
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
2014-11-07 01:46陶冶张元明
草业学报 2014年2期
陶冶,张元明
(1.中国科学院干旱区生物地理与生物资源重点实验室 中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011;2.中国科学院大学,北京 100049)
类短命植物(ephemeroid plant)是指利用早春季或夏初相对充足的水分在短时间里迅速完成生活史,地上部分当年死亡、而地下部分为多年生的一类特殊植物类型[1-2]。类短命植物与短命植物(ephemeral plant)一道成为准噶尔荒漠春季与初夏维持地表稳定、物种多样性及生态系统稳定性的重要贡献者[1,3]。准噶尔荒漠是短命和类短命植物在中亚干旱区分布的最东端,因此,这类植物也是我国荒漠植物区系中重要而独特的组成部分[1-2]。
生物量是植物的基本生物学特征和功能性状之一,是物质和能量积累的基本体现[4-5]。生物量分配指植物生长发育过程中所同化的资源用于根、茎、叶、花、果等各器官的比例,也就是各器官累积的资源在总资源中所占的比例,是植物生殖与生存平衡的结果。生物量分配主要受植物的遗传特性、环境条件和个体大小的影响[6-7],其分配模式在不同生活型植物间差异较大[8]。目前关于荒漠地区短命植物、一年生草本、多年生草本植物等都有较多的研究[8-10],但关于类短命植物这一特殊生活型植物生物量分配的研究仍相当匮乏[11],对其生物量分配的特点和规律仍知之甚少。
异速生长关系,即相关生长关系(allometric relationship),指生物体的某些生物学特征(如个体形态指标、器官生物量等)与个体大小(重量)之间的幂函数关系。这种规律在生物界普遍存在,与尺度无关[12-16]。异速生长关系通过对数化大大减弱了单纯使用生物量学特征的绝对量或比例系数所带来的波动性,能揭示植物器官生物量分配特征间的与尺度无关的内在规律[17-18]。目前,异速生长关系在刻画生物体构件结构与功能特征上得到了大量应用,其中以植物体构件(器官)生物量间的相关生长关系研究最为常见[17]。通过代谢理论和分型网络模型的理论预测均表明,无论有次生组织还是无次生组织的植物,其地上、地下生物量间均为等速生长关系(幂指数为1)[12-16]。
类短命植物是一个生活型的概念,其包含不同科、属的不同物种[1-2]。那么同为短命植物类群,不同物种是否具有一致或相似的生物量分配比例?它们的器官生物量间是否具有一致的相关生长关系?为解决上述科学问题,我们选择了准噶尔荒漠常见的6种类短命植物:百合科(Liliaceae)的小山蒜(Alliumpallasii)、棱叶韭(Alliumcaeruleum)、粗柄独尾草(Eremurusinderiensis)、黑鳞顶冰花(Gageanigra)、伊犁郁金香(Tulipailiensis),以及菊科(Asteraceae)的细叶鸦葱(Scorzonerapusilla)作为研究对象。这6个物种既有不同科的植物,又有同科不同属的和同属不同种的植物。通过对其生物量分配和异速生长关系的对比研究,期望找出类短命植物生物量分配的一些特点及器官生物量分配的协同变异关系,为深入了解该类生活型植物的生存策略与生态功能提供科学依据,并为不同生活型植物构件功能特征的对比研究提供参考。
1 材料与方法
1.1 研究区概况
准噶尔荒漠位于新疆北部,其中心为我国最大的固定和半固定沙漠——古尔班通古特沙漠(44°11′~46°50′ N,84°31′~91°20′ E,4.88×104km2)。该沙漠年均蒸发量>2000 mm,而年均降水量普遍不超过150 mm,沙漠腹地仅有70~100 mm,且主要集中于冬春季;年均温6~10℃,极端高温为40℃以上,≥10℃的年积温可达3000~3500℃,空气相对湿度平均50%~60%,5-8月通常<45%[19-20]。准噶尔荒漠是我国唯一一个受北冰洋气流影响的荒漠区,其物种组成和丰富度与我国其他荒漠地区差异明显[1]。梭梭(Haloxylonammodendron)、白梭梭(Haloxylonpersicum)、心叶驼绒藜(Ceratocarpusewersmanniana)、假木贼(Anabasis)等灌木是准噶尔荒漠植物群落的主要建群种。该地区具有丰富的短命和类短命植物,是早春荒漠生态系统的重要组成部分;同时,准噶尔荒漠也是该类群植物在中亚干旱区分布的最东端[2]。
