
桑天牛幼虫龄数林间识别研究
2014-11-17 12:04LEBaothanh颜学武
湖南林业科技 2014年1期
LE Bao-thanh,颜学武, 许 潜
(1.越南林业大学, 越南 河内 100803; 2.湖南省林业科学院, 湖南 长沙 410004;3.湘潭市林业局, 湖南 湘潭 411100)
桑天牛幼虫龄数林间识别研究
LE Bao-thanh1,颜学武2*, 许 潜3
(1.越南林业大学, 越南 河内 100803; 2.湖南省林业科学院, 湖南 长沙 410004;3.湘潭市林业局, 湖南 湘潭 411100)
通过室内饲养繁殖和野外采集,获得含有各龄桑天牛幼虫的杨树枝干,室内条件下分龄收集幼虫虫粪。对各龄幼虫虫粪进行区分,分离出粪粒和啃食残渣,对最末排粪孔长径和短径以及粪粒的理化性质与龄数关系进行研究。结果表明:桑天牛幼虫粪粒平均宽度值为1.2115mm,平均长度值为2.1018mm;粪粒中木屑平均宽度值为0.1130mm,平均长度值为0.2073mm;粪粒中蛋白质的平均含量为0.1426%,总糖的平均含量为0.6125%,纤维素平均含量为46.72%,木质素平均含量为16.12%。最末排粪孔长径和短径、粪粒长度和宽度、粪粒中木屑长度和宽度、纤维素含量、蛋白质含量均与幼虫龄数存在线性相关性,可作为快速识别幼虫龄数的指标,其中粪粒宽度比较稳定,在各龄数间无重叠,为林间快速识别虫龄的最佳指标。
桑天牛; 幼虫虫粪; 幼虫龄数; 理化性质
天牛幼虫龄数的划分,目前基本是根据Dyar规律对采集的不同发育阶段天牛幼虫虫体坚硬部分进行测定后,应用频次分布、均差分析确定峰值数量划分龄数,进而通过回归分析筛选出最佳分龄构造(所测量的器官部位),以应用于实际分龄工作。上述分龄方法获得的结果总体较为准确,目前黄斑星天牛(Anoplophoranobilis)、松墨天牛(Monochamusalternatus)、星天牛(Anoplophorachinensis)、眉斑并脊天牛(Gleneacantor)、桑天牛(Aprionagermari)等幼虫的龄数已经陆续得到确定[1-5]。不足之处在于研究和实际应用需要剖木取虫,颇为不便。利用虫粪特征推断幼虫龄数和虫口密度在食叶害虫的研究中早有报道,如利用虫粪大小判断模毒蛾(Lymantriamonacha)、欧洲云杉锯角叶蜂(Gfilpiniahercyniae)、云杉卷叶蛾(Choristoneurafumiferana)、马尾松毛虫(Dendrolimuspunctatus)的龄数[6-9]。许多天牛在贮食危害的同时,有明显的排粪孔将粪便和碎屑排出蛀道,随着龄数的增加,排粪孔直径、粪粒大小、粪粒中纤维素含量、蛋白质含量都呈现逐渐增加的趋势[10-13]。桑天牛的幼虫是其主要危害虫态,生活时间长,危害严重。由于隐蔽生活,人们对幼虫在蛀道内的活动习性所知甚少,即使象桑天牛这样重要的易成灾种类,幼虫龄数划分和分龄形态特征等也未得到解决,对幼虫活动特点的了解仅局限于通过排粪获得的间接推断。此外,不同龄期和发育阶段的幼虫,其蛀道结构、活动习性等均存在差异,所对应的防治技术也存在差别。我们通对虫龄林间快速判别进行研究,可为创新天牛防治措施、提高防治效果提供参考。
