看到,厦门和集美、广州和增城这些地理距离相近地区的红火蚁种群都首先聚成了一支,说明它们之间的遗传距离较近,可能是红火蚁种群通过自然扩散而形成。
从UPGMA 聚类图上还可以看到,广州五山和增城的红火蚁种群与厦门和集美种群聚成一支,其间的遗传距离小于与广东其他地区红火蚁种群之间的遗传距离,这些地区的地理距离相隔较远,且多后型蚁巢的扩散方式主要是通过分巢进行,独立建巢能力和远距离迁移能力都比较弱(陆永跃等,2008;许益镌等,2006),因此认为红火蚁不可能靠其自身的迁飞能力扩散,只能靠人为传播,如因园艺植物污染、草皮污染、土壤废土移动、园艺农耕机具设备污染、空货柜污染、车辆等运输工具污染等而进行长距离传播。由于这些地区的生境都适宜红火蚁的定殖,当被红火蚁污染的带土植物经过频繁调运而又未做有效处理时,则利于红火蚁的远距离扩散,从而造成地理距离相隔较远的地区之间存在较小的遗传距离。因此,在进行种苗、花卉、草皮等植物性产品的贸易调运时,为避免红火蚁的传播扩散,一般要求不带土。
李伟丰,杨朗,唐侃,曾玲,梁广文.2007.中国桔小实蝇种群的微卫星多态性分析.昆虫学报,50(12):1255-1262.
陆永跃.2014.中国大陆红火蚁远距离传播速度探讨和趋势预测.广东农业科学,41(10):70-72.
陆永跃,梁广文,曾玲.2008.华南地区红火蚁局域和长距离扩散规律研究.中国农业科学,41(4):1053-1063.
邵敬国.2008a.我国红火蚁Solenopsis invicta 社会型鉴定及不同地理种群遗传差异性分析.武汉:华中农业大学.
邵敬国,罗礼智,陈浩涛,江幸福,雷朝亮.2008b.应用多元PCR 技术对我国红火蚁社会型的鉴定.昆虫学报,51(1):551-555.
许益镌,陆永跃,曾玲.2006.红火蚁局域扩散规律研究.华南农业大学学报,27(1):40-42.
曾玲,陆永跃,陈忠南.2005a.红火蚁监测与防治.广州:广东科技出版社.
曾玲,陆永跃,何晓芳,张维球,梁广文.2005b.入侵中国大陆的红火蚁的鉴定及发生为害调查.昆虫知识,42(2):144-148.
Aanen D K and Eggleton P.2005.Fungus-growing termites originated in African rain forest.Current Biology,15:851-855.
Botstein D,White R L,Skolnick M and Davis R W.1980.Construction of a genetic linkage map in man using restriction fragment length polymorphisms.American Journal of Human Genetics,32:314-331.
Cheng S,Lee C T,Wan M N and Tan S G.2013.Microsatellite markers uncover cryptic species of Odontotermes (Termitoidae:Termitidae)from Peninsular Malaysia.Gene,518:412-418.
He X F,Lu Y Y,Zhang W Q and Zeng L.2006.Three haplotypes found in populations of the red imported fire ant invading China.Acta Entonologica Sinica,49:1046-1049.
Krieger M J B and Keller L.1997.Polymorphism at dinucleotide microsatellite loci in fire ant Solenopsis invicta populations.Molecular Ecology,6:997-999.
Nei M and Li W H.1979.Mathematical model for studing genetic variation in terms of restriction endonnuclese.Proceedings of National Academy of Sciences,76:5269-5280.
Prokop A and Meinertzhagen I A.2006.Development and structure of synaptic contacts in Drosophila.Seminars in Cell& Developmental Biology,17:20-30.
Slatkin M.1985.Gene flow in natural populations.Annual Review of Ecology and Systematics,16:393-430.
Stark A E.2006.A clarification of the Hardy-Weinberg law.Genetics,174:1695-1697.
Vinson S B.1997.Invasion of the red imported fire ant (Hymenoptera:Formicidae):spread,biology and impact.American Entomologist,43:23-39.
Wang L,Lu Y Y,Xu Y J and Zeng L.2013.The current status of research on Solenopsis invicta Buren (Hymenoptera:Formicidae)in mainland China.Asian Myrmecology,5:125-138.
Wang S,Miao X,Zhao W,Huang B,Fan M,Li Z and Huang Y.2005.Genetic diversity and population structure among strains of the entomopathogenic fungus,Beauveria bassiana,as revealed by inter-simple sequence repeats (ISSR).Mycological Research,109:1364-1372.
Yook M J,Lim S H,Song J S,Kim J W,Zhang C J,Lee E J,Ibaragi Y,Lee G J,Nah G and Kim D S.2014.Assessment of genetic diversity of Korean Miscanthus using morphological traits and SSR markers.Biomass and Bioenergy,66:81-92.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
特产研究(2022年6期)2023-01-17
今日农业(2021年8期)2021-07-28
学苑创造·A版(2021年4期)2021-04-18
销售与市场(营销版)(2021年4期)2021-04-16
种子(2021年3期)2021-04-12
四川动物(2017年6期)2017-12-12
四川动物(2017年4期)2017-07-31
外语教学理论与实践(2016年1期)2016-06-11
中学生理科应试(2016年7期)2016-05-14