1.2 植物样品采集与处理
所有植物样品均于2010年5月底类短命植物开花末期采集,6种植物均处于相同物候期。粗柄独尾草、黑鳞顶冰花、细叶鸦葱和小山蒜均采自古尔班通古特沙漠南部地区,伊犁郁金香采自乌鲁木齐雅玛里克山(海拔960 m),棱叶韭采自石河子将军山(海拔580 m)。3个采集地点均属于准噶尔荒漠,其中古尔班通古特沙漠位于准噶尔盆地中央,土壤类型为风沙土;乌鲁木齐雅玛里克山和石河子将军山均属于准噶尔盆地南缘低山丘陵荒漠地貌,土壤类型为棕钙土。
野外调查时,设置2个粗柄独尾草样地,大小为15 m×15 m;4个小山蒜样地,大小为5 m×5 m;其余物种均设置2个样地,大小为10 m×10 m。利用全株挖掘法获取样地内全部供试植物生物量。在获取地下生物量前,将每株植物地上部分齐地面剪除待用。植物根系挖掘的广度和深度因种而异,以植株为圆心,粗柄独尾草挖掘半径为0.6~0.8 m,深度约为0.7 m;黑鳞顶冰花为0.1和0.2 m;细叶鸦葱为0.3和0.6 m;小山蒜为0.1和0.2 m;伊犁郁金香为0.2和0.4 m。粗壮主根直接收获,断裂的细根利用1~2 mm土筛筛选。将每株植物的地上和地下两部分分装在做好标记的信封内,带回实验室后置于75℃烘箱内烘48 h,以保证样品完全烘干。将每株植物的地下根系、地上叶片及繁殖器官单独称重、记录。所用天平精度为0.0001 g。
1.3 生物量及其分配比例计算
百合科植物属于单子叶植物,没有明显的茎,因此,将5种百合科类短命植物个体统一分为根、叶和繁殖器官(包括花和花序轴) 3个部分。分别计算各组分(地上生物量AGB、地下生物量BGB、总生物量TB和叶片生物量LB)的生物量大小,并计算根冠比(root to shoot mass ratio, R/S)、叶生物量比(leaf to total plant mass ratio, L/T)和繁殖分配(reproduction allocation ratio, RAR)。各生物量指标均采用平均值±标准误(mean±SE)的形式给出。
1.4 数据统计分析
对6个物种的AGB、LB、繁殖器官生物量(RB)、BGB、TB、R/S、L/T及RAR进行One-way ANOVA分析。利用Levene’s test检验方差齐性与否,方差齐性时使用Tukey’s HSD test进行多重比较,方差不齐时则使用T2 Tamhane’s test进行多重比较[21]。R/S、RAR随个体大小(地上生物量)的变化趋势通过幂函数来判断。常规数据分析和作图在Excel 2003中完成,One-way ANOVA分析在SPSS 19.0中实现。
1.5 异速生长分析
异速生长关系是生物量分配和器官功能关系研究的重要手段和指标[17-18,22-23]。异速生长关系可以表示为Y=β·Xα,其中,Y是某种生物学特征或者功能,β是标准化常数,X指个体大小,α是异速生长指数。α=1为等速关系,即因变量和自变量呈均匀或等比例生长;α≠1为异速关系,即因变量和自变量间为不均匀或不等比例生长。确定异速生长参数时,须将幂函数转换为logY=logβ+α·logX的形式,利用降主轴回归(reduced major axis regression, RMA, 即Model Type II)方法计算回归模型的指数、95%置信区间(95% CI)及决定系数(R2)[17,22]。普通最小二乘回归(ordinary least squares regression, OLS, 即Model Type I)多用于预测2个变量之间的变化趋势,当需要了解2个变量之间的功能关系时,RMA回归是最合适的方法,而且该方法也是异速生长分析的标准方法[12-17]。对数化之后的α即为幂函数对数化之后线性回归的斜率,logβ为线性回归的截距。