1 材料与方法
1.1试验材料
1.1.1 初龄幼虫粪粒的采集
2011年7—8月,在室内饲养桑天牛成虫,将2年生杨树枝条插入装满湿沙的水桶中,供其产卵,得到21条产过卵的杨树枝条,以3个为1组,共分7组。通过观察发现桑天牛初产卵一般在7~10天后孵化,初龄幼虫会在刻槽一侧咬1个排粪孔,故7天后每天固定时间逐组小心解剖杨树枝条,将1龄幼虫虫粪(粪粒和啃食残渣的混合物)用毛笔轻轻刷入指形管,直至观察到7组杨树枝条内的1龄幼虫蜕皮为止。将收集到的虫粪在8倍显微镜下辨别、分离粪粒和啃食残渣,将粪粒挑出装入指形管,编号。
1.1.2 其它龄幼虫粪粒的采集
2011年4—11月在镇江市上党镇杨树试验林、南京钟山风景区东侧杨树林分别采集有虫枝干,将枝干带回实验室沿蛀道竖剖,小心取出幼虫,测量头壳宽度,并根据以下标准判断幼虫虫龄:1.819±0.093mm为2龄、2.550±0.123mm为3龄、3.686±0.140mm为4龄、4.822±0.123mm为5龄、5.797±0.105mm为6龄[5]。按头壳宽判定虫龄后,放回原枝干内,并用铁丝拧紧固定枝干。枝干两头用黑布包裹并保湿,对各枝干按虫龄大小进行标记。每4h打开1次,取出虫粪,分离粪粒和啃食残渣,收集2~6龄的粪粒,装入指形管内并按龄数对应编号,每龄幼虫虫粪不少于3个样本。
1.1.3 各龄幼虫的蛀道采样
在粪粒采集地进行虫害木采样,并带回实验室,解剖蛀道,取出幼虫,测量头壳宽度判断龄数,最终得到包含2龄幼虫的蛀道23株、3龄幼虫的15株、4龄幼虫的15株、5龄幼虫的10株、6龄幼虫的9株,加上在室内养虫获得1龄幼虫蛀道10株,共82株[5]。
1.2试验方法
1.2.1 粪粒形态和结构分析
分别随机选取各龄幼虫粪粒(1龄幼虫粪粒在显微镜下筛选,其余幼虫粪粒用肉眼直接辨别)30颗,用双目连续变倍体式显微镜(JSZ6S)将粪粒拍成8倍图像,并通过DT2000图像分析软件对各龄粪粒的长度和宽度进行测量;同时观察并描述粪粒的形状、颜色、质地等特征。重复3次。
桑天牛幼虫的排泄物含有未消化成分,即粪粒中含有木屑,不同龄幼虫对食物的消化程度存在差异,利用木屑粗细程度可间接判断虫龄。每个虫龄取10颗粪粒放入培养皿中碾碎,过筛(标准筛孔径0.154mm),加水稀释漂洗,取筛上部分物质(未消化的木屑)在60℃下烘干至恒重。置于10倍显微镜下拍照,并通过图像分析软件测量木屑的长度和宽度。重复3次。
1.2.2 总糖含量的测定
采用蒽酮比色法[14-15],其详细步骤如下。
① 标准曲线的制作。取7支干燥洁净的试管,编号后按表1加入试剂。
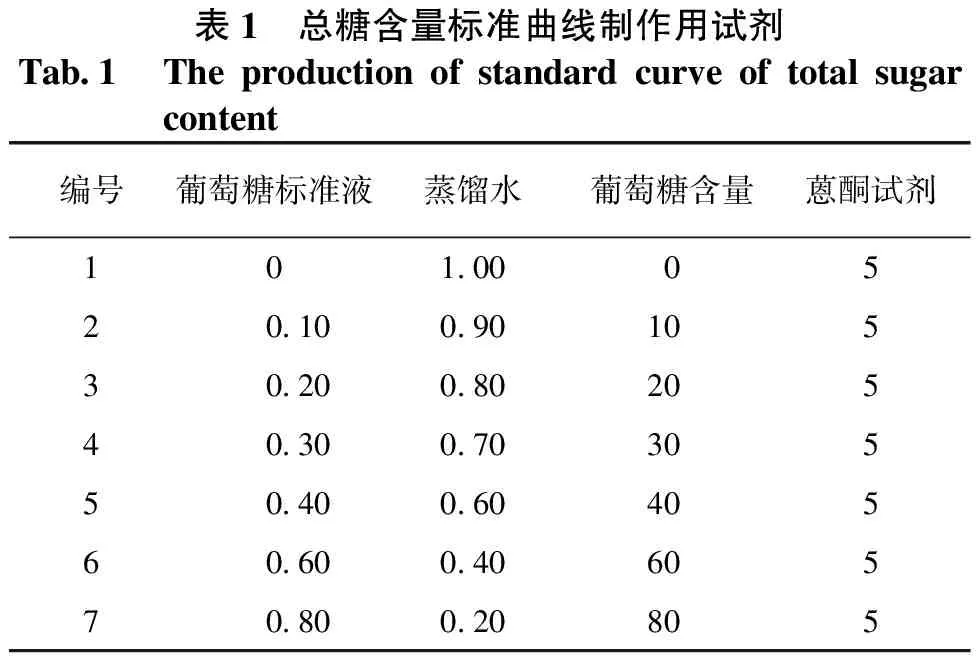
表1 总糖含量标准曲线制作用试剂Tab1 Theproductionofstandardcurveoftotalsugarcontent编号葡萄糖标准液蒸馏水葡萄糖含量蒽酮试剂1010005201009010530200802054030070305504006040560600406057080020805
每管加入葡萄糖标准液和蒸馏水后,同时各管都加入5mL蒽酮试剂混匀,迅速置于沸水浴中,准确加热10min,取出后流动水冷却至室温,平衡20min。用分光光度计在620nm波长下进行比色,记录吸光值。重复3次,以吸光值(A620 nm)为横坐标,葡萄糖含量为纵坐标绘制标准曲线。
② 样品中总糖含量的测定。取粪粒样品在80℃下烘至恒重,然后称取0.2g烘干样品,置于100mL试管中,加入20mL去离子水,塑料薄膜封口,于沸水中提取30min(提取3次),提取液过滤至100mL容量瓶中,用去离子水反复漂洗试管及残渣,定容至100mL。吸取样品提取液1mL,加去离子水1mL,充分振荡后立即将试管放入沸水浴,逐管保温1min,取出后冷却至室温。以空白作对照,在620nm波长下测定样品的吸光度值。重复3次,取平均值。
1.2.3 可溶性蛋白含量的测定
采用考马斯亮蓝G — 250染色法[14],其详细步骤如下。
① 标准曲线的制作。取7支具塞试管,按表2加入试剂(0~100μg/mL的标准蛋白)。混合均匀后,向各管中加入5mL考马斯亮蓝G — 250溶液,摇匀(不要太剧烈,以免产生大量气泡难于消除),放置5min左右,用1cm光径比色皿在595nm下比色测定吸光度。以蛋白质含量(μg)为横坐标,以吸光度为纵坐标绘制标准曲线。
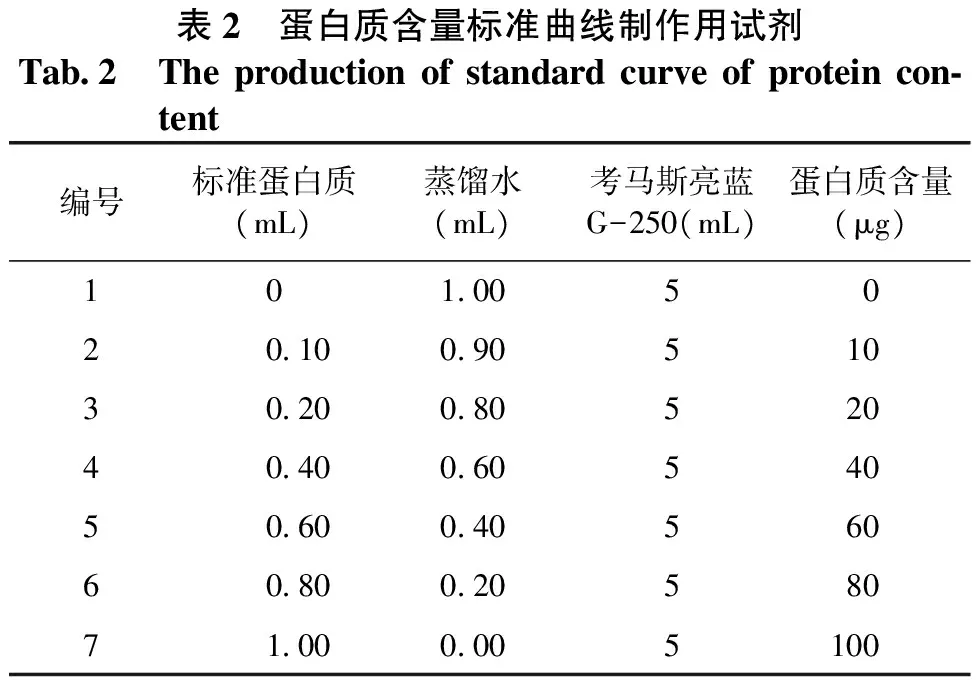
表2 蛋白质含量标准曲线制作用试剂Tab2 Theproductionofstandardcurveofproteincon⁃tent编号标准蛋白质(mL)蒸馏水(mL)考马斯亮蓝G-250(mL)蛋白质含量(μg)10100502010090510302008052040400605405060040560608002058071000005100
② 样品中蛋白质含量的测定。精确称取0.2g干粪粒,用5mL去离子水研磨成匀浆,放入超声波清洗器,经过60min超声提取(温度50℃),再经20000r/min离心20min,取上清液1mL(视蛋白质含量适当稀释)于试管中。用定量加样器于每支试管加入5mL考马斯亮蓝G — 250溶液,充分混合,放置2min(以空白作对照)后,在595nm波长下测定样品的吸光度值。重复3次,取平均值。
1.2.4 纤维素、木质素含量测定
称取各龄干虫粪粒0.2g,放入100mL烧杯中,加入70mL的2M盐酸溶液,置于100℃沸水浴中50min,之后用3号砂芯漏斗过滤,依次将残渣用无水乙醇和丙酮洗涤2次至中性,然后在80℃烘箱中烘至恒重,即为纤维素、木质素、硅酸盐的混合物(残渣1);将残渣1置于150mL烧杯中,加入10mL 72%硫酸,20℃降解4h后加入90mL蒸馏水,室温过夜,次日过滤后残渣用蒸馏水洗至中性烘干至恒重,即为木质素、硅酸盐混合物(残渣2)。残渣2在550℃马福炉中灰化5h,干燥器中平衡至恒重,即为硅酸盐(灰分3)(陈洪章, 2005)。