不同物种间(不同群体间)的异速生长关系主要有图1所示的4种情形[24-25]。一般地,除计算和比较异速生长指数α(即同斜率检验,Shift A检验)和截距(logβ)外,还需要对α进行等速生长检验(即与1.0进行比较)。对若干物种间的α进行多重比较时,若α相同(α不同即出现了Shift A),则需要进一步比较它们的截距是否相同(即截距漂移检验,Shift B检验),同时对α相同的这些物种间进行共轴漂移检验(Shift C检验),即判断它们是否具有同一的线性拟合轴。此时,如果截距不同但共轴,则属于Shift B情形;如果截距相同但发生了共轴漂移,则就属于Shift C的情形;如果截距不同轴也不同,则属于Shift D的情形;如果截距相同也同轴,那就表明这些物种的这对指标间具有完全相同的相关生长关系[24-25]。
上述异速生长指数(α)和截距(logβ)的计算、等速生长检验、截距漂移检验、共轴漂移检验及α的方差分析均使用SMARTR软件完成[26]。
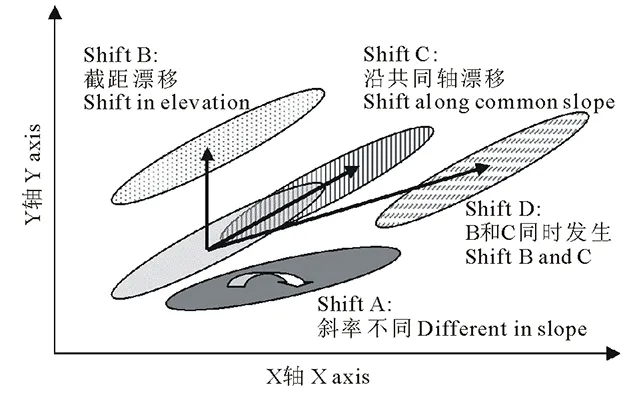
图1 不同群体间异速生长关系几种可能的变化情形[24-25]Fig.1 Schematic illustrating possible differences of allometric relationships among different plant groups[24-25]
2 结果与分析
2.1 生物量及其分配特点
准噶尔荒漠6种类短命植物单株生物量差异明显(表1)。AGB、BGB、LB、RB和TB均以粗柄独尾草为最大,黑鳞顶冰花最小。以TB为例,粗柄独尾草(21.144 g/株)是黑鳞顶冰花(0.094 g/株)的近225倍。TB大于1 g/株的有3种(分别是粗柄独尾草、细叶鸦葱和棱叶韭),另外3种在0.094~0.702 g/株之间。小山蒜和伊犁郁金香的TB最为接近(分别为0.628和0.702 g/株),细叶鸦葱和棱叶韭之间也无显著差异(分别为1.290和1.855 g/株),四者的生物量均介于粗柄独尾草和黑鳞顶冰花之间。
细叶鸦葱具有最大的R/S (3.066±0.223),其次是粗柄独尾草(1.441±0.077),黑鳞顶冰花(0.842±0.035)和小山蒜(0.873±0.028)的R/S基本相等,棱叶韭(0.355±0.019)的R/S最小(图2)。6种类短命植物的L/T以黑鳞顶冰花为最大(0.289±0.006),其次是粗柄独尾草(0.166±0.008)、细叶鸦葱(0.151±0.007)和伊犁郁金香(0.171±0.006),最小的是同为葱属的小山蒜和棱叶韭,仅为(0.105±0.004)和(0.092±0.004)。繁殖分配比例(RAR)最大的是棱叶韭(0.649±0.011),其次为伊犁郁金香(0.496±0.012)和小山蒜(0.433±0.008),最小的是细叶鸦葱(0.111±0.013),而粗柄独尾草(0.251±0.011)和黑鳞顶冰花(0.261±0.007)的RAR几近相等。从繁殖器官生物量与叶片生物量之比(RB/LB)也可以看出,棱叶韭(7.396±0.398)、小山蒜(4.394±0.242)和伊犁郁金香(3.046±0.161)仍较大,细叶鸦葱(0.766±0.105)仍为最小。
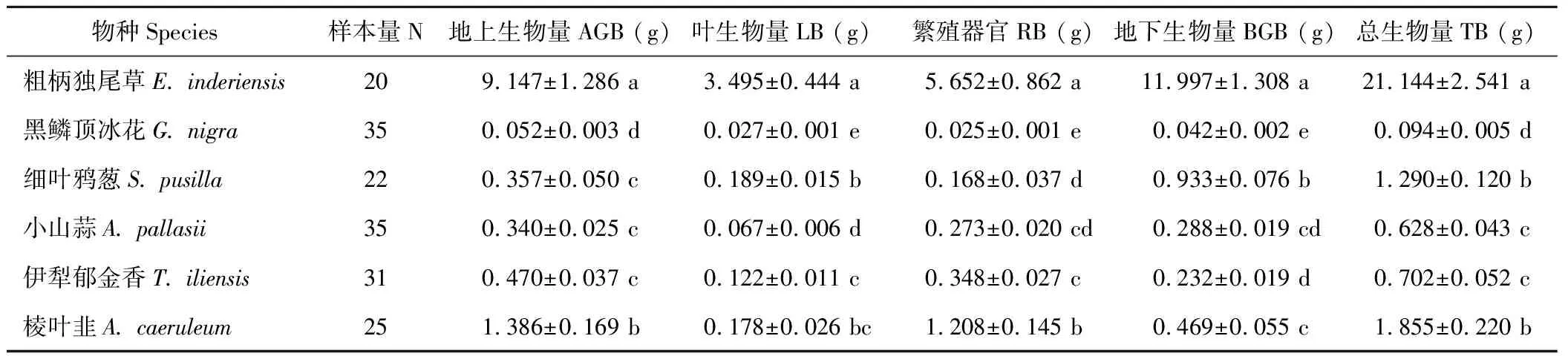
表1 6种类短命植物地上生物量、叶片生物量、繁殖器官生物量、地下生物量及全株生物量Table 1 The aboveground biomass (AGB), leaf biomass (LB), reproduction organ biomass (RB), belowground biomass (BGB) and total plant biomass (TB) of six ephemeroid species
同列不同字母表示差异显著(P<0.05)。Different letters in the same row indicate significant difference (P<0.05).