1.2.5 最末排粪孔大小分析
野外观察发现,桑天牛幼虫期会形成多个排粪孔,排粪孔为椭圆形,伴随杨树的生长,大部分树干上天牛幼虫的排粪孔会变小或者被新生的树皮覆盖。但在一段时期内仅使用一个排粪孔排粪,即最末的排粪孔。虫害木带回实验室后,测量各龄幼虫蛀道的最末排粪孔长短径。
1.3数据分析
用SPSS16.0软件进行统计分析;单因素方差分析、多重比较采用LSD、Duncan法;同时采用Pearson相关分析,并进行回归分析[16]。
2 结果与分析
2.1不同龄数幼虫粪粒外形特征
对各龄幼虫的虫粪颜色、形状、质地观察结果见表3。

表3 不同龄数幼虫粪粒外形特征Tab3 Theexternalmorphologicalcharacteristicsoffecalpelletbydifferentinstarlarvae龄数形状颜色质地1粪粒大小不一,呈圆柱形或椭圆形;两端颜色均匀,多为乳白色,少数浅黄色较紧密,表面平整,纹理不清晰较平滑2粪粒大小较均一,多呈圆柱形,两端平滑颜色均匀,多为浅乳白色,少数为红棕色较紧密,表面较平整,有多条横纹或斜纹3粪粒较2龄明显增大,呈圆柱形,一端略颜色均匀,多为乳白色,少数为浅棕色或较紧密,表面不平整,有明显纹理凹,一端略凸红棕色4粪粒多椭圆形或圆柱形,呈圆柱形的中部颜色略有不均,椭圆形的多为浅红棕色,较紧密,表面较粗糙,有明显的横纹或斜纹略弯曲圆柱形的多为乳白色5粪粒较4龄明显增大,大小较均一,粪粒颜色均匀,多为棕色,少数呈浅棕色紧密,表面较平整,表面横纹较明显多呈圆柱形,两端平截6粪粒多呈圆柱形,两端较平截颜色均匀,多数为红棕色,少数呈浅红棕色紧密,表面粗糙,有明显纹理
2.2不同龄数幼虫粪粒结构差异
由表4和图1可知: 各龄幼虫粪粒长、宽度及粪粒中木屑长、宽度均存在显著差异(F=3.145,P=0.000<0.05;F=380.084,P=0.000<0.05;F=43.070,P=0.000<0.05;F=71.767,P=0.000<0.05),虫粪宽度、长度在龄数间的差异均达显著水平,虫粪中木屑宽度在低龄幼虫间差异不显著,5、6龄间及其与1~4龄间差异显著,1~2龄与3~5龄及6龄间的木屑长度差异显著。其中6龄幼虫粪粒的宽度、长度平均值及粪粒中木屑宽度、长度平均值最大,与其它龄数幼虫均达到显著差异;1龄幼虫的粪粒宽度、长度平均值及粪粒中木屑宽度平均值最小。幼虫期粪粒平均宽度值为1.2115mm,平均长度值为2.1018mm;粪粒中木屑宽度平均值为0.1130mm,长度平均值为0.2073mm。
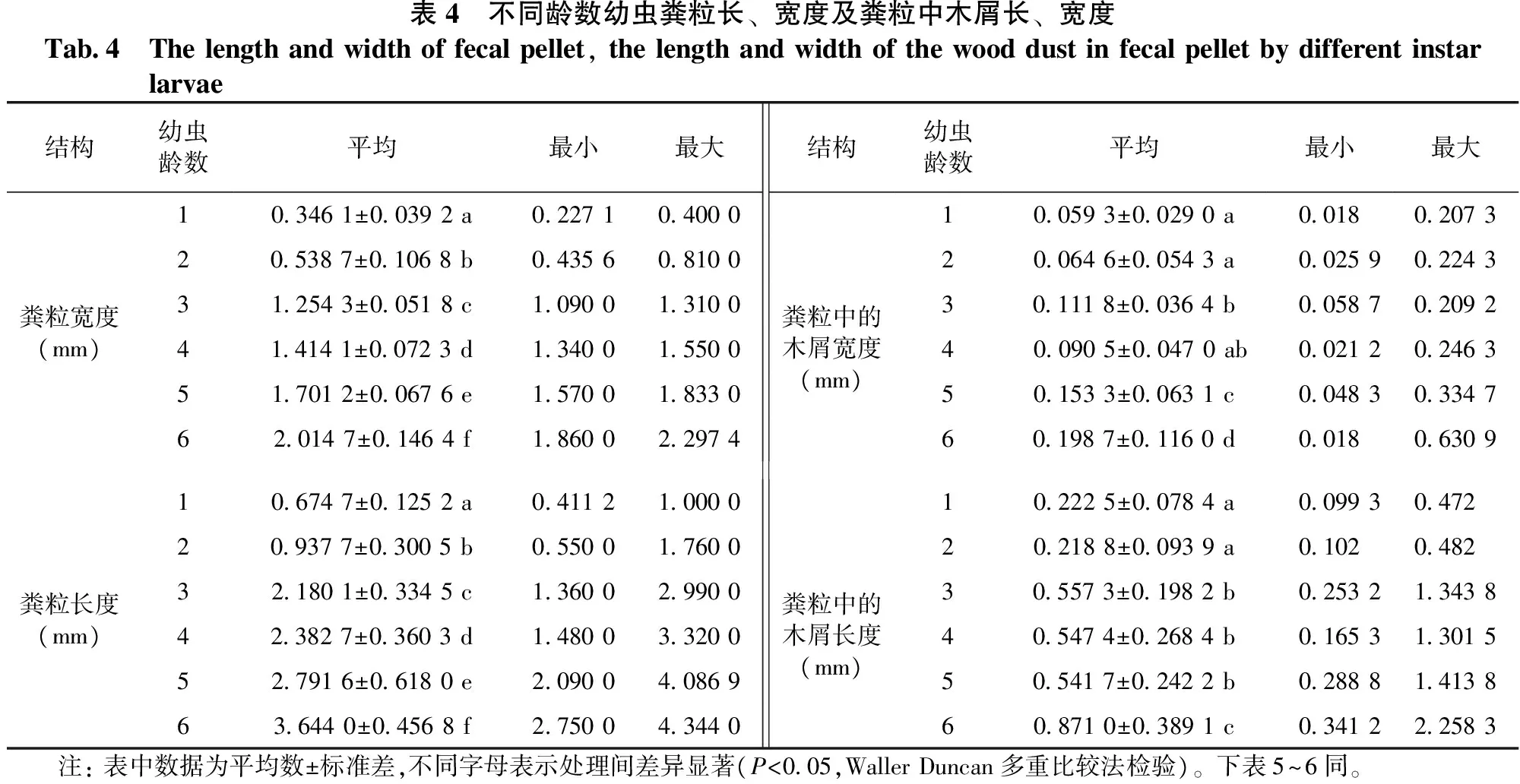