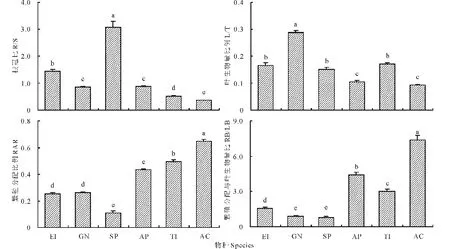
图2 6种类短命植物根冠比、叶生物量比、繁殖分配比例及繁殖生物量与叶片生物量之比Fig.2 The root to shoot mass ratio (R/S), leaf to total plant mass ratio (L/T), reproduction allocation ratio (RAR) and reproduction organ biomass to leaf biomass ratio (RB/LB) of six ephemeroid species 物种EI、GN、SP、AP、TI、AC分别代表粗柄独尾草、黑鳞顶冰花、细叶鸦葱、小山蒜、伊犁郁金香和棱叶韭,不同字母表示差异显著(P<0.05)。The species EI, GN, SP, AP, TI and AC mean E. inderiensis, G. nigra, S. pusilla, A. pallasii, T. iliensis and A. caeruleum; different letters indicate significant difference (P<0.05). 下同 The same below.
总体上,6种类短命植物用于有性繁殖和光合作用的生物量分配比例是不一致的,其中棱叶韭、小山蒜和伊犁郁金香的繁殖分配比例最高(即叶片生物量分配比例低),细叶鸦葱最低。因此,尽管同为类短命植物,不同科、不同属及同属不同种之间的生物量大小及器官分配比例仍差异明显,没有一致的规律可循。
2.2 生物量分配与个体大小的关系
随着植株个体大小的增加,6个物种R/S均呈现下降趋势(图3),除伊犁郁金香外,其他5个物种的R/S下降趋势均达到了显著(P<0.05)或极显著水平(P<0.01),其中以细叶鸦葱最为明显(幂指数为-0.496,R2=0.6498)。这种下降趋势在个体较小时最为强烈,当个体较大时,R/S的下降速度趋缓。因此,即使是同一种植物,由于个体大小的不同,同化产物分配到地上和地下的绝对量也是不同的;但同一种内,体型较大的植物具有相对稳定的地上和地下生物量分配比例。
RAR也随着植株个体大小的增大发生变化(图4),但变化趋势基本与R/S相反。粗柄独尾草、黑鳞顶冰花、细叶鸦葱和小山蒜4种植物的RAR随植株个体的增大均呈显著(P<0.05)或极显著(P<0.01)增加趋势,其中以细叶鸦葱的增加趋势最为明显(幂指数为0.8286,R2=0.7834),表明四者的繁殖分配与个体大小之间存在显著的异速生长关系(幂指数<1)。伊犁郁金香和棱叶韭的RAR随个体的增大没有显著变化(P>0.05),说明二者用于有性繁殖的生物量分配比例是基本恒定的(幂指数趋于0),与个体大小无关。
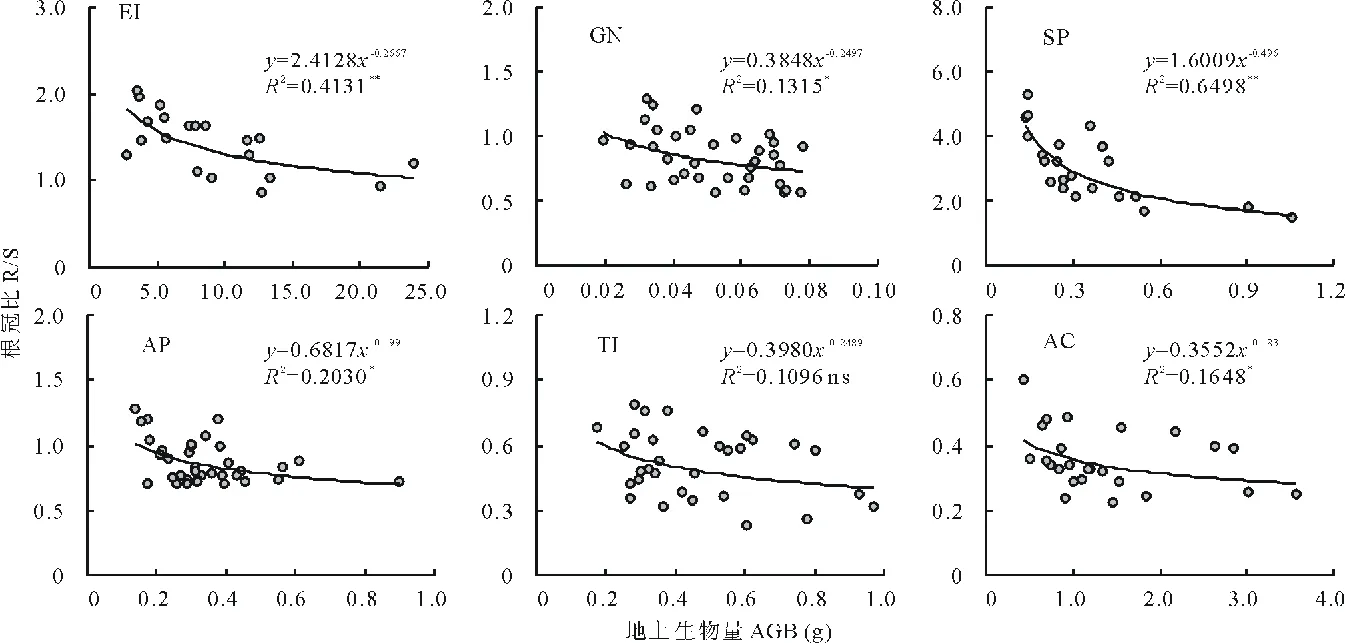
图3 6种类短命植物根冠比随个体大小(地上生物量)的变化趋势Fig.3 The change trends of root to shoot mass ratio (R/S) along with plant size (aboveground biomass, AGB) of six ephemeroid species*: P<0.05, **: P<0.01, ns: P>0.05. 下同 The same below.
2.3 构件生物量间的相关生长关系
6个物种AGB-BGB间、AGB-LB间以及LB-BGB间均具有极显著(P<0.01)的相关生长关系(图5,表2)。等速生长检验表明,粗柄独尾草和细叶鸦葱的AGB-BGB间、细叶鸦葱的AGB-LB间,以及小山蒜和棱叶韭的LB-BGB间的异速生长指数(α)显著<1.0,也即属于因变量分配速率小于自变量的异速生长关系。尽管如此,6种植物AGB-BGB间及LB-BGB间的α差异并不显著,且各具有一个共同的α:AGB-BGB间共同的α为0.8764,LB-BGB间共同的α为0.8585,二者均显著小于1.0,也属于异速生长关系(表2)。6种植物AGB-LB之间的α差异明显,且没有共同的α;其α最大的是小山蒜、伊犁郁金香和棱叶韭,其次是粗柄独尾草和黑鳞顶冰花,最小的是细叶鸦葱(仅为0.6700),而且也仅有细叶鸦葱是异速生长关系。
总体来看,不同科、不同属以及同属不同种之间,器官生物量AGB-BGB间以及LB-BGB间都具有相同的α,与物种种类无关。而对AGB-LB而言,菊科的细叶鸦葱与百合科的5个物种之间的α均差异显著,表明AGB-LB间的异速生长关系在科间是不同的;同属的小山蒜和棱叶韭的α无显著差异,即AGB-LB间的异速生长关系(仅指α)在属内是相同的;而不同属之间AGB-LB间的相关生长关系既有相同的类群(组1内相同,组2内相同;组1:小山蒜、伊犁郁金香和棱叶韭,组2:粗柄独尾草和黑鳞顶冰花),也有不同的类群(组1和组2之间不同)。因此,6种植物器官生物量间的相关生长关系(仅指α)也不尽一致,但大部分表现出等速生长关系和与系统分类无关的特点。