表4 不同龄数幼虫粪粒长、宽度及粪粒中木屑长、宽度Tab4 Thelengthandwidthoffecalpellet,thelengthandwidthofthewooddustinfecalpelletbydifferentinstarlarvae结构幼虫龄数平均最小最大结构幼虫龄数平均最小最大103461±00392a0227104000100593±00290a001802073205387±01068b0435608100200646±00543a0025902243粪粒宽度(mm)312543±00518c1090013100粪粒中的木屑宽度(mm)301118±00364b0058702092414141±00723d13400155004 00905±00470ab0021202463517012±00676e1570018330501533±00631c0048303347620147±01464f1860022974601987±01160d001806309106747±01252a0411210000102225±00784a009930472209377±03005b0550017600202188±00939a01020482粪粒长度(mm)321801±03345c1360029900粪粒中的木屑长度(mm)305573±01982b0253213438423827±03603d1480033200405474±02684b0165313015527916±06180e2090040869505417±02422b0288814138636440±04568f2750043440608710±03891c0341222583 注:表中数据为平均数±标准差,不同字母表示处理间差异显著(P<005,WallerDuncan多重比较法检验)。下表5~6同。
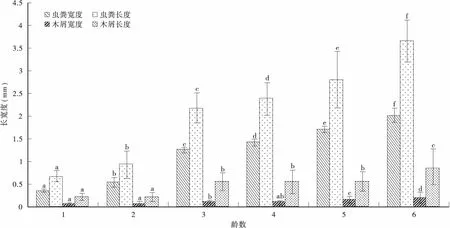
图1 不同龄数幼虫粪粒长、宽度及粪粒中木屑长、宽度Fig.1 The length and width of fecal pellet, the length and width of the wood dust in fecal pellet by different instar larvae
2.3不同龄数幼虫粪粒中组分含量差异
由表5和图2可知: 不同龄数幼虫粪粒中纤维素、木质素含量较均匀,纤维素平均含量为46.72%,木质素平均含量为16.12%。总糖含量、蛋白质含量与吸光度值有较好的线性关系,回归方程为y′=0.0064x,y″=0.0056x-0.0215,相关系数为R2=0.9948、R2=0.9923;粪粒中蛋白质的平均含量为0.1426%,总糖的平均含量为0.6125%。
2.4不同龄数幼虫蛀道的最末排粪孔结构差异
从表6和图3可以看出,最末排粪孔长径、短径在各龄数间的差异均达显著水平。其中6龄幼虫蛀道的最末排粪孔长短径平均值最大,1龄幼虫的最末排粪孔长短径的平均值最小。
2.5最末排粪孔和虫粪结构及虫粪组分含量与幼虫龄数的相关性