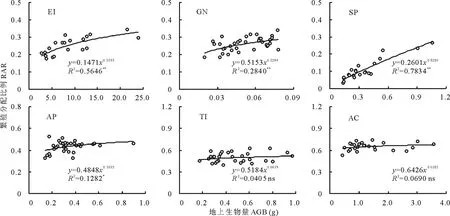
图4 6种类短命植物繁殖分配比例随个体大小(地上生物量)的变化趋势Fig.4 The change trends of reproduction allocation ratio (RAR) along with plant size (aboveground biomass, AGB) of six ephemeroid species
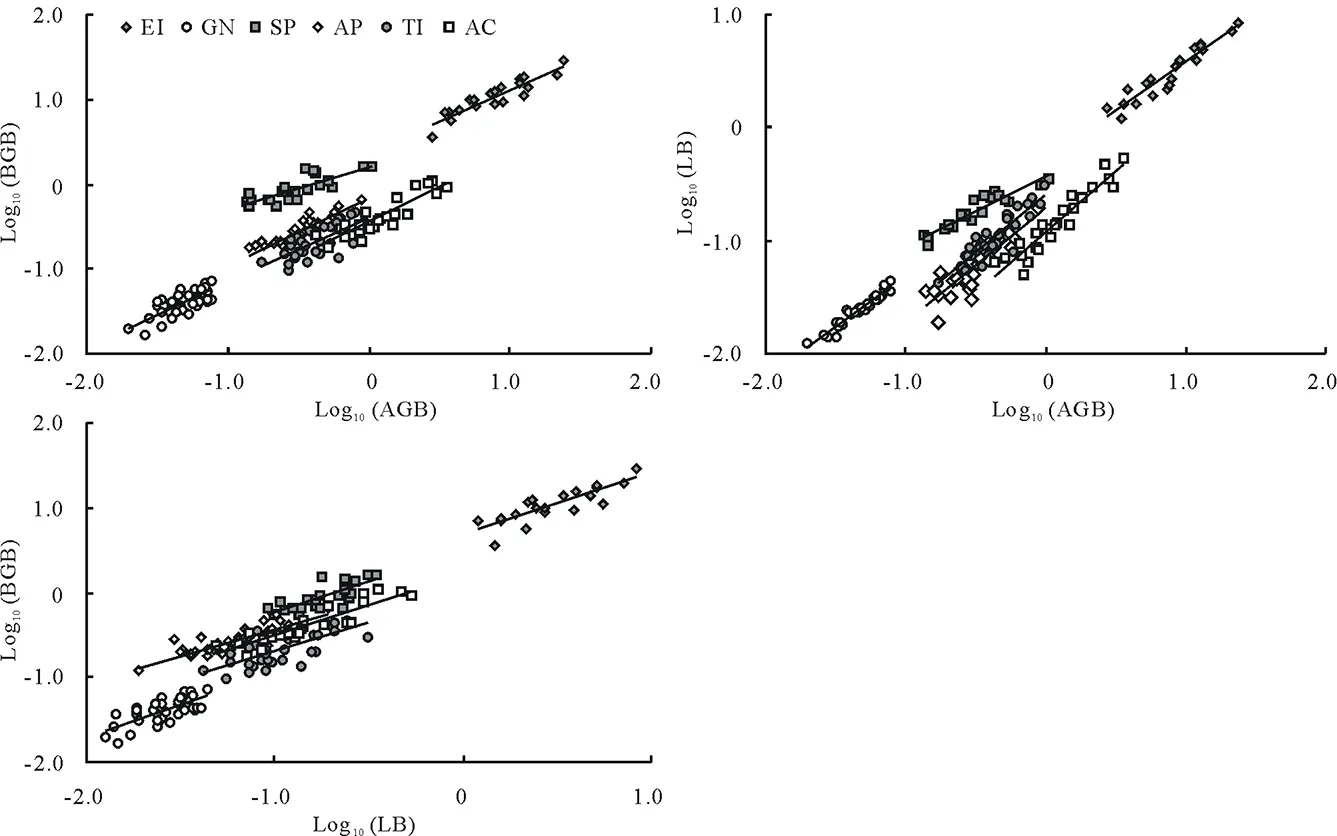
图5 双对数(以10为底)坐标下6种类短命植物地上(AGB)与地下(BGB)、地上与叶片(LB)及叶片与地下生物量间的RMA线性拟合关系Fig.5 Log10-Log10 linear regression (RMA) plots of aboveground biomass (AGB) and belowground biomass (BGB), AGB and leaf biomass (LB), and LB and BGB of six ephemeroid species
由于6个物种AGB-BGB间,以及LB-BGB间各自具有共同α(即未发生Shift A) (表2),故而进一步检测它们是否具有共同的截距(Shift B检验)和线性拟合轴(Shift C检验)。结果表明(表3),AGB-BGB间粗柄独尾草和细叶鸦葱、黑鳞顶冰花和伊犁郁金香的截距没有差异,但发生了共轴漂移(Shift C);LB-BGB间的粗柄独尾草和细叶鸦葱、粗柄独尾草和小山蒜则发生了截距漂移(Shift B);其他所有种对间均同时发生了截距漂移和共轴漂移(Shift D)。
相关生长指数(α)一列的不同字母表示6个物种间α差异显著(P<0.05)。 Different letters indicate significant difference (P<0.05) of six allometric scaling exponents (α).