表7结果表明:桑天牛的幼虫龄数与蛀道的最末排粪孔长径、最末排粪孔短径、粪粒宽度、粪粒长度、粪粒中木屑宽度、粪粒中木屑长度、粪粒中纤维素含量、粪粒中蛋白质含量均呈线性回归关系。其中幼虫龄数与粪粒宽度的相关系数最大,为0.982;幼虫龄数与蛋白质含量的最小,为0.635;幼虫龄数与粪粒中木质素含量、粪粒中总糖含量无线性相关。表明最末排粪孔长径、最末排粪孔短径、粪粒宽度、粪粒长度、粪粒中木屑宽度、粪粒中木屑长度、粪粒中纤维素含量、粪粒中蛋白质含量可以用于幼虫龄数的判别,其中粪粒宽度是最佳的龄数判别指标;粪粒中的木质素含量、总糖含量不能用于虫龄的判别。
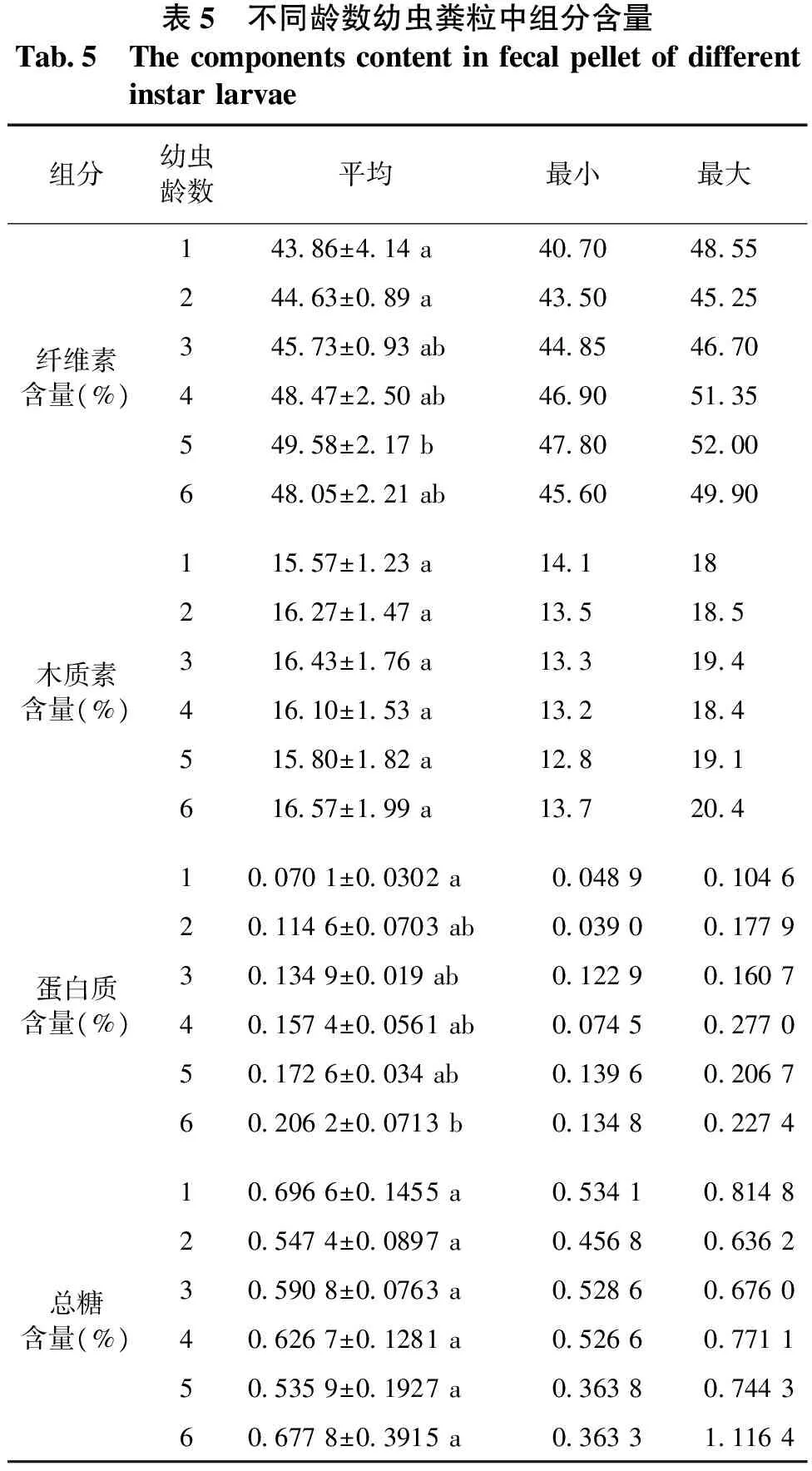
表5 不同龄数幼虫粪粒中组分含量Tab5 Thecomponentscontentinfecalpelletofdifferentinstarlarvae组分幼虫龄数平均最小最大14386±414a4070485524463±089a43504525纤维素含量(%)34573±093ab4485467044847±250ab4690513554958±217b4780520064805±221ab4560499011557±123a1411821627±147a135185木质素含量(%)31643±176a13319441610±153a13218451580±182a12819161657±199a137204100701±00302a0048901046201146±00703ab0039001779蛋白质含量(%)301349±0019ab0122901607401574±00561ab0074502770501726±0034ab0139602067602062±00713b0134802274106966±01455a0534108148205474±00897a0456806362总糖含量(%)305908±00763a0528606760406267±01281a0526607711505359±01927a0363807443606778±03915a0363311164
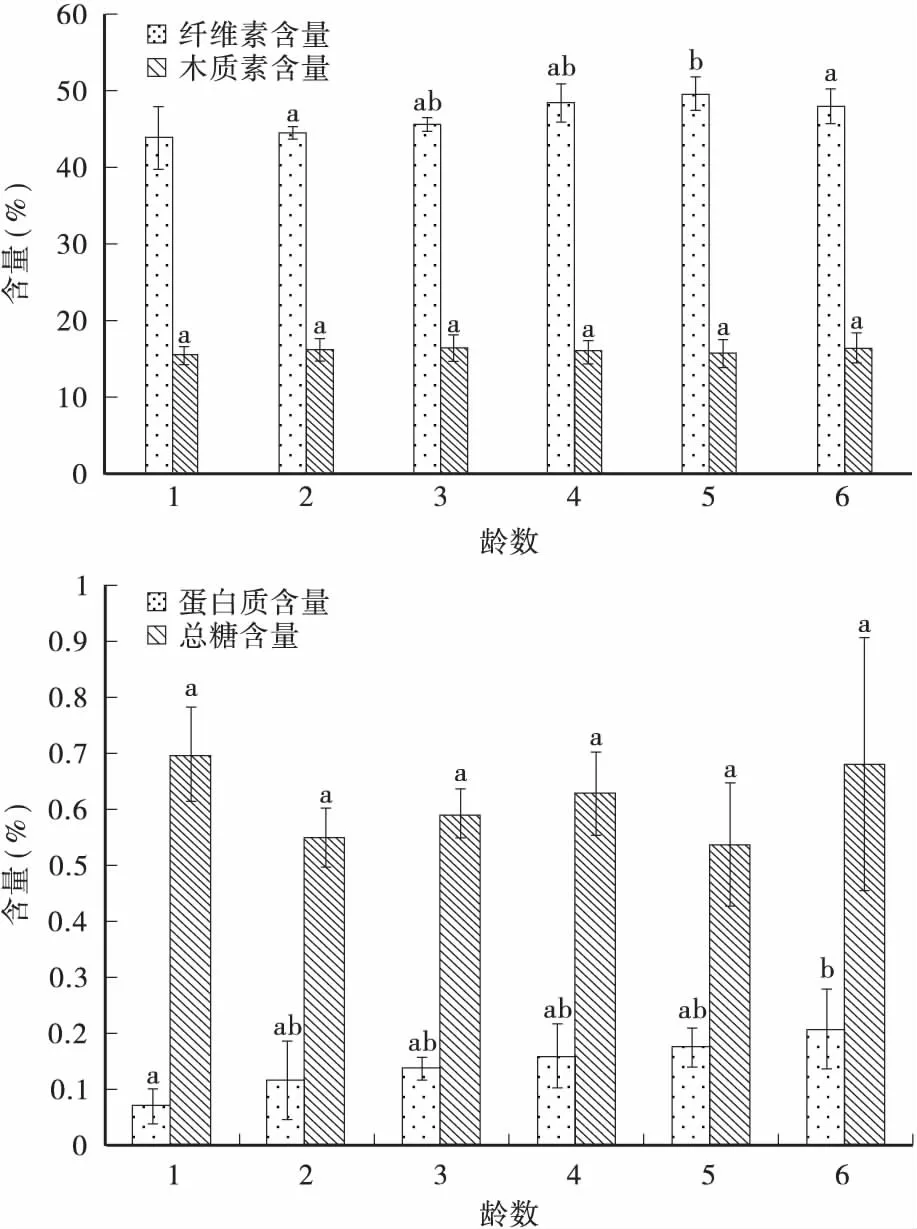
图2 不同龄数幼虫粪粒中的组分含量Fig.2 The components content in fecal pellet of different instar larvae
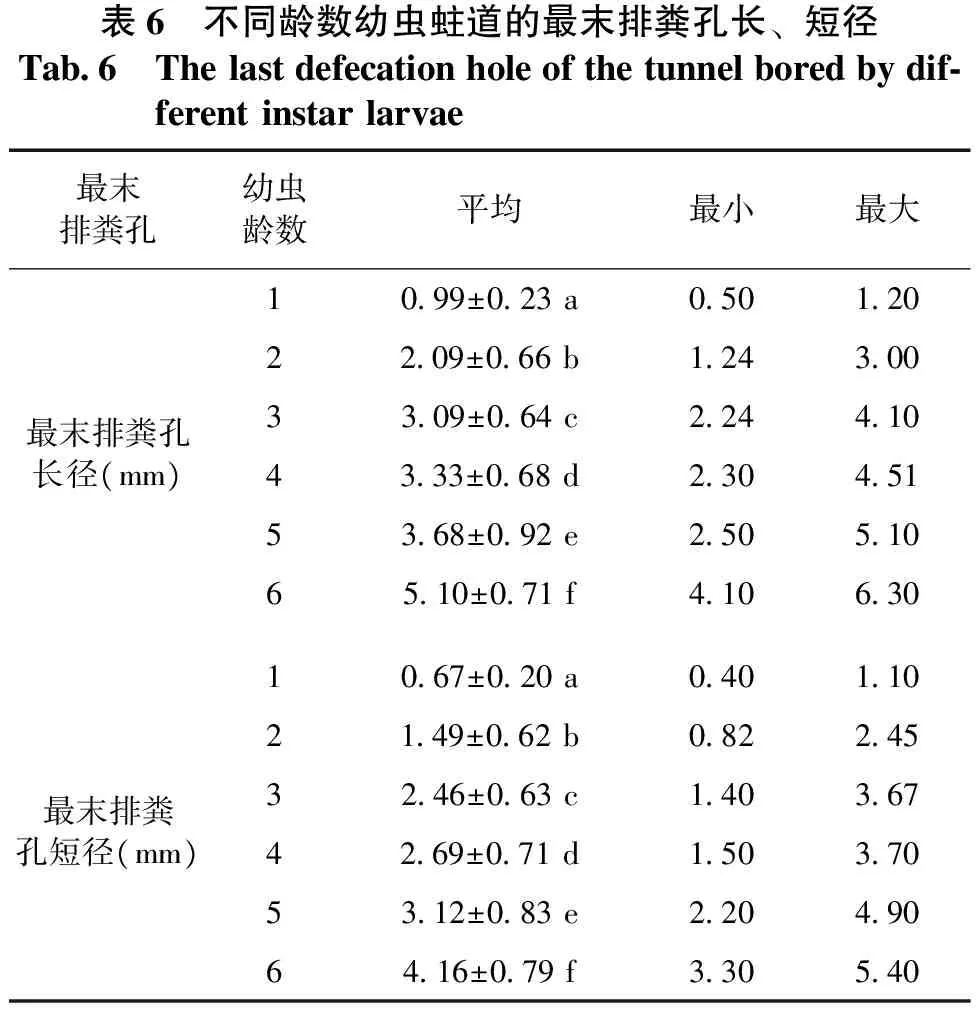
表6 不同龄数幼虫蛀道的最末排粪孔长、短径Tab6 Thelastdefecationholeofthetunnelboredbydif⁃ferentinstarlarvae最末排粪孔幼虫龄数平均最小最大1099±023a0501202209±066b124300最末排粪孔长径(mm)3309±064c2244104333±068d2304515368±092e2505106510±071f4106301067±020a0401102149±062b082245最末排粪孔短径(mm)3246±063c1403674269±071d1503705312±083e2204906416±079f330540
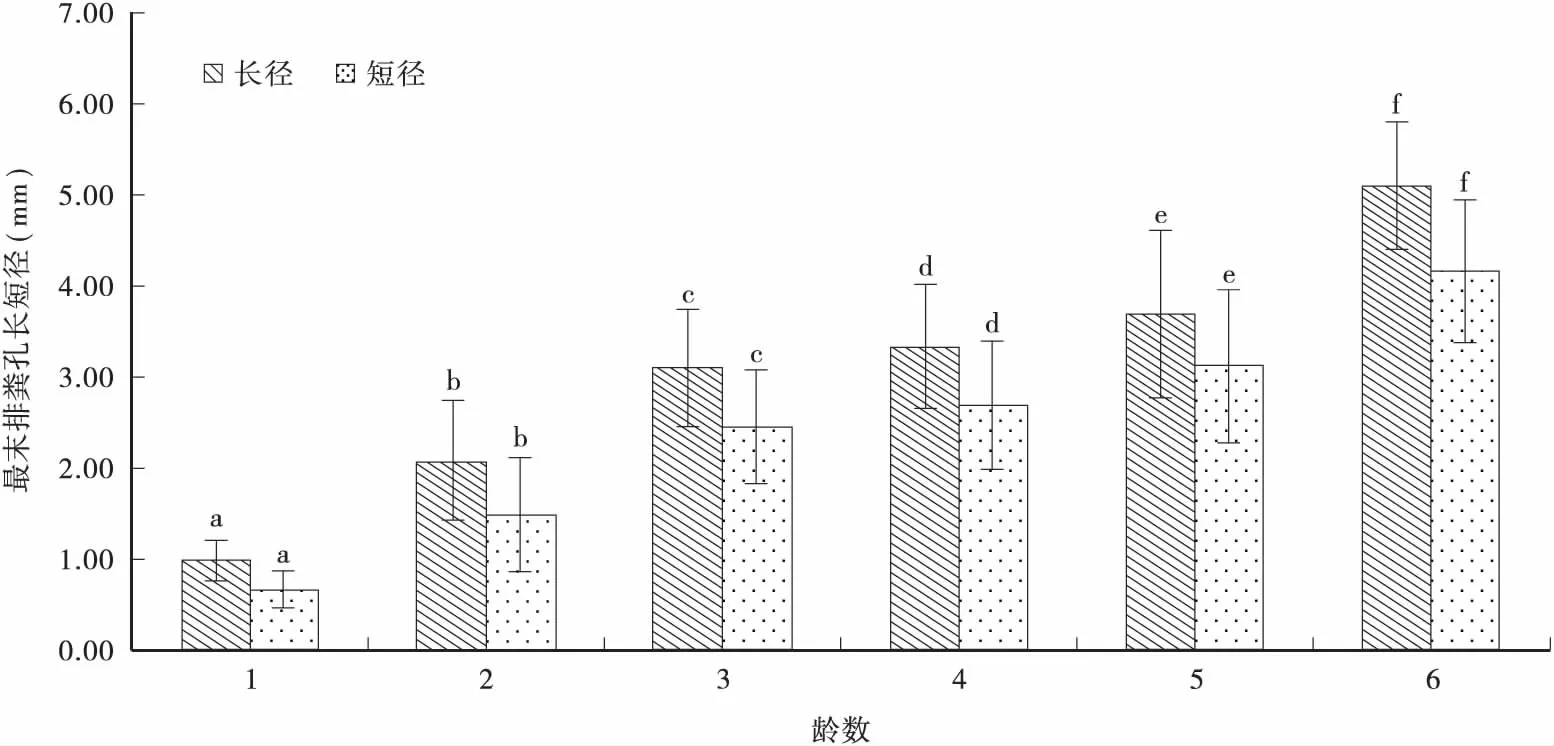
图3 不同龄数幼虫蛀道的最末排粪孔长、短径Fig.3 The last defecation hole of the tunnel bored by different instar larvae
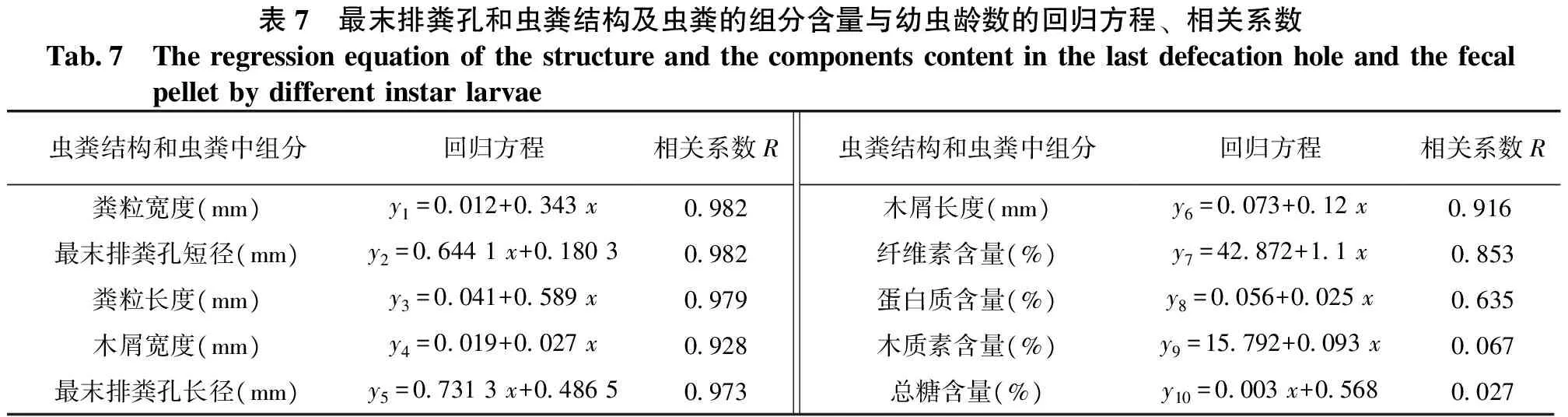
表7 最末排粪孔和虫粪结构及虫粪的组分含量与幼虫龄数的回归方程、相关系数Tab7 Theregressionequationofthestructureandthecomponentscontentinthelastdefecationholeandthefecalpelletbydifferentinstarlarvae虫粪结构和虫粪中组分回归方程相关系数R虫粪结构和虫粪中组分回归方程相关系数R粪粒宽度(mm)y1=0012+0343x0982木屑长度(mm)y6=0073+012x0916最末排粪孔短径(mm)y2=06441x+018030982纤维素含量(%)y7=42872+11x0853粪粒长度(mm)y3=0041+0589x0979蛋白质含量(%)y8=0056+0025x0635木屑宽度(mm)y4=0019+0027x0928木质素含量(%)y9=15792+0093x0067最末排粪孔长径(mm)y5=07313x+048650973总糖含量(%)y10=0003x+05680027
3 结论与讨论
(1) 桑天牛幼虫最末排粪孔长径和短径、粪粒宽度和长度、虫粪中木屑长度和宽度、纤维素含量、蛋白质含量在龄数间存在着线性相关关系,可作为林间虫龄识别指标。其中各龄幼虫的粪粒宽度比较稳定、相互之间没有重叠现象,是林间识别虫龄的最佳指标。桑天牛各龄幼虫虫粪的颜色、形状、质地也存在一定差别,可作为林间虫龄识别的参考。以往划分天牛幼虫龄数均以Dyar规律为基础,通过测定幼虫各项指标后分析得出龄数,其工作量较大[17-21]。而天牛幼虫蛀道的排粪孔比较容易观察,虫粪较易获得,通过对各龄幼虫最末排粪孔大小、虫粪的理化性质分析,找出各因素与龄数之间的关系,最终达到林间快速识别虫龄的目的。
(2) 在林间易于收集到的虫粪一般是被幼虫推出排粪孔外的虫粪,受环境条件(温度、湿度、光照等)的影响比较大。本研究在室内通过解剖蛀道收集虫粪,与林间直接观察的虫粪颜色、大小等特征会有一定差别[9]。因此,在林间收集虫粪时,应尽量选择刚排出的新鲜虫粪,减少环境对虫粪特征的影响。本研究提出的粪粒长宽度、粪粒中纤维素含量等指标可以作为判别桑天牛幼虫虫龄的方法,对其他种类幼虫分龄的研究可提供参考,能否广泛推广应用还有待进一步验证。
(3) 蒋书楠等、殷幼平等认为天牛是否能自身消化纤维素,至今尚无定论,但确证桑天牛消化道中有完整的纤维素酶体系,可以消化食物中的纤维素[13,22]。本研究结果表明不同龄数幼虫的虫粪中纤维素含量有一定变化,这可能与枝干危害部位或是纤维素在幼虫消化道的分解过程有关,具体原因还有待进一步研究。
[1] 杜宝善,刘春梅.黄斑星天牛幼虫龄数测定[J].昆虫知识, 1986(1):22-23.