表3 6种类短命植物3对生物量指标共45个种对异速生长关系类型Table 3 Allometric relationships of 45 species pairs related with three biomass pairs of six ephemeroid species
物种代号1~6分别是EI、GN、SP、AP、TI、AC,同图2。The species number 1-6 are EI, GN, SP, AP, TI and AC, the same with Fig.2.
6个物种ABG-LB间没有共同α,有11个种对的α差异显著(发生Shift A) (表3),但有2组 (组1:小山蒜、伊犁郁金香和棱叶韭,组2:粗柄独尾草和黑鳞顶冰花)共4对植物的α间没有显著差异(表2)。对这4个种对进行进一步的异速生长检验表明,粗柄独尾草和黑鳞顶冰花具有相同的截距但发生了共轴漂移(Shift C),另外3对既发生了截距漂移又出现了拟合轴的漂移(Shift D)。因此,LB-ABG间并没有Shift B的情形出现。
3 讨论
3.1 不同生活型植物生物量分配特征
长期生活在相同环境下的不同物种,往往会形成相似的生物学特征,这就是趋同适应[27]。类短命植物便是由不同物种组成的具有相似生物学特性的一类功能群(functional group),它表现为地上短生、地下多年生的特点,是草本植物对干旱荒漠气候和不均匀的季节性降水的适应。植物生物量分配是指植物将有限资源分配到不同器官及结构功能上,从而实现对外界环境的更好适应[28]。对干旱区的草本植物而言,高的根生物量分配比例也不是荒漠化演替中植被的普遍特征,其比值的高低变化和生活型或温度的关系也许更强于和干旱程度的关系[29]。
研究表明,一次结实的草本植物繁殖分配一般占其净同化能量的20%~40%,多次结实植物在20%以下[30]。对准噶尔荒漠几种胡卢巴属短命植物生物量分配的研究表明,其繁殖分配一般在50%以上,其中网脉胡卢巴(Trigonellacancellata)可达62.3%[10]。大部分短命植物的根冠比在0.1以下,地下生物量分配比例很小[9]。对科尔沁沙地70种草本植物生物量分配研究表明,无论一年生、二年生还是多年生草本,绝大部分的物种繁殖分配比例<20%、叶片生物量比例<30%,而大部分多年生草本植物表现出更大的根冠比[8]。可见,不同生活型草本植物间的生物量分配有较大差异。
艾沙江等[11]对天山北麓特有类短命植物新疆郁金香(Tulipasinkiangensis)生物量分配的研究表明,在果实成熟期新疆郁金香地下鳞茎生物量占全株的50%左右(也即根冠比约为1),繁殖分配比例可达35%以上,而叶片生物量比例不到15%。可见,新疆郁金香具有较高的有性繁殖分配比例和根系生物量分配比例,前者为土壤提供大量的种子库来源,保证了该物种有性繁殖的继续,而后者则为植株存储足够的有机物质以保证来年植株的再次萌发。
本研究表明,6种类短命植物根冠比在0.355~3.066之间,叶片生物量比例在9.2%~28.9%之间,繁殖分配比例在11.1%~64.9%之间。可见,类短命植物根的生物量在全株生物量中占有很大的比例。不同物种间的有性繁殖和光合作用的生物量分配也有明显差异。菊科的细叶鸦葱与百合科的粗柄独尾草和伊犁郁金香具有相似的叶片生物量比例,且同为葱属的小山蒜和棱叶韭的叶片生物量比例也相近(但最小)。棱叶韭、伊犁郁金香和小山蒜的繁殖器官生物量比例较大,细叶鸦葱最小。因此,这种生物量分配比例的差异表明不同科、不同属及同属不同种之间的生物量大小及器官分配比例没有一致的规律可循。尽管如此,与荒漠一年生草本[8]和短命植物[7]相比,类短命植物地下生物量比例较大,体现了类短命植物地上短生、地下多年生的特点。
同一物种不同生境间的生物量分配一般会发生明显改变,主要归因于植物对环境胁迫的适应[25,31],体现了植物的可塑性(plasticity)。而不同物种间生物量分配的异同可能主要体现了系统分类地位或遗传发育的影响,如本研究发现同为葱属的小山蒜和棱叶韭具有相同的叶片生物量比例和相近的繁殖器官生物量比例,而属间及科间不同物种生物量分配的相似性很低。因而,属一级分类单元内的物种间可能具有较相似的生物量分配特征,而属间及科间则会有较大差异。
3.2 植物生物量分配与个体大小的关系