[2] 柳建定,李百万,王菊英,等.松褐天牛龄级划分方法初探[J].浙江林业科技, 2008,28(4):84-86.
[3] 赖开平,陆温,刘德星,等.眉斑并脊天牛幼虫龄数及各龄特征[J].昆虫知识, 2008,45(1):138-139.
[4] 张海滨,王建伟,朱小峰,等.星天牛幼虫龄期的划分[J].应用昆虫学报, 2011,48(3):688-694.
[5] 黎保清,嵇保中,刘曙雯,等.桑天牛幼虫龄数与划分特征研究[J].林业科技开发, 2012,26(3):38-41.
[6] Eckstein K. Das Bohrmehl des waldgartners,MyelophiluspimperdaL., nebst Bemerkungen uber den ’Frass’ der Borkenafer und anderen Insekten Ara[J]. Physiol.angew.Ent, 1939,6:32-41.
[7] Morris R.F. Frass-drop mersurement in studies of the European spruce sawfly[J]. Uniu. Michigan sch. Forestry and conseru. Bull,1949,12: 58.
[8] Bean J.L. Frass size as an indicator of spruce budworm larval instars[J]. Ann. ent. Soc. Am,1959,52:608.
[9] 薛贤清,严教金,茅洪新,等.利用幼虫粪粒数推算高大树上虫口密度[J].南京林业大学学报, 1986 (4):83-90.
[10] 陈洪章.纤维素生物技术[M].化学工业出版社, 2005.
[11] 丁保福,魏建荣,赵建兴,等.光肩星天牛幼虫虫粪挥发物成分分析[J].环境昆虫学报, 2009,31(3):208-212.
[12] 姜莉,魏建荣,乔鲁芹,等.利用固相微萃取技术分析锈色粒肩天牛幼虫虫粪所含的挥发物成分[J].环境昆虫学报, 2010,32(3):357-362.
[13] 蒋书楠,殷幼平,王中康.几种天牛纤维素酶的来源[J].林业科学, 1996,32(5):441-446.
[14] 李合生.植物生理生化试验原理和技术[M].北京:高等教育出版社,2000.
[15] 王秀奇,秦淑媛,高天慧,等.基础生物化学实验[M].北京:高等教育出版社,2005.
[16] Tuat NH, Binh NT. Khai thác và sǔ dung SPSS dê xǔ ly sliêu nghiên cúu trong lm nghiêp[M]. Hà Ni, Nhà xuát bn Nng nghiêp. 2005.
[17] Morris RF. A use of frass in the identification of forest insect damage[J], Canada Entomol, 1942,74:164-167.
[18] 沙生,庞正轰.油松毛虫食叶量及排粪量的测定[J].北京林业大学学报, 1988,10(4):85-90.
[19] 王缉健.博白松林下虫粪初探[J].广西林业科学, 1996,25(2):100-101.
[20] 张旭,张东流,查光济.虫粪在森林昆虫识别中的应用初探[J].林业科学, 1990,26(5):461-464.
[21] 温艳菊,李继泉,韩永峰,等.取食不同寄主植物桑天牛的虫粪挥发物对桑天牛卵啮小蜂的引诱活性及其组分分析[J].昆虫学报, 2010,53(11):1281-1286.
[22] 殷幼平,程惊秋,蒋书楠.桑粒肩天牛纤维素酶的性质研究[J].林业科学, 1996,32(5):454-458.
(文字编校:唐效蓉)
StudyonlarvalfecesandinstaridentificationofAprionagermari(Hope)
LE Bao-thanh1, YAN Xuewu2*, XU Qian3
(1.Vietnam Forestry University,Hanoi 100803,Vietnam;2.Hunan Forestry Academy,Changsha 410004,China;3.Forestry Bureau of Xiangtan City,Xiangtan 411100,China)
This study obtained poplar branches containingAprionagermarilarvae via indoor breeding and field collecting. In the laboratory, feces of different instar larvae were collected respectively. Feces of different instar larvae cloud separate fecal pellet and residue, and the relation with fecal pellet’ physical and chemical properties and larvae instar was researched. The results showed that the average width of fecal pellet was 1.211 5 mm, the average length of fecal pellet was 2.101 8 mm; The average width of the wood dust in fecal pellet was 0.113 0 mm, the average length of the wood dust in fecal pellet was 0.207 3 mm; The average content of protein was 0.142 6%, the average content of total sugar was 0.612 5%, the average content of cellulose was 46.72%, the average content of lignin was 16.12%. There were liner correlations between the larvae instars and the length of fecal pellet, the width of fecal pellet, the width of the wood dust in fecal pellet, the length of the wood dust in fecal pellet, the content of cellulose and the content of protein respectively. The results showed that aforesaid features used recognition of larva instars in field were feasible. And the width of fecal pellet was the best index for identifying the larva instar rapidly in field, with the least overlap between neighbouring instars.
Aprionagermari(Hope); larval feces; instar; physical and chemical properties
2013-12-02
湖南省林业厅资助项目“张家界松褐天牛生物防治工程”。
LE Bao-thanh(1974-),男,越南河内人,博士,讲师,研究方向为森林保护学。
* 为通讯作者。
S 763.38
A
1003 — 5710(2014)01 — 0007 — 07
10. 3969/j. issn. 1003 — 5710. 2014. 01. 002
猜你喜欢
小哥白尼(野生动物)(2021年1期)2021-07-16
分子影像学杂志(2021年1期)2021-03-19
结直肠肛门外科(2020年5期)2020-11-12
飞天(2020年4期)2020-04-17
世界华人消化杂志(2019年2期)2019-02-21
小学生必读(低年级版)(2018年10期)2019-01-04
故事作文·低年级(2018年10期)2018-10-25
中国继续医学教育(2015年1期)2016-01-06
东北电力大学学报(2015年4期)2015-11-13
作文与考试·小学低年级版(2015年11期)2015-07-17