个体大小显著影响植物生物量向营养器官和繁殖器官的分配比例,二者之间存在“此消彼长”的权衡关系[11]。换句话说,生物量分配存在显著的个体发育漂变(ontogenetic drift)[25]。本研究中,6种短命植物中有5种植物的根冠比随个体的增大而显著减小,表明个体较大的植株其根系生物量分配比例下降而地上部分生物量分配比例增加。增加的地上生物量分配既可能用于营养器官,也有可能用于繁殖器官,这主要取决于外界环境和资源状况[32]。当外界环境较优越时,植物会更多地将同化产物分配给营养器官,而偏轻于繁殖器官;反之,当外界环境较为恶劣时,植物会增大繁殖投入,以确保产出足够的种子。
综合以往研究,植物花期繁殖体重量确实随植株个体的增大而显著增加,但是繁殖分配与个体大小间则没有表现出固定的模式。5种毛茛科(Ranunculaceae)植物[33]、3种风毛菊属(Saussurea)植物[34]、黄帚橐吾(Ligulariavirgaurea)[35]等均表现为个体越大繁殖分配越低,即个体大小与繁殖分配存在异速关系,说明这些物种的繁殖分配受个体大小制约。小半灌木油蒿(Artemisiaordosica)的繁殖分配则呈现出在一定范围内随个体增大而上升,然后又随个体继续增大而下降的趋势[36]。青藏高原东缘高寒草甸5种一年生龙胆属(Gentiana)植物繁殖分配与个体大小间无固定的相关模式,体现了不同物种对相似环境压力的适应对策不同[37]。垂穗披碱草(Elymusnutans)的繁殖分配比例与个体大小间没有明显的相关性,其繁殖分配比例表现为一个相对固定的值[38]。类似地,4 种胡卢巴属(Trigonella)短命植物的繁殖器官生物量也不随个体大小而发生变化,表明其生殖生物量是不具大小依赖性的,营养生长与生殖生长的增加是成比率的[10]。
本文对6种类短命植物的研究表明,有4种植物(粗柄独尾草、黑鳞顶冰花、细叶鸦葱和小山蒜)的繁殖分配随个体的增加而显著增加,但属于明显的异速生长关系(幂指数<1),也即繁殖分配与个体大小的生长是不等比例的;另外2个物种(伊犁郁金香和棱叶韭)的繁殖分配没有显著变化。这表明不同种类的类短命植物器官生物量分配与个体大小间无一致的变化规律,但多表现为显著的异速增加的趋势。由此可见,本研究的结果与毛茛科植物[33]、风毛菊属植物[34]及黄帚橐吾[35]等生境相对优越的植物的研究结果相反,也不同于同地区的短命植物的结果[10],这是否是类短命植物的一个共同特点?
3.3 类短命植物构件生物量间的相关生长关系
植物体生物学特征间的相关生长关系已通过分形分配网络模型、代谢理论等多种方法得到有力证明,并得到大量实测数据的验证[12-18,23]。相关生长关系与尺度无关,其通过对数化除去了个体大小(尺度)对生物量分配规律的影响,揭示了不同物种间生物量分配的基本速率问题。相关生长关系理论认为在全球尺度范围内,自然条件下种内和种间个体水平生物量具有相同的分配速率——由一个世界范围的数据库(包括生长于不同生境的草本与树木,包括双子叶植物、单子叶植物和裸子植物)得到[12-16]。相关指数始终为1.0体现了恒定的资源分配速率,而变化的相关常数logβ则反映了不同物种器官生物量绝对量的差异,在自然条件下这是由物种自身属性所决定的。
因此,仅从异速生长指数上看,6种植物器官生物量间大部分表现为等速生长关系,这符合理论预测[12-16];而且AGB-BGB间及LB-BGB还各自具有共同的异速生长指数,这是物种间趋同适应的表现;但6个物种间没有出现完全一致的相关生长关系,这可能是由系统分类地位决定的遗传特性的差异导致的。
致谢:中国科学院新疆生态与地理研究所博士研究生周晓兵、程军回、吴林参与了野外取样,新疆农业大学硕士研究生张乐、王韵、齐会楠帮助测定部分植物的生物量,邱东帮助测定、录入和校对数据,在此一并感谢。
猜你喜欢
今日农业(2020年19期)2020-12-14
东坡赤壁诗词(2020年3期)2020-07-04
热带农业科学(2018年2期)2018-06-21
学生导报·东方少年(2018年4期)2018-05-14
速读·中旬(2017年11期)2017-11-11
理论导刊(2017年8期)2017-08-30
广西林业科学(2016年3期)2016-03-16
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2012年5期)2012-10-26
